
Abstract
The current study explored the potential links between breast cancer and human interferon α-2b (hIFNα-2b) gene mutations. The hIFNα-2b gene was amplified from breast cancer tumor tissue samples (N = 60) by polymerase chain reaction (PCR) and the products were subjected to gene sequencing. A total of 38 (63.3%) samples showed positive PCR amplification results. Several of these also exhibited frequent alterations (mutations) after 400 bp and, in particular, adenine was replaced by other bases. A total of 19 selected mutated amino acids were analyzed for local/general fold pattern changes. Human IFNα-2b receptor (IFNAR): ligand (hIFNα-2b protein) interactions through a Z-DOCK (3.0.2) server were also evaluated to assess the binding patterns of each ligand to receptor to induce Janus-Kinase-signal transducer and activator of transcription antiproliferative signal transduction pathway inside the cancer cells. Certain local structural and conformational changes were predicted to be induced by mutations in the ligand. The variant models of the hIFNα-2b displayed structural and conformational changes that signified that changes to hIFNα-2b may be a risk factor in addition to other known factors associated with onset/progression of female breast carcinoma. It was hoped that others might build upon the research in this study evaluating protein structural models with mutations and their consequent interactions with receptors in the development of potent immune therapeutic drugs for breast cancer that are based on recombinant hIFNα-2b.
Introduction
Female breast cancer is one of the most challenging diseases of the current era owing incomplete knowledge about the exact mechanism(s) of causation and an unavailability of effective therapeutics. Breast cancer is the commonest cellular and molecular disorder among women in the world. 1 It is second to lung cancer in incidence, with the involvement of molecular processes that lead to deregulation of normal division of cells. 2 In 2012, the occurrence ratio of breast cancer was 1.7 million in the world; this level reflected an augmented rate of incidence and mortality of >20% and 14%, respectively, since 2008 values. It is the most commonly diagnosed cancer in 140/184 countries and cause of death (522,000 in 2012) among women. It now represents 1 in 4 cases of all cancers in women. 3 The current research was carried out to evaluate the role that potential hIFNα-2b gene mutations may have in breast cancer patients and to identify if changes could be associated as risk factor with the pathology.
The breast cancer incidence in young women varies widely in various Asian countries. In Pakistan, the most recent epidemiologic data indicated a 38.5% incidence and mortality rate of 25.7%; interestingly, in previous years, the most common cause of cancer death among women was cervical cancer. 4,5 The incidence of breast cancer in Karachi (Pakistan) was 69.1/100,000, with those at Stages III and IV being quite common. 1 Not surprisingly, the levels noted in Karachi were the highest for breast cancer incidence in any Asian population evaluated. 6
Many epidemiological, clinical, genetic, and molecular studies have identified some risk factors for breast cancer. 7,8 The occurrence of several common cancers is largely due to interactions between genetic and environmental factors that may cause changes in expression of genes that, in turn, might lead to changes in the structure/function of the body. 9,10 The genes usually associated with carcinoma of the breast are tumor suppressor genes (TSGs) and oncogenes. The most prevalent are C-erb-B2 or (HER2/neu) and C-myc and PRAD1, while the key TSGs are BRCA-I and BRCA-II, p53, RB and APC. The C-erb-B2 or (HER2/neu) and C-myc are the most frequently reported genes. Myc proteins (i.e., C-myc, L-myc, and N-myc) are nuclear phosphoproteins that play roles in differentiation, mitogenesis, and cell proliferation; changes in these proteins often result in malignancies in humans. 11 Patient genomic data—in correlation with risk factors—may provide information that can be utilized for better and specific treatments of these diseases early enough to yield a better prognosis. 12
Interferons (IFNs), apart from their abilities to exert pleiotropic effects on the body through multiple signaling pathways, might also play a significant role in breast cancer. Type-I IFNs have common receptors IFNAR1 (interferon α receptor 1) and IFNAR2 (interferon α receptor 2) are transmembrane proteins 13,14 binding to which results in interactions with tyrosine kinases tyk-2 (IFNAR1) or Janus-Kinase (JAK)-1 (IFNAR2) and resultant phosphorylation of signal transducer and activator of transcription (STAT) transcription factor. The STAT-1 and STAT-2, along with a non-STAT transcriptional factor p48 (IRF9-interferon regulatory factor 9), form ISGF-3 (IFN-stimulated gene factor-3). The ISGF-3 complex moves to the nucleus and binds IFN-stimulated response elements to prompt gene transcription. 15,16 In addition to IFN protein and receptors, there are various (>50) proteins involved in IFN signal transduction pathways, including hIFNα-2b. 17 –20
A number of roles attributed to IFN include regulation of cell multiplication, antiviral and antiangiogenic events, cell differentiation, and immune function in humans. 16,21,22 IFNα is used in the treatment of several cancers (e.g., hairy-cell leukemia/lymphoma, bladder cancer, Kaposi's sarcoma, AIDS-related leukemia, chronic myelogenous leukemia, hairy-cell non-Hodgkin's melanoma, multiple myeloma, as well as renal, cervical, and thyroid cancers. 23,24 These therapeutic roles for IFN in a variety of malignant and nonmalignant disorders have opened new horizons for the development of cytokine-based therapies in humans. It is expected that the potential use of hIFNα-2b can also be demonstrated. The current study first analyzed a group of risk factors and assessed clinical history for a selected group of breast cancer patients. Next, the authors screened, classified, and investigated 60 biopsy samples from breast cancer patients, with respect to gene expression of hIFNα-2b, an important negative modulator of cell growth, mediator of tumor cytotoxicity, and stimulator of immune response, thereby assuming an endogenous cancer-fighting role and a potential therapeutic trigger. Moreover, the authors analyzed for sequence mutations within the polymerase chain reaction (PCR) (positive) samples and further assessed their hIFNα-2b ligand-receptor binding capacity (Docking).
Materials and Methods
Data and sample collection
Breast tumor tissue samples (N = 60) were obtained from the Sir Ganga Ram Hospital, Lahore, Pakistan. Detailed discussion with oncologists and molecular biologists were carried out to formulate a schedule and to obtain interviews with patients diagnosed. A formulated questionnaire was generated based on different parameters to ascertain general information related to the patient's current and previous health statuses with informed consents and also information that could be more pertinent to breast cancer patients. A total of 60 breast cancer patients were eventually enrolled in the study, along with a control group (healthy individuals; N = 10) with medical history. Age-matched controls were selected to also have the same educational and socioeconomic backgrounds as the cancer patients.
Molecular Analysis
Sample processing and PCR analysis
Fresh surgically-removed tumor tissue samples (50 mg) from each patient were obtained which were pathologically confirmed by histopathology at the collaborative hospital. All samples were processed to isolate genomic DNA using commercially available kits (Fermentas, Waltham, MA). The isolated DNA was then subjected to electrophoresis over a 1% (w/v) agarose gel containing ethidium bromide (0.5 μg/mL), and fragment lengths were estimated by comparison against a 1 kb DNA ladder run in each gel (Vivantis Technologies, Kuala Lumpur, Malaysia).
For the analyses, two different sets of primers (IFN1F and IFN2R and NIF and N2R) were used 25 (Table 1). The complete hIFNα-2b gene along with the signal peptide region (695 bp) was amplified from human genomic DNA using primer sets and then subjected to PCR. An internal primer set was used to amplify only the encoding region of mature hIFNα-2b (625 bp). The amplified hIFNα-2b gene products underwent a gene-clean procedure using a QIAquick Gel Extraction Kit (QIAGEN, Düsseldorf, Germany) before sequencing.
Sequence and structural analysis
Of the 60 breast tumor samples, 38 with positive hIFNα-2b gene amplification underwent further sequencing. Sequencing products were analyzed using Chromas 2.33 software for peak correction, ClustalW 2.0 for multiple sequence alignment, and ExPASy for translating the DNA sequence into protein to study the changes in the derived proteins at the amino acid level. The 3-D structural models of the derived modified hIFNα-2b proteins were drawn by considering changes at the amino acid level. An automated mode of ‘Swiss Model’ was used, in which the model automatically searches templates from the database ExPDP and generates a homology model. The structural models of each hIFNα-2b variant were docked with human interferon α-2b receptor (IFNAR) protein using a protein–protein docking program Z-DOCK 3.0.2, and protein–protein interactions were evaluated using PDBSum.
Statistical analysis
A Chi-square test was applied to judge relative significance (p < 0.050) between PCR positive out of total and PCR negative samples out of total. Similarly, Chi-square test is applied on different parameters described in Table 2 for the same purpose.
IDC (Grade II): Samples 1–40; IDC (Grade III): Samples 41–47; DCIS (Grade I): Samples 50–51; DCIS (Grade 0): Samples 48, 49, 52–60.
DCIS, ductal carcinoma in situ; IDC, invasive ductal carcinoma.
Results
Patient information
The clinical and risk factors (Table 2) were analyzed in Pakistani women during 2010–2012. Sixty breast cancer patients of age 31–50 years, whose chemotherapy treatment had not yet commenced, were registered along with healthy controls. In seven patients (11.7%), diabetes, anorexia, hepatitis C, tuberculosis, and hypertension were observed. The time of initial diagnosis before surgery was within 6 months in 75% of the patients, the largest number of patients (92%) did not report any family history of breast cancer, 42% had their menstrual cycles > twice per month, and 53% had one to three children with lactation periods <1 year. Invasive ductal carcinoma (IDC) was found in 78.3% breast cancer patients, and ductal carcinoma in situ (DCIS) was found in 21.6% of the patients. Grade-II tumor formation was observed in 66.7% of the patients compared to 11.7%, which was classified as Grade-III.
There existed a significant difference (p = 0.000) between “time of initial diagnosis before surgery” between 0.6 years and greater than 0.6 years (Chi-square = 15.00; df = 1). There existed a significant difference (p = 0.000) between positive “family history of breast cancer” and negative “family history of breast cancer” (Chi-square = 41.667; df = 1). There existed a significant difference (p = 0.000) between menstrual cycles: 1 per month, 2 per month and >2 per month (Chi-square = 24.00; df = 2). There existed a significant difference (p = 0.000) between “family marriage” and “nonfamily marriage” (Chi-square = 38.000; df = 1). There existed a significant difference (p = 0.000) between cancer type IDC and cancer type DCIS (Chi-square = 19.267; df = 1). There existed a significant difference (p = 0.000) between tumor grades I, II, and III (Chi-square = 52.204; df = 2).
Molecular analysis
Breast tumor tissue samples were used to extract genomic DNA and an (∼15 kb) intact genomic DNA band was noted in all samples. Among the 60 samples, 38 (63.3%) were found positive in PCR for the hIFNα-2b gene. There existed a significant difference (p = 0.039) between positive PCR and negative PCR samples out of total (Chi-square = 4.267; df = 1). Positive products were subjected to genetic sequencing and aligned using ClustalW software. Normal hIFNα-2b gene sequences were amplified from each of the 10 healthy controls, and a consensus sequence was generated. Figure 1 shows PCR amplifications of the hIFNα-2b gene using IFN1F and IFN2R primers, while Figure 2 shows amplification products using the N1F and N2R primers.

PCR amplification of human interferon α-2b (hIFNα-2b) gene using primer set IFN1F and IFN2R. 1–3, PCR of breast cancer patient's biopsy samples; M, gene ruler DNA ladder mix (SM00331); PCR, polymerase chain reaction.

PCR amplification of hIFNα-2b gene using internal primer set N1F and N2R. 1–6, breast cancer patient's biopsy samples; M, gene ruler 50 bp DNA ladder (SM 0371).
The amino acid changes in human interferon α2b (hIFNα-2b) gene proteins from different breast cancer patients' samples were more frequent after amino acid position 140 as determined by alignment analysis. Among all the breast cancer samples, samples 1–40 were in Stage II and samples 41–47 were in Stage III and were IDC; breast cancer samples 48–60 were DCIS and all were in Stage 0 except for samples 50 and 51. The genetic changes through natural mutations (Table 3) in different isolates showed that in most cases the replacement was by an adenine (A) base leading to base mismatch and alterations in protein structure/conformation. Sequence data analysis through multiple sequence alignment showed that most of the mutations were in fact expressed as changes in amino acids. The various amino acid changes in hIFNα-2b proteins, with respect to base changes, are summarized in Table 4.
DCIS: Sample no. 48–60; IDC: Sample no. 1–47.
At position 141, there was a frameshift mutation that would result in a negatively-charged amino acid being replaced with a polar amino acid (E→G) that, in turn, would yield a nonfunctional protein. A mutation in the Patient 49 sample showed there would be a new strong binding pattern with receptor, and hence, this tumor was not developed much (i.e., at Stage 0); in contrast, while in samples from Patients 46 and 47, no binding of hIFNα-2b proteins with IFN receptors was observed and so tumor in these patients was then expected at a more advanced Stage III. In another case, at position 107, a silent and missense mutation was observed that coded for both similar amino acid (E→E) and different amino acid (E→K) sequences in Patient 28 (Stage II) and Patient 46 (Stage III), respectively. In addition, at position 145, during translation, a nonpolar amino acid would be replaced with the polar one (A→T), resulting in a nonleaky mutation that would not be associated with any reduction in protein activity. In samples from Patient 28 (Stage II) and Patients 46–48 (Stage III) of IDC, at position 146, negatively charged amino acids would be predicted to be replaced with a positively-charged amino acid (E→K); this missense mutation would result in protein structural and functional alterations and so likely give rise to more aggressive tumors (Grades II–III).
hIFNα-2b structural and functional analysis
A multiple sequence alignment of hIFNα-2b protein showing mutations at the amino acid level (read by BioEdit software) is showed in Figure 3. The 3-D structures of all the variants of hIFNα-2b modeled by “Swiss Model” were compared by superimposition with each other (Fig. 4). The comparisons that showed no significant changes in protein fold level were apparent. In contrast, there were local structural/conformational changes due to changes in intrachain interactions among amino acids (Fig. 5).

Multiple sequence alignment of hIFNα-2b protein showing mutations at amino acid level read by BioEdit software.
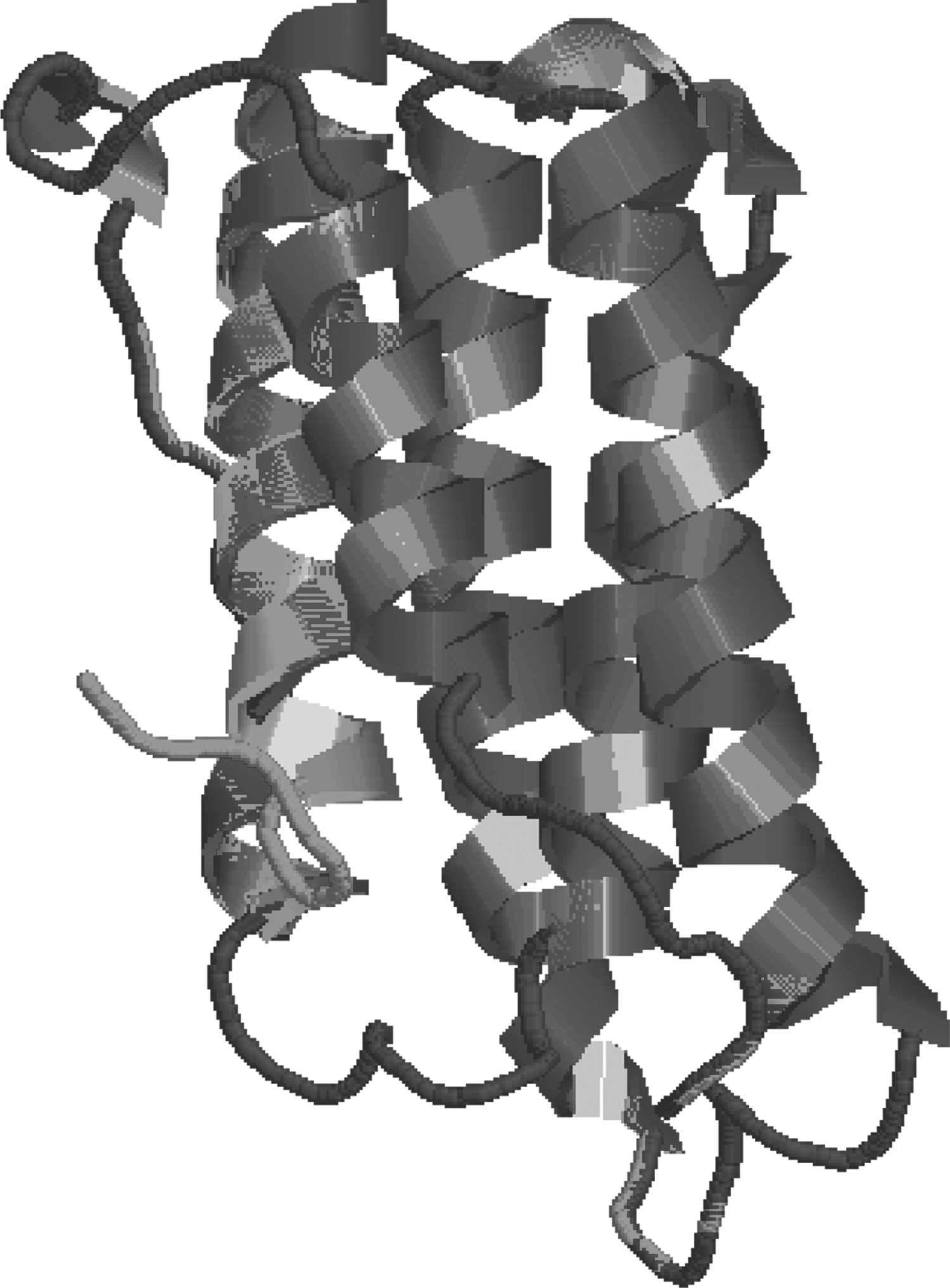
Protein structural model showing mutation at general fold pattern changes by superimposition of 19 hIFNα-2b variants.
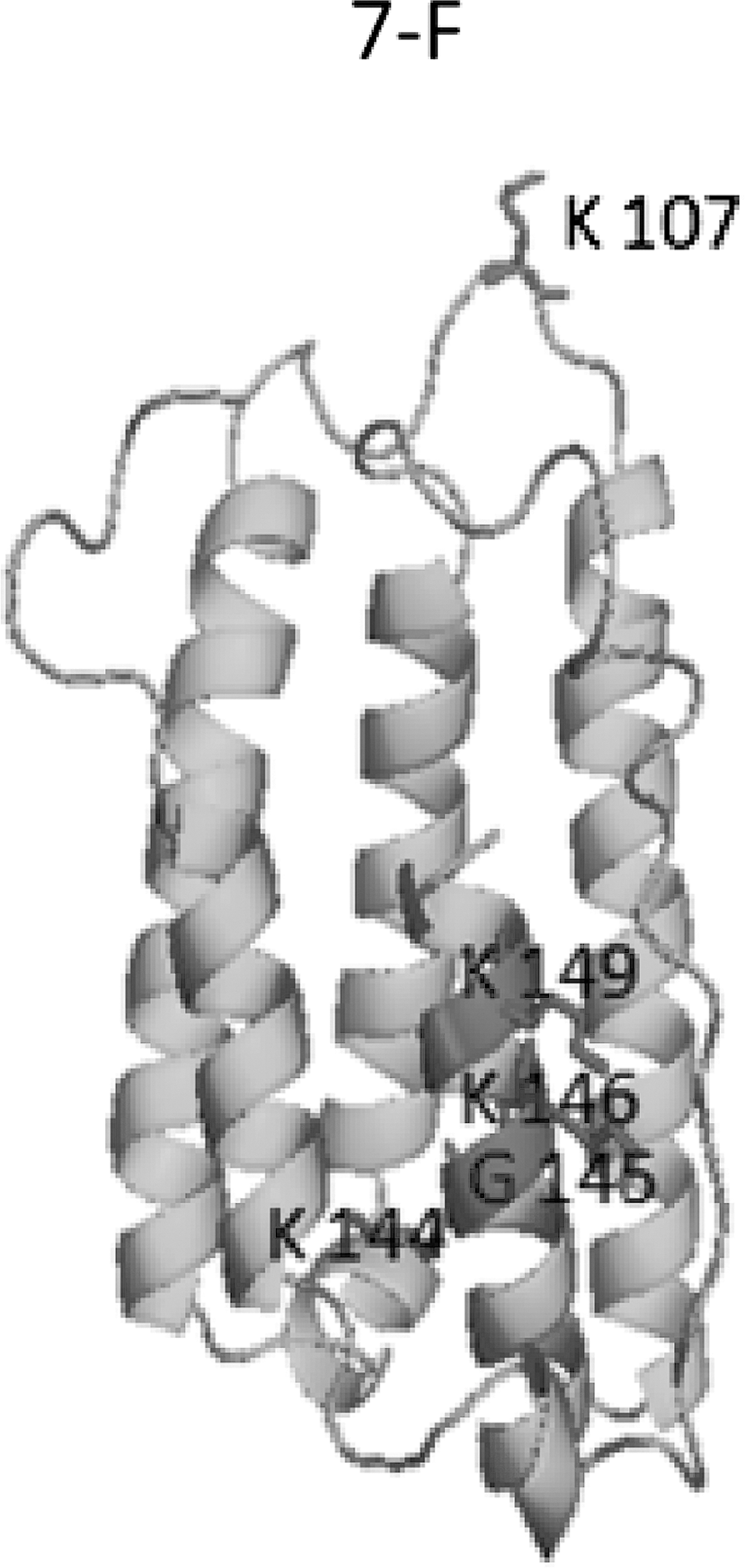
Protein structural model showing mutation at local level predicted by PyMOL as in product IFN-7F in cartoon style. The amino acids indicated with numbers are mutated.
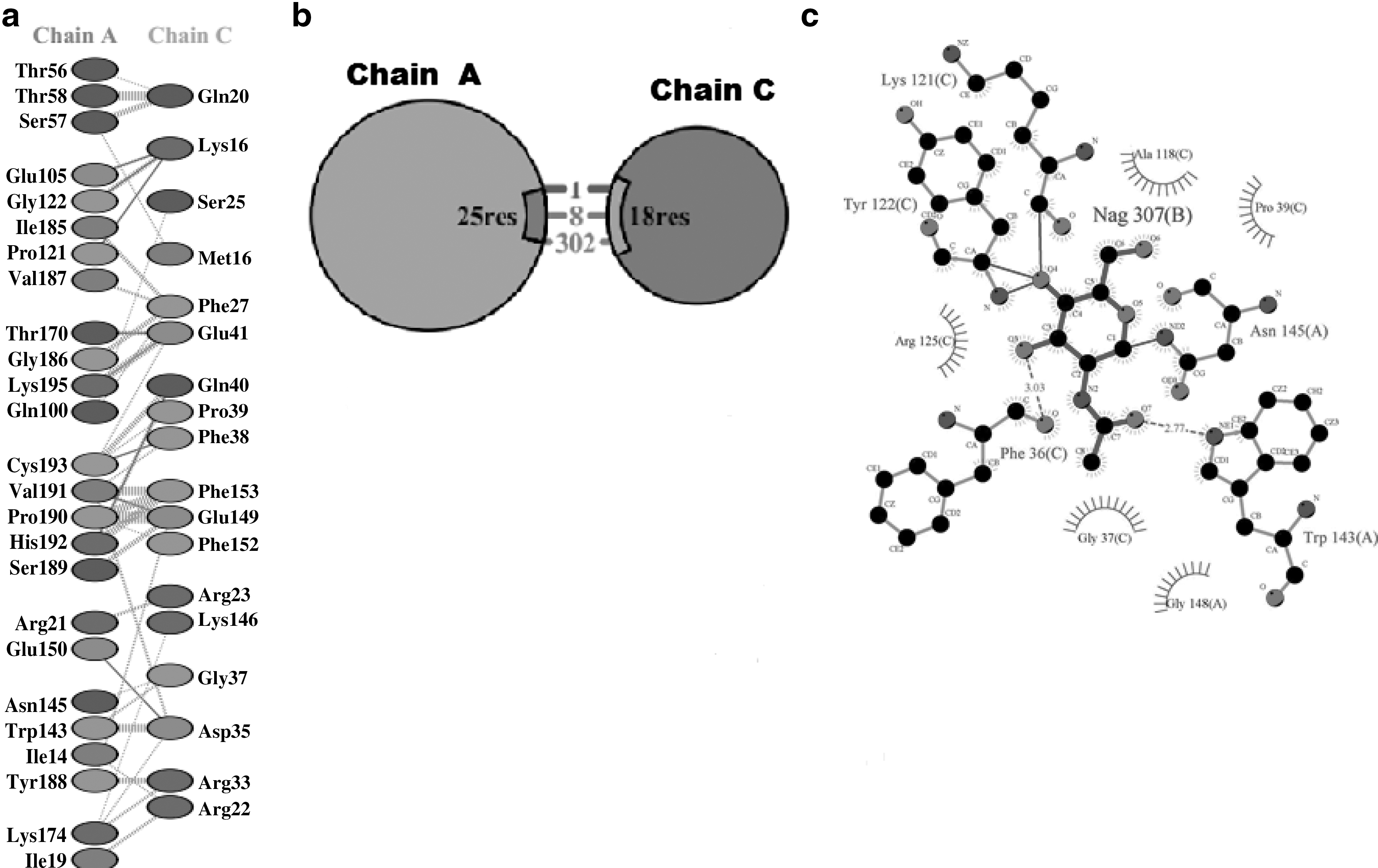
hIFNα-2b protein sequences were obtained through “PDBsum,” and docking results of normal interactions with mutated products were then compared (Fig. 6). For this, the protein models were submitted to Z-DOCK (along with IFNARI), receptor-complexes were generated, and interactions analyzed. “PDBsum” data included protein–surface interaction, protein–protein interactions of each sample with amino acid residues, numbers of hydrogen bonds, nonbonded interactions, disulfide bonds, and interface area. Interaction analyses of hIFNα-2b with IFNAR1 (using “PyMOL” and “PDBsum”) indicated that one hIFNα-2b variable (i.e., 49F) had very strong binding with a GlcNAc (N-acetyl-glucosamine) moiety on IFNAR1 (Fig. 7) in addition to interchain interactions of the two protein residues; all the remaining variants showed equivalent binding patterns with the GlcNAc along with interchain interactions of hIFNα-2b-IFNAR1. Product 46F did not show any interaction of its hIFNα-2b with IFNARI (Fig. 8), indicating that this hIFNα-2b bound a single residue of GlcNAc only. Such a loss of hIFNα-2b-IFNARI interactions would inhibit signaling. The product 28F showed only protein–protein interactions between IFN and IFNAR1 among different amino acids of the two chains (Fig. 9).
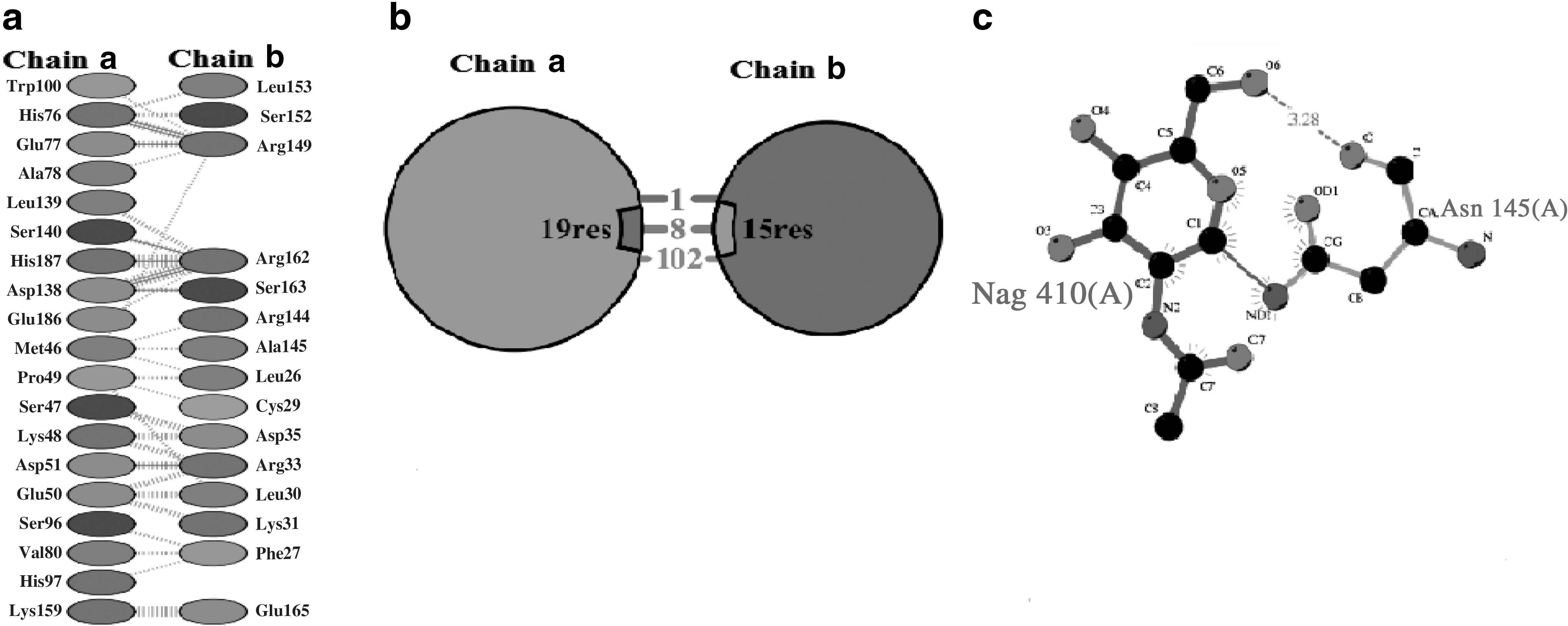
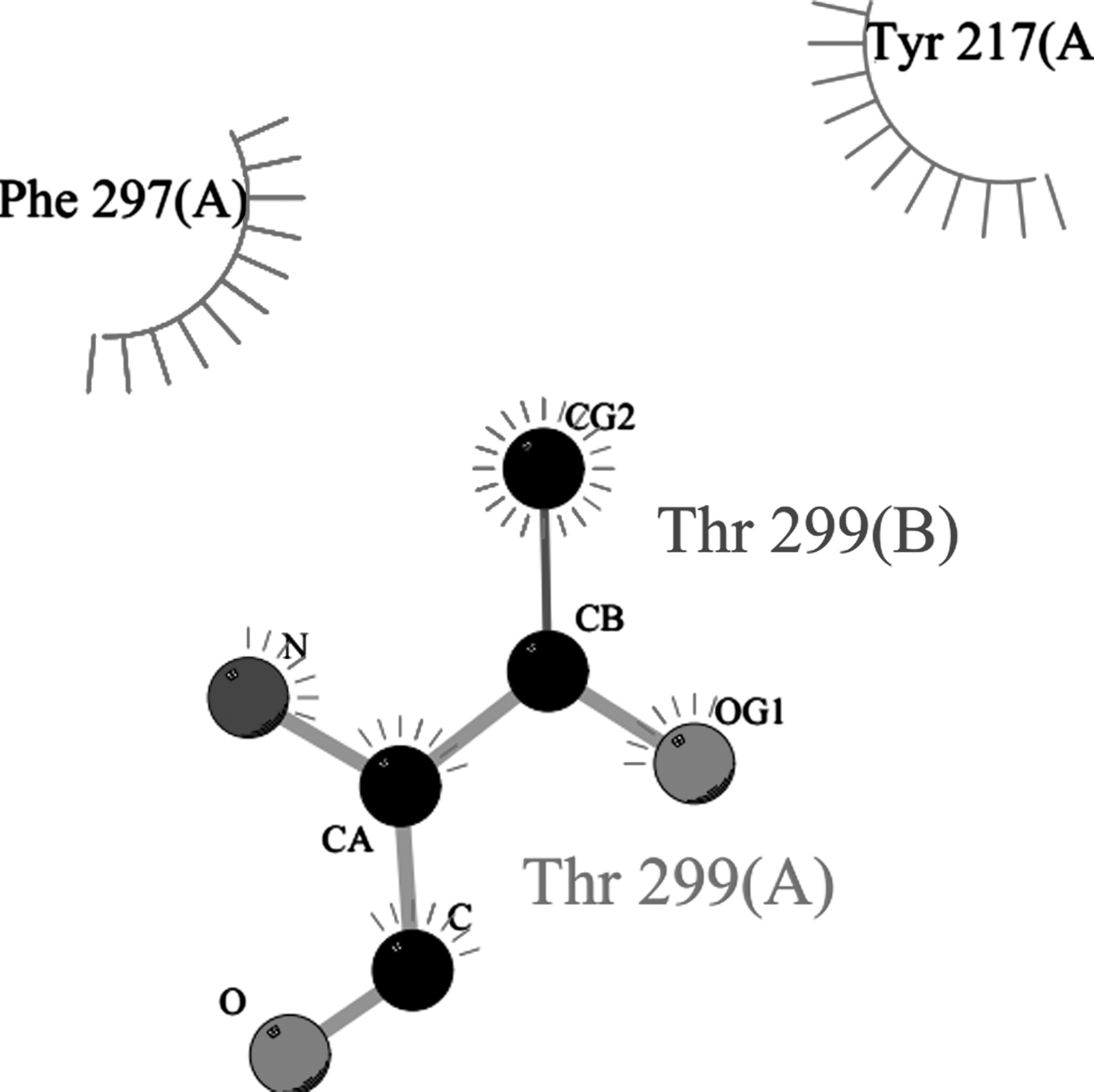
Product 46F hIFNα-2b showing no interaction of IFNAR1 with GlcNAc moiety of IFNα-2b predicted by “PDBsum”.
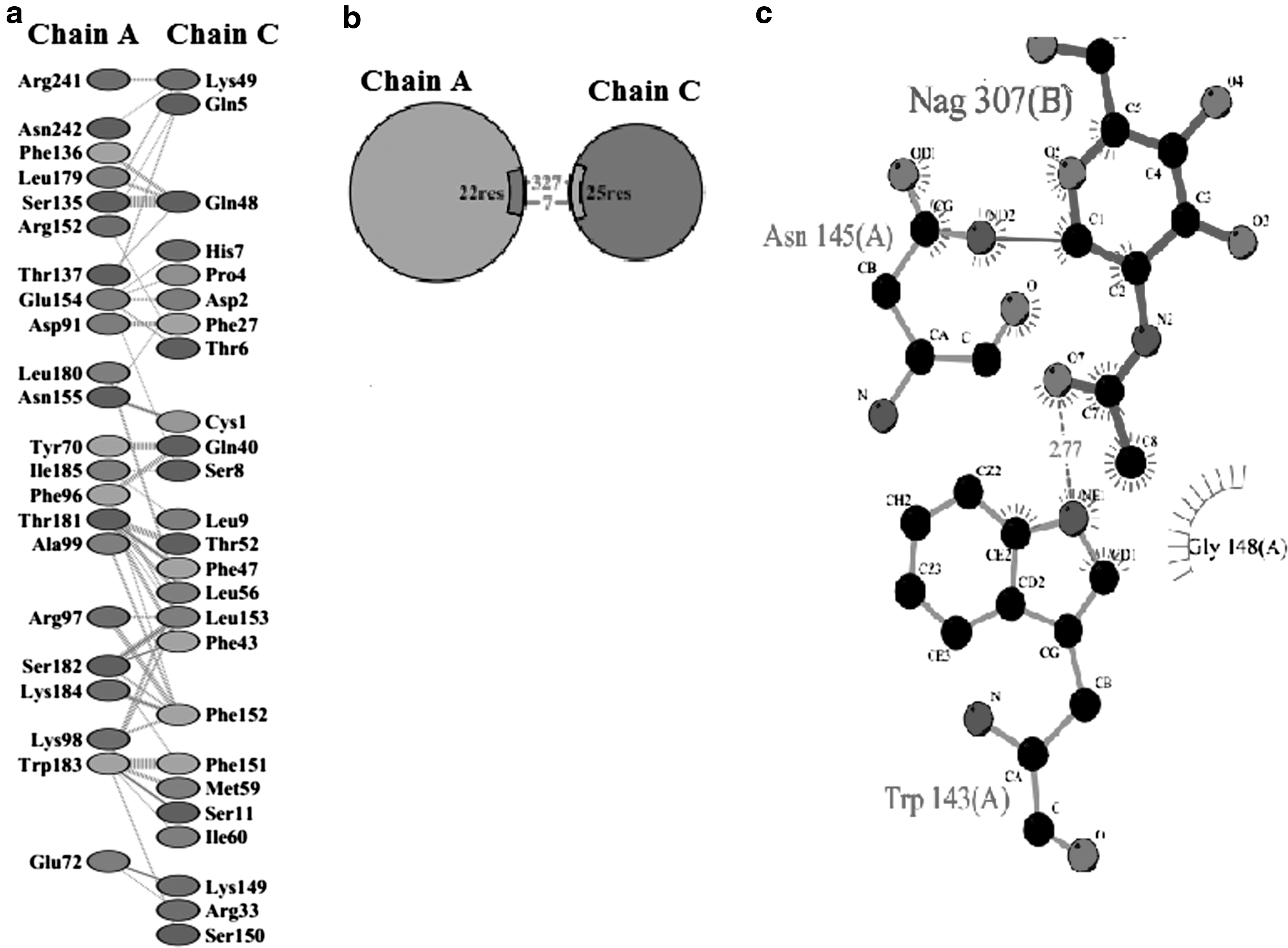
The product 28F showing only protein–protein interactions between IFN and IFNAR1 among different amino acids of two chains:
Discussion
Failure in eradicating a cancer is mainly due to a lack of identification of exact risk factors, the precise time of occurrence, and progression mechanisms in the body. In the current research, the initial stage related to the involvement of various risk factors was determined in 60 breast cancer patients whose tumor samples were subjected to molecular analysis and chemotherapy was not commenced. The patients were included in research during 2010–2012 along 10 healthy controls. The majority of breast cancer patients (78.3%) had IDC and 66.7% had Stage II tumors. Interestingly, 86.7% had marriages in the family, 63.7% had more than two menses per month, 58.3% had lactation periods <1 year, 91.7% had no family history of breast cancer, 53.3% had one to three children, and 68.3% were aged between 31–50 years. Clearly then, there is need for annual complete medical checkups, including mammography, for females especially more than 30 years of age if risk factors like those described in the current study are detected in the patient.
The present research sought to evaluate what role hIFNα-2b gene mutations could have in the onset/progression of breast cancer. Altered IFN responses have been reported to be a cause of immune defects in some cancer patients. 26 Furthermore, IFN signaling has an important role in the proper activation and homeostatic control of T cells, B cells, and NK cells; as such, impaired IFN signaling in cells may provide the required background to allow for increased survival of tumor cells in a host. In this study, of 60 breast cancer patient tumor DNA samples, 38 were found positive for the hIFNα-2b gene in PCR and were subjected to sequencing and aligned by ClustalW software in comparison with normal hIFNα-2b consensus sequence derived by sequencing of hIFNα-2b from 10 healthy individuals. The samples found negative indicated that there might be some deletion at the ends of this gene in some cancer patients as blood cancers; chromosomal deletions were reported on chromosome 9, where interferon gene cluster is present. 25,27
It has been postulated that the gene expression profile of a primary breast cancer patient could predict responses to some chemotherapeutics. 28 Alone, changed expression in TSGs (e.g., BRCA1, BRCA2, and p53) does not cause a high prevalence of breast cancers. 29 –31 Bouker et al. (2005) evaluated the role of interferon regulatory factor-1 (IRF-1) as a TSG in breast cancer cells to determine if suppressor function was modified so as to affect the cell cycle transition, apoptosis, or caspase activation, even in a manner that acted independent of p53 capacity to regulate apoptosis. 32
In the current study, single-strand changes in different positions were observed after 400 bp, seeming to result in drastic effects. Single-stranded mutations in hIFNα-2b gene have also been identified earlier in leukemia 25 and in brain cancers. 19 It was noted that in most of the cases, the normal bases were replaced by DNA base adenine, an outcome that altered the natural conformation of the gene and, ultimately, changed the final product amino acid. Out of 22 different types of mutations, in 11 mutations, an adenine base was involved—this base was replaced, deleted, or inserted, resulting in a change of amino acids in the hIFNα-2b gene protein in those breast cancer patients. These altered amino acid sequences might have suppressed the antiproliferative properties of hIFNα-2b protein and this finding is further supported by the observation of Staveley-O'Carroll et al. (1998) that in the development of cancer, an immune dysfunction is reported, which aids the metastatic lesion, and interferon is a key regulator of an immune system. 33 In other research, maloney leukemia virus-based retroviral vectors for expression of “murine angiostatin,” “endostatin,” and “IFN-α1” were generated and used to transduce breast cancer cell lines (MCF7 and MDA-MB435); a significant decrease in division of breast cancer cells was observed. 34 Accordingly, IFN-α1 and endostatin gene therapy might be useful for treatment of some breast cancers.
In the DCIS patients in the current study, two mutations are specifically present in the hIFNα-2b gene at DNA base positions 421 (G+) and 437 (G →A). These result in a frameshift of protein from 141 amino acid position and a change of 146 amino acid (E→K) in protein, respectively, while other mutations were absent in this breast cancer subtype. In IDC, these two mutations were also present along several other mutations (with a prevalence of 15% and 27%, respectively). In breast cancer IDC, mutations with a prevalence of 16.66% in the hIFNα-2b gene included the following: 319 (G)→(A), 321 (G)→(A), and 447 (G)→(A). Mutations with a prevalence of 11.66% included 418 (T)→(A) and 434 (G)→(A), while mutation 7 (C)→(T) had a prevalence of 10.0%. All remaining other mutations had <10.0% prevalence in the breast cancer patients in this study.
In the current study, at the amino acid level, protein predicted models were generated and superimposed upon each other to observe changes at the fold, as well as local level. There were no significant changes detected in the fold pattern, but frequent localized changes were seen. The analysis of docking results from PDBsum displayed that different variants of hIFNα-2b were almost in similar contact patterns in terms of contact surface area of the two proteins (hIFNα-2b and its receptor protein) and the number of residues involved in different types of binding interactions. For breast cancer patient 49 (grade 0), hIFNα-2b protein showed new binding contacts with receptor (IFNAR1) through its sugar moiety (GlcNAc) resulting in the enhanced binding strength of hIFNα-2b protein with the receptor and activation of the JAK-STAT pathway. The modified hIFNα-2b protein has a strong binding strength with receptor, and hence, it can be exploited clinically in cancer biotherapy. For breast cancer patient 46 (Grade III), docking results of “PDBsum” indicated that there was no binding of hIFNα-2b with IFNAR1 and seemed to be associated with loss of signal transduction pathway; hence functional impairment of hIFNα-2b was a likely result in those patients' cells.
These findings also suggest it is key to explore mechanisms behind the induction of signal transduction pathways and other inducing factors that may disturb the normal activity of hIFNα-2b protein. Identification of multiple signaling pathways activated through IFN receptors has dramatically advanced knowledge of cellular/molecular mechanisms, by which IFN facilitates the effects in situ. It is known from cancer prevention studies that selective amplification of the immune system may help reduce tumor growth. 35
Various immunotherapeutic strategies are still in use, and advances related to the cancer patient's abilities to overcome the immunosuppressive effects from, and obstacles generated by, the cancer itself continue to be made. 36 –41 Various IFNα and IL-2 therapies have been in use for more than 2.5 decades 42 and it is known that an efficient signal of IFN is an important event to help initiate full activation of lymphocyte clonal expansion and efficient NK cell-mediated cytotoxicity. 43 An altered IFN response is reported to be a cause of an immune defect in various cancer patients. 26 Furthermore, IFN signaling plays an important role in the proper activation and homeostatic control of T cells, B cells, and NK cells; as such, impaired IFN signaling in cells may provide the needed background to allow for increased survival of tumor cells in a host. Impaired signaling of IFN was found in breast cancer patients (at Stages II, III, and IV), 44 and same results were observed in breast cancer patients who were treated with neoadjuvant and/or adjuvant chemotherapy and in patients who had not received systemic therapy. Impaired IFN signaling may pose problems to those therapeutic approaches that include stimulating antitumor immunity. 26,44,45
Conclusions
Interferons are significant managers of many biological functions in humans and outstanding pharmacological agents for numerous diseases, including viral infections and malignancies. Molecular analyses in this study showed that in many of the biopsy samples, the hIFNα-2b gene after 400 bp position exhibited frequent alterations (i.e., replacements with adenine base). Most frequent mutations at the genetic level—when subjected to different bioinformatics tools—confirm their implications both at the sequence and structural level of producing proteins. In this context, the docking results were executed and their results depicted that all samples of hIFNα-2b have shown binding with IFNAR1 without sugar moiety except one (49F), which showed novel binding of hIFNα-2b with IFNAR1 through its sugar moiety (Asn 145) in addition to protein–protein interactions, while the product “46F” showed no binding with IFNAR1. From these results, it can be deduced that specific mutational variants of hIFNα-2b can be the risk factors sideways to genetic, as well as environmental risk factors, which may be associated with breast cancer. Clearly, it is critical to explore new molecular markers, TSGs, and oncogenes that might be important for understanding the etiology/progression of breast cancer. The authors' findings showed that there would likely be an impaired IFN response by some of the breast cancer patients' cells and hence an immune dysfunction during any immunotherapy. Such molecular studies can lead to the discovery of novel drugs that could be helpful with regard to immunotherapies to selectively enhance immune response in cancer patients.
Footnotes
Acknowledgments
The authors thank the Fatima Jinnah Medical College (Lahore) and the Sir Ganga Ram Hospital (Lahore) for providing all the biopsy samples used in these studies.
Disclosure Statement
No competing financial interests exist.