
Abstract
Drug resistance is extremely common in nonsmall cell lung cancer (NSCLC) and is one of the major problems in NSCLC chemotherapy. However, the detailed mechanisms remain largely unknown. Unfolded protein response (UPR) is involved in the tumorigenesis of NSCLC. Here, the authors demonstrated that the UPR promotes poly (ADP-ribose) polymerase activation (PARP) cleavage in NSCLC cells on doxorubicin treatment, which is a hallmark of apoptosis and caspase activation. In NSCLC cells, doxorubicin treatment triggers the UPR activation, which subsequently promotes doxorubicin-mediated apoptosis. Importantly, mild endoplasmic reticulum stress precondition enhances the sensitivity of NSCLC cells to doxorubicin-initiated apoptosis. Furthermore, the eukaryotic translation initiation factor 2α (eIF2α) branch of the UPR is involved in the synergistic role of the UPR in NSCLC cell apoptosis on doxorubicin treatment. They also demonstrated that the mTOR pathway plays an essential role in synergistic induction of apoptosis by the UPR and doxorubicin in NSCLC cells. Taken together, these results provide a potential mechanism that the UPR promotes doxorubicin-induced apoptosis in NSCLC cells, at least in part, by eIF2α-mediated mTOR signal inactivation.
Introduction
Lung cancer is one of the most common cancers worldwide and is the leading cause of cancer morbidity. 1,2 Lung cancer has received considerable attention in recent years because of its rapidly increasing incidence. Lung cancer can be categorized into small-cell lung cancer, and nonsmall cell lung cancer (NSCLC). NSCLC is the most dominant form of lung cancer, accounting for more than 80% of this cancer. 3 Patients diagnosed with lung cancer have a poor prognosis because of the aggressive features of patients with NSCLC. 4,5 The traditional treatment for lung cancer includes surgery, radiotherapy, chemotherapy, and their combination. 6 Chemotherapy has become a recommended treatment for NSCLC. However, the development of drug resistance to chemotherapeutic agents in NSCLC cells is the major cause for failure of chemotherapy for NSCLC. 7,8 Doxorubicin is a widely used anticancer agent against a broad range of tumors, such as breast cancer, and hepatocellular carcinoma. 9,10 However, the clinical application of doxorubicin in lung cancer chemotherapy is limited by acquired or intrinsic resistance of cancer cells to this drug. 11 Therefore, there is an urgent need to enhance the efficiency of doxorubicin therapy of lung cancer.
The endoplasmic reticulum (ER) is a dynamic organelle responsible for the synthesis and maturation of secreted, membrane-bound, and some organelle-targeted proteins. Perturbation of ER homeostasis leads to the accumulation of unfolded or misfolded proteins, and triggers the unfolded protein response (UPR). 12,13 The activation of UPR is believed to alleviate ER stress and promote cell survival. 12 –14 Accumulating evidence showed that the UPR is related to the sensitiveness of cancer cells to chemotherapeutic agents, making them either more sensitive in some cases or more resistant in others. 15,16 Several studies have indicated a potential link between the UPR signal pathways and cisplatin resistance in NSCLC cells. 17,18 Whether the UPR plays different roles in the sensitiveness of NSCLC cells to different chemotherapeutic agents is largely unknown.
In this study, the authors investigated the roles of the UPR in the sensitiveness of NSCLC cells to doxorubicin. They found that the UPR promotes PARP cleavage, suggesting apoptosis of NSCLC due to doxorubicin treatment through the mTOR pathway.
Materials and Methods
Materials
Doxorubicin (Dox), tunicamycin (Tun), and tauroursodeoxycholate (TUDCA) were purchased from Sigma Chemical Company. Salubrinal (Sal) and rapamycin (Rap) were purchased from Selleck Chemicals. Antibodies against PARP, phospho-eIF2α (Ser51), eIF2α, phosphor-RPS6, and β-actin were purchased from Cell Signaling Technology.
Cell culture and treatment
Human NSCLC cell lines A549 and PC9 were maintained at 37°C in a humidified incubator containing 5% CO2, in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin/gentamicin. Tunicamycin (0.5 μg/mL) was used to induce mild ER stress response. The cells were pretreated with 100 μM TUDCA, a specific inhibitor of ER stress, or 25 μM salubrinal, a specific inhibitor of eIF2α dephosphorylation, for 1 hour before doxorubicin (1 μM) treatment.
Western blot analysis
A549 and PC9 cells were harvested in the appropriate buffer for the purpose of each experiment. Protein concentrations were measured using the BCA assay (Santa Cruz, CA). Equal proteins (60 μg) were applied to sodium dodecyl sulfate/polyacrylamide gel electrophoresis. After electrophoresis, proteins were blotted to polyvinylidene fluoride membranes and then blocked with 5% skim milk powder with 0.1% Tween-20. The blots were then probed at 4°C overnight with the relevant primary antibodies, washed three times with TBST (TBS containing 0.1% Tween-20), and probed with the appropriate horseradish peroxidase-conjugated secondary antibodies at room temperature for 2 hours. Immunoreactive material was detected using the ECL kit (Santa Cruz) according to the manufacturer's instructions.
RNA preparation and reverse transcription-PCR analysis
Total RNA was prepared as detailed previously. 19 Reverse transcription was performed with 0.2 μg of isolated total RNA using the M-MLV reverse transcriptase kit (Promega). Actin was used as a normalization control. PCR products were run on 1% agarose gel. The primers used in this study were as follows: GADD153-forward AGT CAT TGC CTT TCT CTT CG, GADD153-reverse GGT GCA GAT TCA CCA TTC GG; Actin-forward ATC TGG CAC CAC ACC TTC TAC, Actin-reverse CAG CCA GGT CCA GAC GCA GG.
Statistical analysis
Results are expressed as mean ± standard deviation. Statistical analysis was performed using Student's t-test. p < 0.05 was considered statistically significant.
Results
Doxorubicin initiates NSCLC cell apoptosis
To investigate the potential role of doxorubicin in the induction of apoptosis in human NSCLC cells, A549 and PC9 cells were used to investigate the cytotoxicity effects of doxorubicin. Western blot analysis revealed that doxorubicin treatment induced the cleavage of PARP in A549 and PC9 cells in a time-dependent manner (Fig. 1A, B). These data suggest that doxorubicin can initiate NSCLC cell apoptosis.
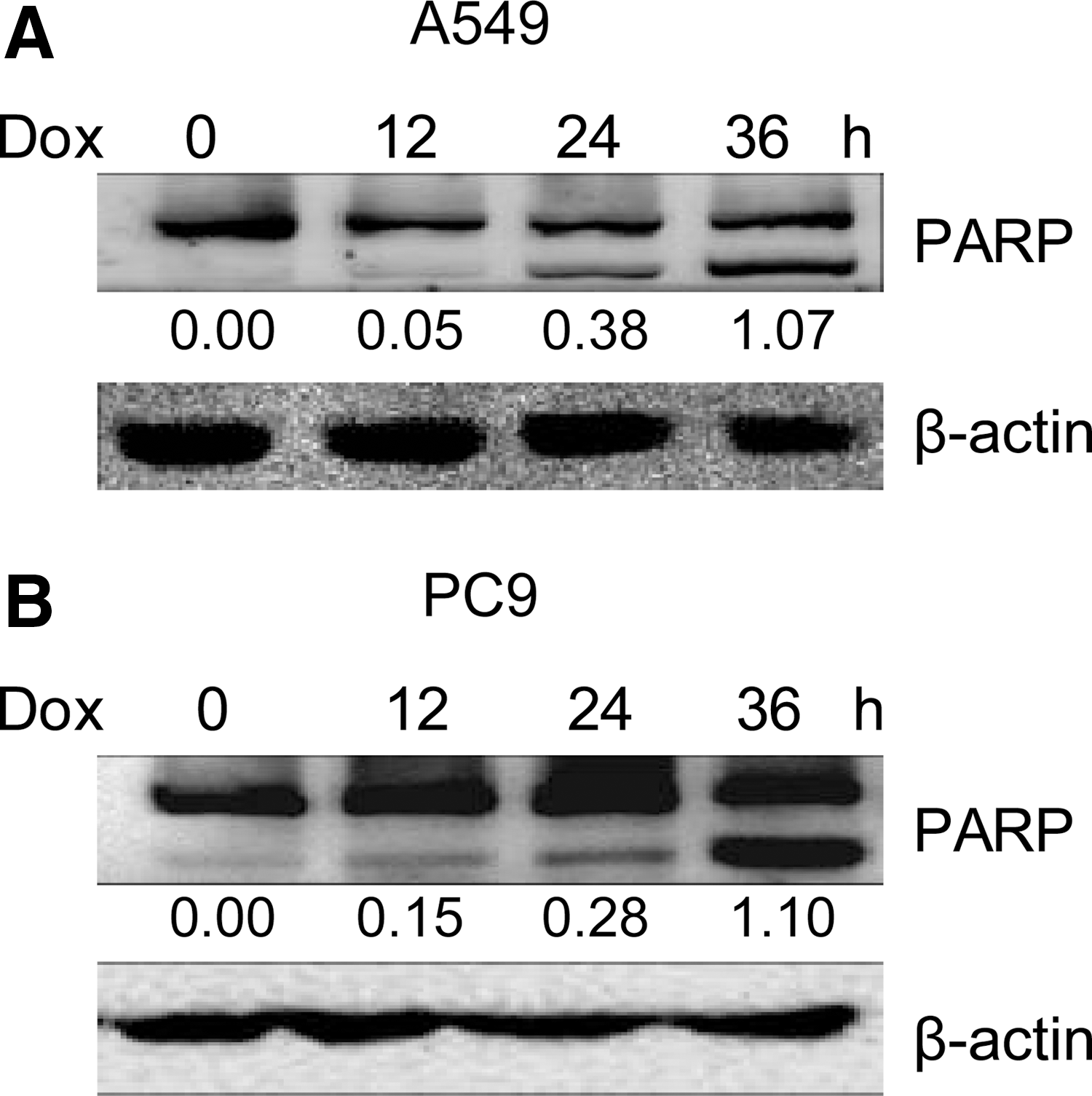
Doxorubicin initiates A549 and PC9 cell apoptosis.
Doxorubicin initiates UPR in NSCLC cells
To examine the effect of doxorubicin on the UPR activation in human NSCLC cells, the protein levels of phospho-eIF2α were determined in doxorubicin-treated A549 and PC9 cells. The authors found that doxorubicin treatment increased the protein levels of phospho-eIF2α in a time-dependent manner in A549 and PC9 cells (Fig. 2A, B), indicating the UPR activation. In addition, the mRNA levels of growth arrest and DNA damage-inducible 153 (GADD153) were increased in both A549 and PC9 cell lines after doxorubicin treatment (Fig. 2C, D), confirming the UPR activation. Tunicamycin (Tun) that directly inhibits N-linked glycosylation was used to induce activation of ER stress in these studies, and here the authors use Tun as a positive control of ER stress response. These results indicate that doxorubicin induces the activation of UPR in NSCLC cells.

Doxorubicin induces the UPR in A549 and PC9 cells.
TUDCA inhibits doxorubicin-initiated NSCLC cell apoptosis
As the UPR plays a pivotal cytoprotective role under stress conditions, the authors examined whether the UPR can affect human NSCLC cell apoptosis on doxorubicin treatment. Figure 3A, B showed that the UPR inhibitor TUDCA pretreatment significantly inhibited doxorubicin-induced apoptosis in A549 and PC9 cells. These results reveal that the UPR is involved in promoting doxorubicin-mediated apoptosis in NSCLC cells.

TUDCA inhibits doxorubicin-initiated A549 and PC9 cell apoptosis.
The UPR precondition promotes doxorubicin-initiated NSCLC cell apoptosis
Since the UPR is obviously activated in lung cancer, 20 tunicamycin was used to establish the ER stress microenvironment in A549 and PC9 cells. Based on this mimicked ER stress microenvironment, the authors investigated whether the UPR precondition plays some roles in regulating doxorubicin-mediated human NSCLC cell death. The results showed that tunicamycin pretreatment increases the cleavage of PARP in A549 (Fig. 4A) and PC9 (Fig. 4B), which is a hallmark of apoptosis confirming that cells undergo apoptosis after doxorubicin treatment. These results indicate that the UPR can make NSCLC cells more sensitive to doxorubicin.
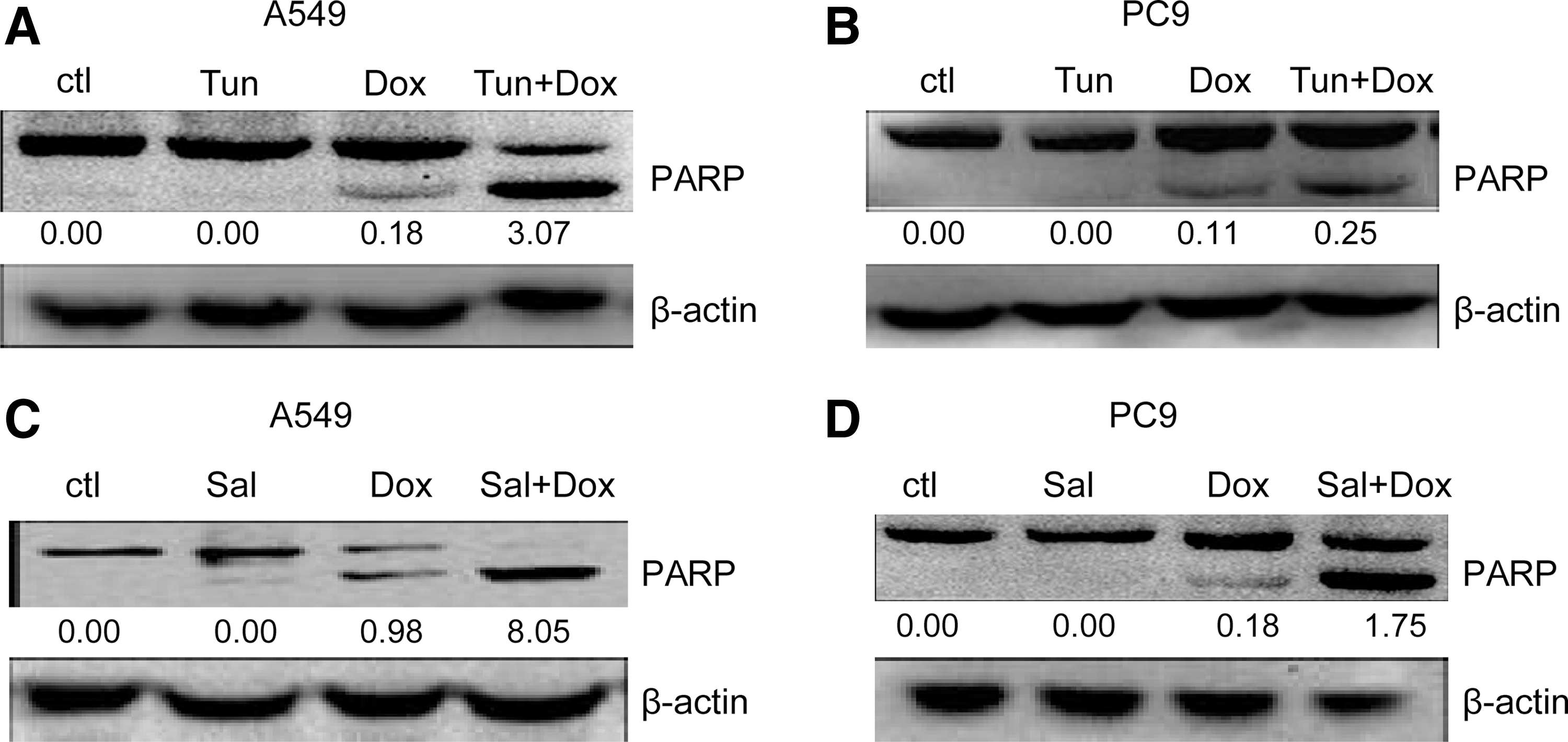
The UPR precondition promotes doxorubicin-initiated A549 and PC9 cell apoptosis.
Considering that the eIF2α pathway plays an essential role in the UPR signal system-induced apoptosis, the roles of eIF2α signal in the toxic effects of doxorubicin in human NSCLC cells were investigated. The data showed that salubrinal pretreatment enhanced A549 (Fig. 4C) and PC9 (Fig. 4D) cell apoptosis on doxorubicin treatment. Taken together, these data indicate that the eIF2α branch of the UPR pathways can enhance doxorubicin-mediated NSCLC cell apoptosis.
eIF2α/mTOR signal contributes to the synergistic effect of the UPR and doxorubicin
Further investigations were conducted to explore the mechanisms of eIF2α pathway in promoting doxorubicin-mediated human NSCLC cell apoptosis. These data showed that eIF2α pathway agonist salubrinal pretreatment inhibited the phosphorylation of ribosomal protein S6 (rpS6), a downstream effector of mammalian target of rapamycin (mTOR), in doxorubicin-treated A549 (Fig. 5A) and PC9 (Fig. 5B) cells. Moreover, mTOR inhibitor rapamycin pretreatment promoted doxorubicin-induced A549 (Fig. 5C) and PC9 (Fig. 5D) cell apoptosis. These results suggest that the synergistic function of the UPR in doxorubicin-treated NSCLC cells is mediated, at least in part, by the mTOR pathway inhibition.

eIF2α/mTOR signal contributes to the synergistic effect of the UPR and doxorubicin.
Discussion
Due to inadequate vascularization and rapid growth, tumor cells experience hypoxia and anoxia, which can lead to the accumulation of unfolded proteins in ER, resulting in UPR. 21 Accumulating evidence showed that hypoxia and anoxia play an important role in regulating the sensitivity of solid tumors to drug. Both hypoxia and anoxia can result in ER stress, initiating the UPR. It has been reported that the UPR is activated in lung cancer, but how this contributes to chemosensitivity in lung cancer cells remains largely unknown. 20 In this study, the authors demonstrated that the UPR promotes human NSCLC cell apoptosis through eIF2α/mTOR signal regulation on doxorubicin treatment.
Doxorubicin had been reported to initiate the UPR activation, and the UPR is related to the sensitiveness of cancer cells to this drug, making them either more sensitive in some cases or more resistant in others. 22 –24 In this study, the authors found that doxorubicin treatment resulted in phospho-eIF2α and GADD153 induction, both are indicators of UPR activation, in A549 and PC9 cells. Moreover, the findings that the blockade of UPR activation by the inhibitor TUDCA decreased the expression of a hallmark of apoptosis (PARP cleavage) strongly suggest that the UPR mediates doxorubicin-induced NSCLC cell apoptosis. Interestingly, they found the total PARP expression was downregulated by the TUDCA plus doxorubicin treatment, but not by either one of them, which may need to be further investigated. To confirm the synergistic role of the UPR in doxorubicin-mediated NSCLC cell death, mild ER stress was triggered before doxorubicin treatment. These data demonstrated that optimum concentration of ER stress inducer tunicamycin preincubation significantly enhanced doxorubicin-induced apoptosis in NSCLC cells. This is consistent with the recent study by Mihailidou et al. that Tun increases the anticancer efficacy of doxorubicin in tumor-bearing mice. 23 The authors therefore conclude that mild stress promotes UPR and consequently NSCLC cell death under doxorubicin treatment.
A recent report has revealed that the phosphorylation of eIF2a suppresses cisplatin-induced A549 cell apoptosis, and phospho-eIF2α inhibited cisplatin-induced A549 cell apoptosis, at least in part, by suppressing the p38 pathway. 17 They found that salubrinal, which induces the phosphorylation of eIF2α, improved the efficacy of doxorubicin in A549 and PC9 cells. Furthermore, these data showed that mild ER stress induced by tunicamycin inhibited cisplatin-induced A549 and PC9 cell apoptosis (data not shown). Mechanistically, these results suggest that the UPR is related to the sensitiveness of NSCLC cells to chemotherapeutic agents, making NSCLC cells either more sensitive in some cases or more resistant in others.
It is notable that combination treatment with salubrinal and doxorubicin inhibited the activity of the mTOR pathway in A549 and PC9 cells. Moreover, mTOR inhibitor rapamycin apparently promoted doxorubicin-induced apoptosis in A549 and PC9 cells. Based on these data, it is reasonable to suggest that the eIF2α pathway-mediated mTOR pathway inhibition is involved in the synergistic role of the UPR in doxorubicin-mediated NSCLC cell death. In a future study, the authors are going to do the in vivo experiment to confirm these in vitro data.
In conclusion, the authors reported that the UPR increases the sensitivity of human NSCLC cells to death on doxorubicin treatment. The synergistic role of the UPR under doxorubicin treatment is mediated, at least in part, by the eIF2α pathway activation. They also showed that the mTOR pathway plays an essential role in synergistic induction of apoptosis by the UPR and doxorubicin in human NSCLC cells. It is notable that these findings are now limited in NSCLC cells in vitro. Considering the high heterogeneity of NSCLC patients could result in differences in chemotherapy efficiency, whether a combination of UPR induction and doxorubicin can be used in NSCLC patient treatment needs personalized analysis. In the authors' opinion, response to combined UPR induction and doxorubicin may correlate with the mTOR signaling activation. Those patients with mTOR signaling overactivation may benefit from this therapeutic strategy. Further studies on the detailed mechanisms of combination treatment with salubrinal and doxorubicin-mediated mTOR pathway inhibition will contribute to the development of new therapeutic strategies against lung cancer.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (81472312), Innovation Team of Education Department of Sichuan Province (16TD0021), Luzhou City-Southwest Medical University Foundation (2013LZLY-J06, 2015LZCYD-S01-8/15, 2015LZCYD-S01-14/15), and Sichuan Province-Luzhou City-Southwest Medical University Foundation (14JC0082, 14JC0038).
Disclosure Statement
No conflicting financial interests exist.