
Abstract
Background:
Overexpression of TSPAN8 has been involved in several epithelial cancers and TSPAN8 can form a complex with a variety of proteins to participate in several import cellular functions. However, the effects of TSPAN8 in nonsmall cell lung cancer (NSCLC) remain unclear.
Materials and Methods:
In this study, the authors determined the expression of TSPAN in several NSCLC cell lines (95C, A549, H1299, and 95D) and human bronchial epithelial (HBE) cells. Furthermore, the authors investigated the biological function of TSPAN8 in NSCLC cell lines using gain-of-function and loss-of-function assays, as well as the underlying mechanisms.
Results:
TSPAN8 was found to be overexpressed in NSCLC cells compared with normal HBE cells, of which the expression in H1299 is the highest and, in 95C, it is relatively lowest. Functional assays indicated that knockdown of TSPAN8 in H1299 remarkably reduced cell viability and proliferation, while overexpression of TSPAN8 in 95C dramatically enhanced cell viability and proliferation. In addition, TSPAN8 knockdown led to G1 phase arrest and apoptosis by downregulating CDK2, CDK4, and Cyclin D1 and upregulating Bax and PARP.
Conclusions:
These results provide evidence that TSPAN8 may contribute to the pathogenesis of lung cancer by promoting cell viability and proliferation. TSPAN8 silencing may provide a potential therapeutic intervention for the treatment of NSCLC.
Introduction
Lung cancer is the main cause of cancer-related death especially in males worldwide, which accounts for about 13% of total deadliest cancers. 1 It has been classified as nonsmall cell lung cancer (NSCLC) and small cell lung cancer in most studies according World Health Organization (WHO) Classification of Tumors of the Lung. 2,3 Nevertheless, NSCLC subtypes, including 40% adenocarcinomas, 25%–30% squamous cell carcinomas, and 30%–35% large cell carcinomas, cover between 85% and 90% of all diagnosed lung cancers and the predicted 5-year survival rate is only 15.9%. 4 Chemotherapy, radiotherapy, and surgery are currently the two basic and major treatments to improve clinical outcomes, but overall survival remains poor and often leads to undesirable side-effects on normal cells or tissues. 5,6 Thus, a novel therapeutic target for lung cancer is needed as its incidence has been dramatically increasing in recent years.
Tetraspanins (TSPANs) are a superfamily of small proteins that consist of four transmembrane domains, three cytosolic domains, and two extracellular loops (SEL and LEL). They form complexes with a large variety of other transmembrane and/or cytosolic proteins to regulate several important cellular functions by building a network of interactions referred to as TSPAN webs. TSPANs actually act as molecular facilitators to stabilize, modulate, or inhibit activities of related molecules. 7 –9 TSPAN8 (also known as TM4SF3 or CO-029), a member of the TSPAN superfamily, is highly conserved from yeast to humans. 10 Studies have reported that the overexpression of TSPAN8 is correlated with several tumors, such as hepatocellular cancer, pancreatic cancer, colon cancer, esophageal cancer, prostate cancer, ovarian cancer, and cervical cancers. 11 –13
TSPAN8 plays a role in a variety of cancer cell functions, including cancer cell survival, migration, metastasis, and tumor angiogenesis. For example, a report has shown that TSPAN8 expression is associated with poor prognosis in colorectal cancer and similar phenomenon has been observed of esophageal carcinoma, hepatocellular carcinoma, and melanoma. 14,15 It has also been reported that TSPAN8 enhances esophageal cancer cell invasion through upregulation of the expression of disintegrin and metalloproteinase. 16 Moreover, TSPAN8 and CD151 have been reported to be essential exosome components to enhance tumor cell migration and invasion for the cross talk between cancer-initiating cells and their surroundings. 17 Several reports show that TSPAN8 is associated with bipolar disorder and schizophrenia too. 7 However, the role and relevance of TSPAN8 are yet to be investigated in lung cancer.
In this study, the authors found that TSPAN8 was overexpressed in human NSCLC. Herein, overexpression and knockdown of TSPAN8 are constructed in 95C and H1299 cells, respectively. The cell proliferation assay was performed in both 95C and H1299 cells. In addition, the cell cycle and apoptosis and possible downstream molecules were detected in H1299. The data demonstrate that TSPAN8 is very important for NSCLC cell viability and proliferation and it may be a new biomarker for treatment of lung cancer.
Materials and Methods
Cell lines and culture
Human NSCLC cells, 95-C, 95-D, A549, H1299, and human embryonic kidney 293T cells (HEK293T), were obtained from the Cell Bank of Chinese Academy of Science (Shanghai, China). Human bronchial epithelial (HBE) cells were obtained from American Type Culture Collection (ATCC). HEK293T cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Cat. No. SH30243.01B; Hyclone) supplemented with 10% fetal bovine serum (FBS; Cat. No. 04-001-1A; BI); 95-C, 95-D, A549, H1299, and HBE were maintained in 10% FBS 1640. All cells were cultured at 37°C in a humidified incubator containing 5% CO2.
Lentivirus-mediated short hairpin RNA knockdown of TSPAN8
The short hairpin RNA (shRNA) sequence targeting the TSPAN8 gene was 5′-AATCAGCGCATCTACTTCGAT-3′ and the negative control shRNA was 5′-TTCTCCGAACGTGTCACGTTTC-3′. The construction of plasmid, lentivirus packaging, and purification was based on the methods previously described. 18 H1299 cells (10,000 cells/well) were added into 24-well plates and transfected with lentivirus using 8 μg/mL polybrene (Sigma). Stable expression cells were filtered and screened in medium containing 800 ng/mL puromycin (Sigma). Silencing efficiency was estimated at messenger RNA (mRNA) and protein levels by reverse transcriptase–quantitative polymerase chain reaction (RT-qPCR) and western blotting.
The construction of TSPAN8 overexpression vector
The full-length TSPAN8 coding sequence was amplified from complementary DNA (cDNA) synthesized by RT-PCR using the total 5 μg RNA extracted from H1299 cells as the template. Briefly, cDNA was synthesized by reverse transcription using the oligo dT18 and subjected to PCR amplification of the full-length TSPAN8 coding sequence as the template. The primer sequences for TSPAN8 were 5′-GTCATCCGCGGAAAAGAC-3′ (the forward primer) and 5′-GGGTGTAAAAACGCAACTA-3′ (the reverse primer). The PCR products were purified by the Gel Extraction purification kit (OMEGA), digested with BamHI and NotI, and inserted into the pcDNA3.0 vector (Invitrogen, Carlsbad, CA) to obtain the recombinant plasmid TSPAN8
RT-PCR and qPCR
Total cellular RNA was extracted from NSCLC and HBE cells using TRIzol® reagent (Cat. No. 15596-018; Invitrogen). RNA quantity and quality were determined by spectrophotometry and agarose gel electrophoresis, respectively. Total 1 μg of extracted RNA sample was reverse transcribed into cDNA with the Bestar™ qPCR RT Kit according to the manufacturer's protocol. RT-qPCR was carried out with the DBI Bestar® SYBR Green qPCR Master Mix according to the manufacturer's instructions with regard to Agilent Stratagene Real-Time quantitative PCR Mx3000P Detection System. Primer sets used were as follows: for GAPDH, 5′-GTGGACATCCGCAAAGAC-3′ (forward) and 5′-AAAGGGTGTAACGCAACTA-3′ (reverse), and for TSPAN8, 5′-CCCAACTACTGCGACCAGAT-3′ (forward) and 5′-CCCGTCACCTCACATCATTC-3′ (reverse). The cycling conditions were initial denaturation at 95°C for 60 seconds, 95°C for 5 seconds, and 58°C for 20 seconds, 40 cycles. Data analysis was performed using the 2−ΔΔCt method.
Western blot
Cells were harvested and lysed in ice-cold RIPA (Cat. No. P0013B; Beyotime). Total protein concentration of the lysate was determined using the BCA Protein Assay Kit (Cat. No. 23235; Pierce Biotechnology). Thirty micrograms of protein in each lane was electrophoresed on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membrane (Cat. No. 162–0177; BioRad) and incubated with rabbit anti-TSPAN8 (Cat. No. 117515-1-AP, 1:1000 dilution; Proteintech), anti-CDK2 (Cat. No. 4668, 1:1000 dilution; Cell Signaling), rabbit anti-CDK4 (Cat. No. 9252, 1:1000 dilution; Cell Signaling), rabbit anti-Cyclin D1 (Cat. No. 9215, 1:500 dilution; Cell Signaling), rabbit anti-bax (Cat. No. 9212, 1:1000 dilution; Cell Signaling), rabbit anti-PARP (Cat. No. 9212, 1:1000 dilution; Cell Signaling), and rabbit anti-GAPDH (Cat. No. 10494-1-AP; 1:1000 dilution; Proteintech) antibodies overnight at 4°C. Membranes were then incubated with horseradish peroxidase (HRP)-conjugated goat antirabbit IgG antibody (Cat. No. SC-2054; 1:5000 dilution; Santa Cruz). Signals were detected using the ECL-PLUS/Kit (Cat. No. RPN2132; Amersham) following the manufacturer's protocol. GAPDH was served as the internal standard.
Cell proliferation assay
To determine whether TSPAN8 plays an important role in lung cancer cell viability, MTT assay was performed in H1299 cells after TSPAN8 knockdown of lentivirus infection or 95C cells after transfection TSPAN8 overexpression plasmid. Briefly, the mid-log-phase growth cells were seeded into 96-well plates at a density of 2500 cells per well in triplicates, and a total of 10 μL CCK-8 solution was added to each well following incubation for 1, 2, 3, 4, and 5 days, respectively, followed by another 2-hour incubation. Optical density values at wavelength of 450 nm (OD450) were measured using the Epoch microplate spectrophotometer (Biotek).
Colony formation assay
To determine whether TSPAN8 plays an important role in lung cancer cell colony formation ability, 95-C/pcDNA3.0, 95-C/TSPAN8 and H1299/NC, and H1299/shTSPAN8 cells (800 cells/well) were reseeded in six-well plates and cultured at 37°C with 5% CO2 for 9 days to form normal colonies. The media were replaced every 3 days. Then, the colonies were washed twice with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde for 30 minutes at room temperature. The fixed cells were washed twice with PBS and stained for 10 minutes by 1% crystal violet (Cat. No. C0121; Beyotime). Then, they were washed with ddH2O and dried in air. The total number of colonies, which contained more than 50 cells, was counted under light microscopy. Image analysis was conducted using Image-Pro Plus 6.0 software (Media Cybernetics).
Flow cytometry
Cell cycle distribution and cell apoptosis were analyzed using flow cytometry. Briefly, H1299/NC and H1299/shTSPAN8 cells were trypsinized, collected, washed, and stained according to the manual. For cycle distribution analysis, cells were fixed in 70% alcohol at 4°C for 2 hours and stained with 50 μg/mL propidium iodide (PI) (Sigma-Aldrich) for 30 minutes at room temperature in the dark. For cell apoptosis, cells were stained with Annexin V-fluorescein isothiocyanate (FITC; BD Pharmingen) and PI for 10 minutes at room temperature in the dark according to the manufacturer's protocol (BD Pharmingen).
Statistical analysis
Data analysis was performed using GraphPad Prism 5.01 software. All assays were repeated three times, and the results are expressed as the mean ± standard deviation. The statistical significance of results between each group was determined using one-way analysis of variance. Differences were considered significant at *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
TSPAN8 was abnormally upregulated in NSCLC cell lines and TSPAN8 expression was altered by knockdown and overexpression
To determine the expression levels of TSPAN8 in NSCLC cell lines, TSPAN8 mRNA level and protein expression in human lung adenocarcinoma and normal HBE cells were determined by RT-PCR and western blot. The authors found that low expression of TSPAN8 was detected only in HBE cells, while strong TSPAN8 expression was observed in NSCLC cell lines (Fig. 1A–C). To explore the role of TSPAN8 in NSCLC, knockdown of TSPAN8 was performed in H1299 cells (relatively highest expression of TSPAN8) and overexpression was executed in 95C cells (relatively lowest expression of TSPAN8), respectively. Subsequently, TSPAN8 protein expression was determined by western blot. The data suggest that shTSPAN8 specifically reduced TSPAN8 expression in H1299 cells and pcDNA3.1 efficiently enhanced TSPAN8 expression in 95C cells (Fig. 1D). It also reflected that the silence and overexpression methods were successful.
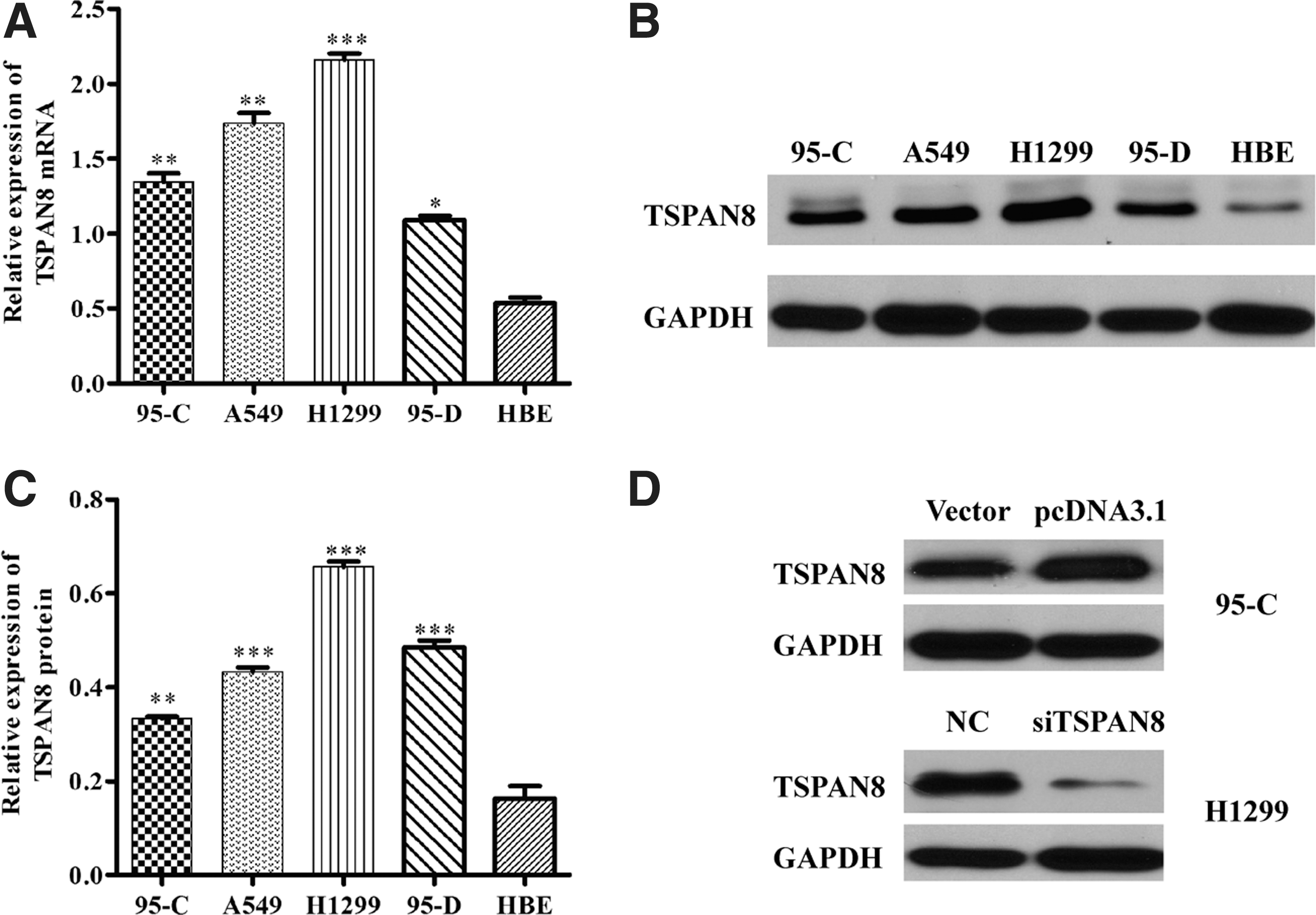
TSPAN8 was abnormally upregulated in NSCLC cell lines and TSPAN8 expression was changed by silencing and overexpression. Compared with normal HBE cell, high expression of TSPAN8 is found in nonsmall lung cancer cell lines (95C, A549, H1299, and 95D).
TSPAN8 affected cell proliferation and colony formation of NSCLC
To further assess the role of TSPAN8 in regulating NSCLC cell viability and proliferation, CCK-8 assays were performed on 95C/pcDNA3.0, 95-C/TSPAN8, H1299/NC, and H1299/shTSPAN8 cells. Figure 2A shows that the viability of 95C/TSPAN8 cells was markedly increased by overexpression of TSPAN8 (p < 0.001 compared with control). Meanwhile, the cell viability of H1299 cells was markedly inhibited by TSPAN8 knockdown (Fig. 2B, p < 0.001 compared with control).
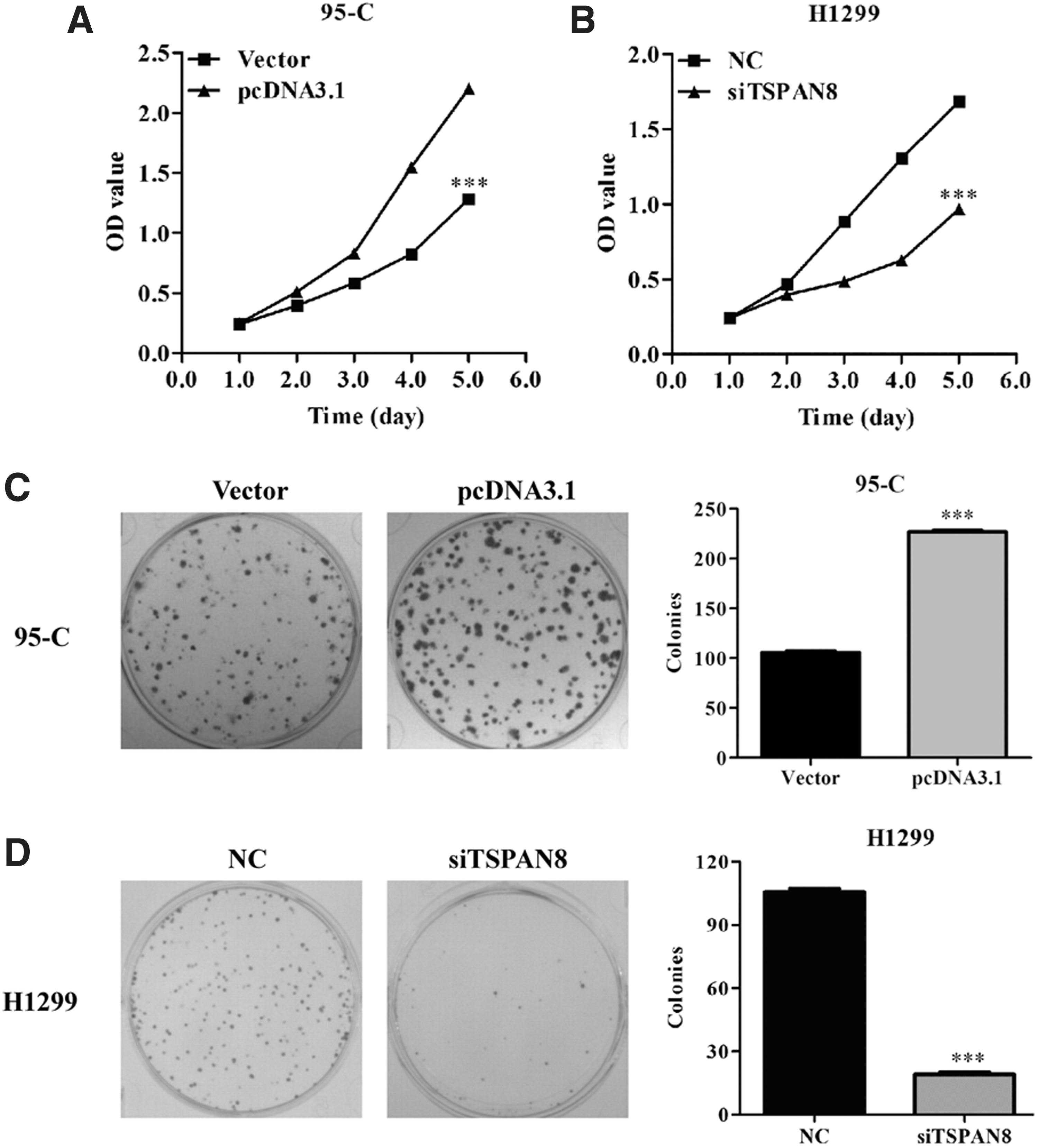
TSPAN8 expression affected the viability and proliferation of 95C and H1299 cells.
Moreover, the effect of TSPAN8 on colony formation ability was determined on two lung cancer cell lines. For 95C cells, representative images of colonies from different treatments (control vector and pcDNA3.1) indicated that overexpression of TSPAN8 significantly enhanced the number and size of colonies (Fig. 2C, p < 0.001). On the contrary, the ability of colony formation was remarkably impaired by shTSPAN8 in H1299 (Fig. 2D, p < 0.001). These results showed that TSPAN8 might exhibit a significant effect in NSCLC cell viability and proliferation.
Knockdown of TSPAN8 induced cell cycle progression and induced apoptosis in NSCLC
To further investigate the underlying mechanisms of modified cell proliferation and colony formation induced by TSPAN8, flow cytometry with PI staining for cell cycle distribution profiles and AnnexinV-FITC/7-ADD for cell apoptosis were performed in H1299 infected with shTSPAN8. As shown in Figure 3A, obvious differences in the cell cycle distribution profiles between H1299/NC and H1299/shTSPAN8 are displayed. Further analysis (Fig. 3B) showed that knockdown of TSPAN8 significantly increased the percentage of cells in G0/G1 phase from 51.74% to 65.62% (p < 0.001), while it decreased the percentage of cells in G2/M phase from 9.15% to 7.14% (p < 0.001). In addition, flow cytometry analysis showed that the percentage of both early apoptotic (Annexin V+/7-AAD−) and late apoptotic cells (Annexin V+/7-AAD+) was higher in H1299 cells infected with shTSPAN8 (Fig. 3C). Concretely, the apoptosis rate (early and late apoptotic cells) of H1299 was about 18.02% for H1299/shTSPAN8, which is markedly higher compared with the H1299/NC group, 9.88% (p < 0.001).
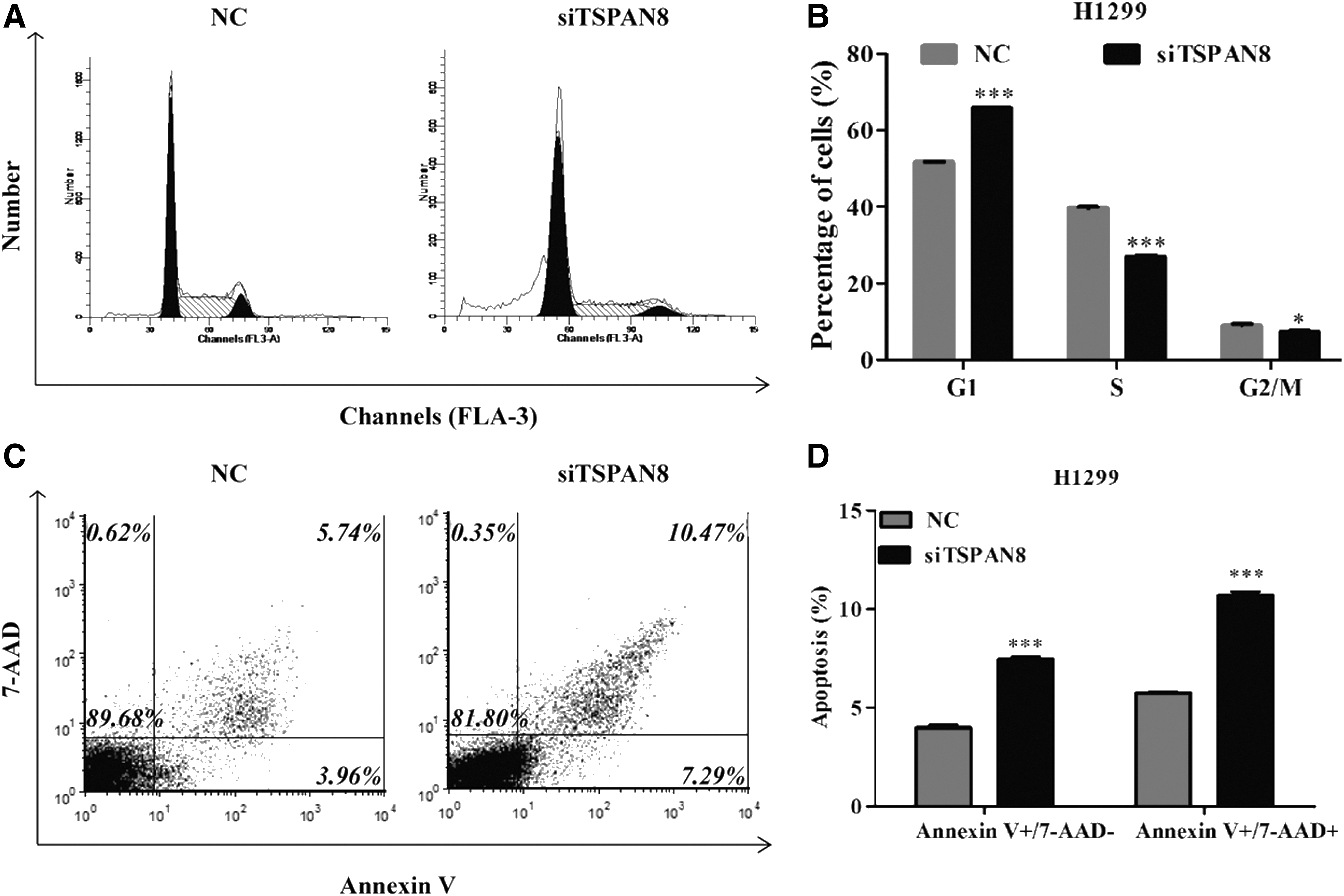
shTSPAN8 modulated cell cycle progression and induced apoptosis in H1299 cells.
Knockdown of TSPAN8 modified the expression of cell cycle and apoptosis-related proteins
To further illuminate above findings, the authors detected the expression alterations of several cell cycle and apoptosis-related regulators with western blot analysis. As shown in Figure 4, silence of TSPAN8 upregulated proapoptotic protein (the cleavage of PARP and Bax), but downregulated cell cycle checkpoint kinase (CDK2 and CDK4) and Cyclin D1 in H1299 cells. These results clearly indicated that TSPAN8 silencing inhibited the proliferation of H1299 cells by inducing G2/M phase cell cycle arrest and apoptosis.

shTSPAN8 effects on cell apoptosis and cycle regulatory protein level. The levels of CDK2, CDK4, and Cyclin D1 protein in H1299 cells were analyzed by western blot of control vector and shTSPAN8. The levels of BAX and PARP protein in H1299 cells were detected by western blot of control vector and shTSPAN8. GAPDH was used as an internal loading control. Data are reported as mean ± SD of three independent experiments. **p < 0.01, compared with H1299/control vector.
Discussion
NSCLC is the most common type of lung cancer accounting for ∼90% of all cases, in which over 70% of cases are at locoregionally advanced stages or have distant metastasis at the time of presentation with subsequent poor prognosis. Thus, a number of studies were performed to overcome the challenge and develop novel therapeutic regimens. In this study, the authors discovered that TSPAN8 was overexpressed in several lung cancer cell lines, and then they carried out some work surrounding it.
TSPAN8 belongs to the TSPAN family and encodes TSPAN8 protein. TSPAN family proteins are involved in adhesion, motility, cell fusion, and matrix modulation, which have been studied to promote cell growth and migration by cooperation with integrins and nonintegrin partners. 19 It has been reported that TSPAN8 is abnormally expressed in different degrees among various tumors, including human colon carcinoma, colorectal tumor, pancreatic adenocarcinoma, prostate cancer, and gastric cancer. 20 –23 TSPAN8 plays a role with several partners and pathways, for example, CD151, ADAM12, S100B, OLFAM, E-Cadherin/p120-Catenin, AR, CD44V6, and α6β4 molecules, and also MEK and MAPK signals. 24,25 The influence of TSPAN8 has been studied in so many kinds of tumors and very few report on lung cancer, especially NSCLC. In combination with this research, it is worth exploring its role in NSCLC.
With regard to lung cancer, high expression of TSPAN8 has been found in nonsmall lung cancer cell lines (95C, A549, H1299, and 95D) compared with normal HBE cell. Among these, the expression in H1299 is the highest and, in 95C, it is relatively lowest. By using TSPAN8 overexpression and TSPAN8 shRNA vectors, the authors found that upregulation of TSPAN8 enhanced cell proliferation in 95C and downregulation of TSPAN8 inhibited cell proliferation in H1299, which demonstrates that TSPAN8 regulates NSCLC cell growth and colony formation capacity. TSPAN8 knockdown in H1299 cells led to a significant cell cycle arrest in G0/G1 phase. In addition, flow cytometry analysis also showed an increase of apoptosis in siTSPAN8-treated H1299 cells. Thus, these results strongly suggest that TSPAN8 may play a central role in NSCLC. Furthermore, silencing of TSPAN8 led to upregulation of proapoptotic protein (the cleavage of PARP and Bax), but downregulation of cell cycle checkpoint kinase (CDK2 and CDK4) and Cyclin D1 in H1299 cells (Fig. 4). These results clearly indicated that TSPAN8 silencing inhibited the proliferation of H1299 cells by inducing G1 phase cell cycle arrest and apoptosis. The authors further observed upregulation of Bcl-2-associated X protein (BAX) and PARP in TSPAN8 silencing H1299 cells. BAX functions as an apoptotic activator by binding to BCL2. The heterodimer can lead to the release of cytochrome c, which then triggers apoptosis and further promotes activation of Caspase 3, and thereby apoptosis. 26 PARP, whose prime activator is DNA damage and overexpression, has been proven to be relevant with the pathogenesis of a number of tumors. 27 So, there may be several links between TSPAN8 and PARP. In addition, TSPAN8 silencing may inhibit cell viability and proliferation through different pathways. CDK2/Cyclin complex acts in the G1-S transition to promote the E2F transcriptional program and the initiation of DNA synthesis and modulates G2 progression. 28 It is well known that Cyclin D-CDK4 complexes are major integrators of various mitogenic and antimitogenic signals. Cyclin D-CDK4 (DC) complexes regulate the cell cycle during G1/S transition by phosphorylating and inhibiting members of the retinoblastoma (RB) protein family. 29 The authors also find that CDK2, CDK4, and Cyclin D1 dramatically decreased in H1299 cells transfected with shTSPAN8. In combination with these, TSPAN8 silencing can induce NSCLC cell apoptosis and cell cycle arrest in the G0/G1 phase.
In conclusion, the observations show that the TSPAN8 gene may be considered as a novel candidate target for inhibiting NSCLC cell viability and proliferation, even though the deeply molecular mechanism of TSPAN8 in NSCLC cells should be explored in vivo. To some degree, the authors suggest that TSPAN8 could be a new biomarker in antilung cancer target.
Footnotes
Acknowledgment
Source of support: departmental sources.
Disclosure Statement
There are no existing financial conflicts.