
Abstract
Recent successes in the development of new therapies for metastatic melanoma, such as mitogen-activated protein kinase pathway inhibitors, anticytotoxic T lymphocyte-associated antigen-4, and programmed cell death protein 1/programmed cell death ligand 1 (PD-L1) pathway-blocking antibodies, as well as combination strategies, all yielded promising results, changing the continually evolving landscape of therapeutic options for patients with melanoma. One promising new treatment modality is based on the use of immunomodulatory monoclonal antibodies that enhance the function of components of the antitumor immune response such as T cells or block immunologic checkpoints that restrain effective antitumor immunity. Program death-1 receptor and its ligand, PD-L1, is a major mechanism by which a tumor suppresses T cell-mediated antitumor immune responses. Studies in mice have shown that GK-1, an 18 amino acid peptide from Taenia crassiceps cisticerci, has the potential to be used as a primary or adjuvant component for the treatment of cancers by stimulating proinflammatory cytokines. The authors hypothesized that treatment with GK-1 in combination with anti-PD-L1 will increase survival in mice bearing melanoma tumors. C57BL/6 mice were injected with B16-F10-luc2 cells and separated into four groups: control, GK-1, anti-PD-L1, and GK-1/anti-PD-L1. The tumor sizes were measured and monitored using calipers and bioluminescence. The GK-1 peptide in combination with anti-PD-L1 showed significantly longer survival (34 days) compared with the other groups (23–27 days). This means an increase; survival increased 47.82% in the mice treated with GK-1+anti-PD-L1, 21.7% in mice treated with GK-1 alone, and 6.08% in those mice treated with anti-PD-L1 only. Blood samples were collected at days 0, 14, and at euthanization or end of the experiment and monitored for cytokines using mouse-specific V-PLEX Proinflammatory Panel. A decrease in TNF-α, IL-4, IL-5, IL-6, and IL-10 serum levels was observed in the GK-1/anti-PD-L1 combination group that may explain the beneficial effects of the combination treatment in prolonging the life of mice bearing melanoma. The data indicate that GK-1/anti-PD-L1 combined therapy affectively increases survival and warrants further clinical investigations.
Introduction
Melanoma is the most malignant form of skin cancer, with its highest incidence in the Caucasian population. 1,2 New cases of melanoma in the United States would reached 73,870 in 2015 and that year, 9940 patients were expected to die from the disease. 3
Until recently, systemic therapy for metastatic melanoma was ineffective, the 5-year survival rate for patients with distant metastases was <10%, 4,5 but recent successes in the development of new therapies for metastatic melanoma, such as mitogen-activated protein kinase pathway inhibitors, anticytotoxic T lymphocyte-associated antigen-4 (CTLA-4), and programmed cell death protein 1 (PD-1)/programmed cell death ligand 1 (PD-L1) pathway-blocking antibodies, have all yielded promising results, changing the continually evolving landscape of therapeutic options for patients with the disease. 6 Melanoma immunotherapy is a promising new treatment modality based on the use of immunomodulatory monoclonal antibodies. These blocking strategies target the immune evasion of tumors by removing suppressive cell types through PD-1/PD-L1 neutralization, rescuing normal immune cell functionality, thus augmenting effectively the antitumor T cell response. 7
GK-1 is an 18-amino acid peptide derived from Taenia crassiceps cysticerci. 8 It has been shown to enhance CD86 and major histocompatibility complex (MHC) class II molecule expression on dendritic cells (DCs) and antigen-specific T cell activation. GK-1 stimulates the production of IFN-γ, TNF-α, and MCP-1. 9,10 Recently, the authors have demonstrated for the first time that subcutaneous GK-1-treated mice with experimental melanoma show a delayed tumor growth and had increased survival time by 42.58%. 11 Moreover, peripheral blood from GK-1-treated mice demonstrated a statistically significant decrease in serum IFN-γ, while lung metastatic analysis demonstrated a significant increase in IFN-γ and IL-12p70. The authors also observed a significant decrease in IL-17, IL-4, IL-22, IL-23, and IL-12p40 and a marginal decrease in IL-1b compared with controls. 12
These data suggest that GK-1 could exert antitumoral effects by promoting Th1 polarization 13,14 and decreasing regulatory T cells. 15 Considering the antimelanoma action of anti-PD-L1, the authors hypothesize that combined GK-1/anti-PD-L1 therapies could synergize and maximize antitumoral effects and prolong survival.
Materials and Methods
Animals
Six- to 8-week-old C57BL/6 male mice were purchased from Charles River Laboratories (Willmington, MA). All animal procedures were performed in accordance with the guidelines of the Institutional Animal Care and Use Committee at Texas Tech University Health Sciences Center (Lubbock, TX).
Cells
The B16-F10-luc2 Bioware cells were purchased from Perkin-Elmer Corp. (Norwalk, CT). B16-F10-luc2 cells were confirmed to be pathogen free by the IMPACT Profile I (polymerase chain reaction) at the University of Missouri Research Animal Diagnostic and Investigative Laboratory. Cells were grown in RPMI medium (Gibco, Grand Island, NY) supplemented with 10% heat-inactivated fetal bovine serum (PAA, Pasching, Austria) at 37°C in a humid 5% CO2 atmosphere. 11,16
Experimental procedures
Mice (n = 40) were injected subcutaneously in the flank with 2 × 105 B16-F10-luc2 murine melanoma cells. Two weeks after injection, the tumor size was assessed with caliper measurements of the length of the short and long tumor diameter. The tumor size was calculated as the tumor volume using the formula: V = (A 2 • B)/2. 17 Only mice that developed a tumor of 20 mm3 were included in the study. The day that the tumor reached 20 mm3 in size was defined as day 0 and treatments were started that day with GK-1 (an 18-amino acid synthetic peptide [GYYYPSDPNT-FYAPPYSA]) (AnaSpec, Inc., Fremont, CA), anti-PD-L1 (10F.9G2; BioXCell, West Lebanon, NH) or both. Mice were euthanized when moribund or lethargic or/and failing to respond to gentle stimuli, and the date of death was recorded. The health and care of all animals used in this project complied with the Principles of Laboratory Animal Care and the Guide for the Care and Use of Laboratory Animals (NIH publication No. 80-23, revised 1985) and the University Laboratory Animal Care Committee at Texas Tech University Health Sciences Center.
Tumor-bearing mice were separated into four different groups: GK-1 (n = 9), anti-PD-L1 (n = 8), GK-1/anti-PD-L1 (n = 10), control peritumoral (n = 5), and control intraperitoneal (IP; n = 5). The GK-1 treatment group received peritumoral injections of GK-1 (10 μg/100 μL of sterile saline solution) every 5 days as described earlier. 11 The anti-PD-L1 treatment group received IP injections of anti-PD-L1 antibody (200 μg in 100 μL of phosphate-buffered saline [PBS]) every 3 days as described by Cooper et al. 18 The combined immunotherapy group received peritumoral injections of GK-1 every 5 days and of anti-PD-L1 (200 μg in 100 μL PBS) every 3 days. The control GK-1 group received injections of sterile saline solution (100 μL) and the control anti-PD-L1 group received IP injections of 100 μL PBS. All experimental procedures were done under conditions that preclude discomfort for the animals.
Blood collection and cytokine measurement
At days 0, 14, and at euthanization, blood samples (0.5 mL) were collected from the facial vein in Eppendorf Tube® without an anticoagulant. Blood samples were centrifuged (2000 rpm) for 10 minutes at 4°C using a microcentrifuge (Eppendorf North America, Hauppauge, NY). Serum was collected and stored at −80°C. TNF-α, IL-1b, IL-2, IL-4, IL-5, IL-6, IL-10, IL-12p70, KC/GRO, and IFN-γ were evaluated using a mouse-specific V-PLEX Proinflammatory Panel (MesoScale Discovery System, Rockville, MD) and in group serum samples using the electrochemiluminescence multiplex system Sector 2400 imager (MesoScale Discovery System).
In vivo bioluminescence imaging
In vivo imaging of melanoma tumor growth was performed with the Caliper IVIS Lumina XR in vivo animal imaging system (PerkinElmer, Waltham, MA). The images were obtained on day 0 and every 3 days until the end of the experiment. The mice were anesthetized using the Caliper IVIS isoflurane anesthesia system by inhalation of 1%–2% isoflurane (Baxter Healthcare Corp., Deerfield, IL), and the IP injection of 200 μL D-luciferin (150 mg/kg) was performed before image acquisition. The photons emitted from the mice (positioned prone) in vivo were acquired for 1 minute. Regions of interests from displayed images were drawn around the tumor and quantified as photons per second using the Living Image software (PerkinElmer).
Histopathological analysis
Two mice per group euthanized between days 25 and 30 were randomly selected for histopathology evaluation. Sections of melanomas, lungs, livers, spleens, and kidneys were fixed in buffered 10% formalin. Tissues were processed in the usual technique to be paraffin embedded and then tissue sections were stained with hematoxylin and eosin. Slides were examined for inflammatory changes in lung sections and for metastases in the liver, lung, and kidney sections; for size and lymphoid follicle counts in spleen sections; and for mitosis counts and necrosis in melanoma sections.
Statistical analyses
The authors used Kaplan–Meier survival curves, Cox regression analysis for survival evaluation, and log γ regression for tumor size evaluation. Gaussian regression and Student's t-test were used to evaluate cytokines. Two comparisons were made for spleen size, spleen lymphoid follicle count, tumor mitosis count, and tumor necrosis ratio. Control mice were compared with treated mice. Combined treatment mice were compared with mice treated with individual therapies. For each comparison group, means were calculated with 15,000 bootstrap replicates taken. The ratios of the actual means were calculated. The 2.5th and 97.5th percentiles of the ratios of the 15,000 bootstrap replicates served as 95% confidence intervals (CIs). Variance and standard error for each of the 15,000 bootstrap ratio replicates were calculated. Wald's chi-square test was used. Null hypotheses were rejected when p < 0.05.
Results
Experimental melanoma
Forty male C57BL/6 mice were injected subcutaneously in the flank with 2 × 105 B16-F10-luc2 murine melanoma cells. The authors found that 87.5% of the mice (35 of 40 animals) developed a 20 mm3 tumor according to the established criteria. The 20 mm3 of tumor volume as inclusion criterion was achieved for 15 mice (42.85%) by the first week and 20 mice (57.14%) by the second week. In 3 mice, in which melanoma growth was not observed, melanotic patches near the sites of the injections were observed, which eventually disappeared. The success rate for generating tumors is higher than 80%, similar to rates reported by other groups. 11,19 –21
In vivo bioluminescence imaging
Longitudinal bioluminescence imaging was evaluated after inoculation of B16-F10-luc2 cells (time = 0) and every 3 days thereafter until euthanization. Interestingly, only 7.5% of the mice did not develop a tumor despite exhibiting bioluminescence. No metastatic spread was noted during the study. Quantification of the bioluminescence signal over time demonstrated a progressive increase in tumor growth (Fig. 1). However, linearity disappeared after 22 days, suggesting either poor vascularization or oxygenation. In addition, no significant difference was observed among the groups despite the survival effect noted in the combination group. For purposes of data presentation, only controls are shown. Importantly, these findings indicate that the use of bioluminescence to monitor tumor growth should be avoided when there is either poor vascularization or poor circulation since luminescence requires oxygen and luciferase that is injected to visualize the tumors requires good circulation. Thus, only data obtained using calipers to measure tumor size are presented.
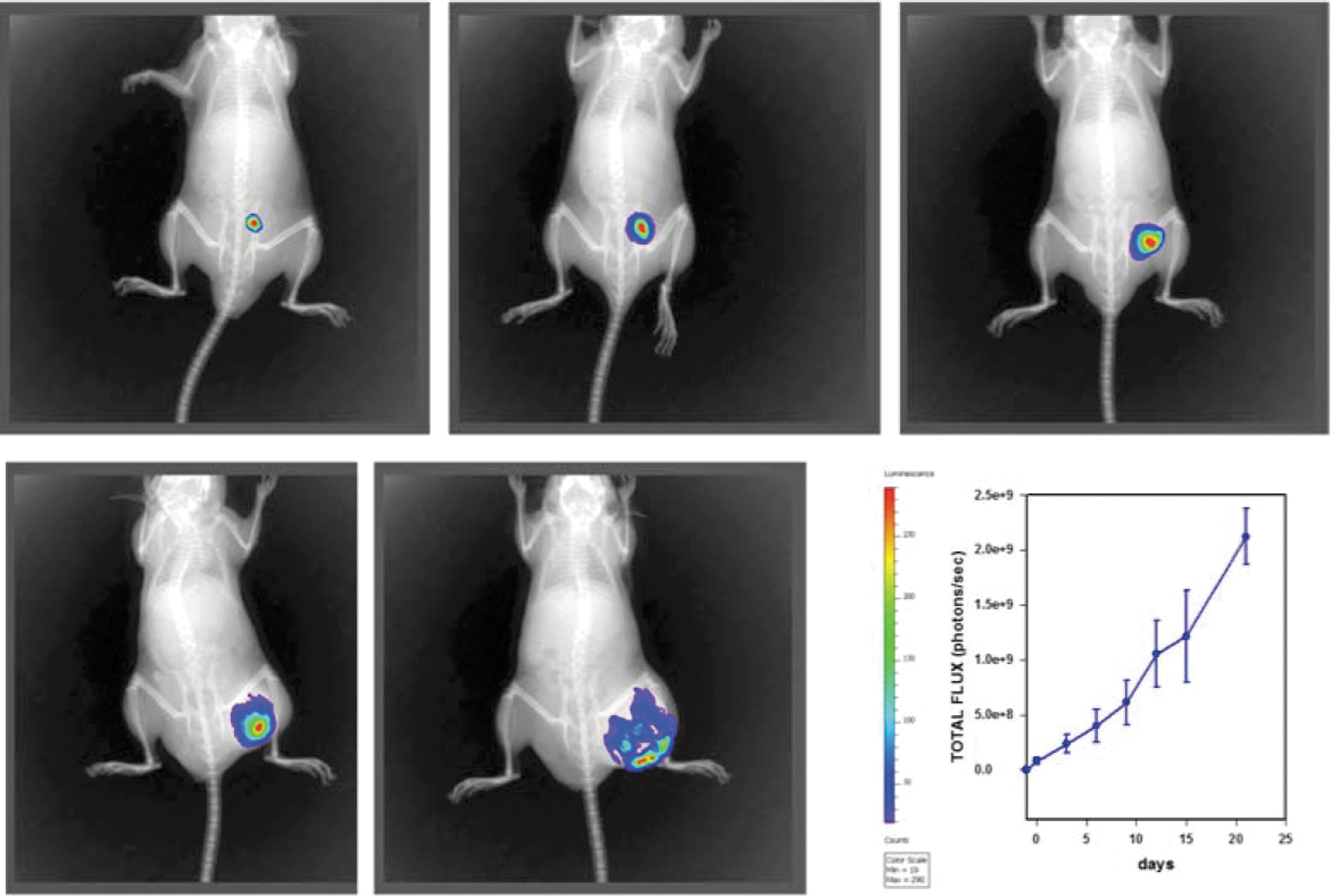
Six- to 8-week-old C57BL/6 mice were injected with melanoma cells (2 × 105 B16-F10-luc2 cells). Luminescent images of tumors were obtained with the caliper imaging system. The graph represents the mean and SEM of the total flux (photons/s) from six control mice. Similar images were obtained from mice treated with GK-1, anti-PD-L1, and the combination GK-1+anti-PD-L1 (not shown). PD-L1, programmed cell death ligand 1.
In vivo effect of GK-1, anti-PD-L1, and GK-1/anti-PD-L1 combination in experimental melanoma
From the 35 mice that developed a tumor, 5 mice died of causes unrelated to the tumor after the inoculum: 2 in the combination group, 1 in the anti-PD-L1 group, 0 in the GK-1 group, and 2 in the IP control group. Deaths were associated with blood sample collection and IP injections.
The GK-1/anti-PD-L1 group demonstrated a significantly longer survival rate compared with the other groups (p < 0.05). Interestingly, GK-1 treatment alone demonstrated a longer survival rate compared with the control group and the anti-PD-L1 treatment group (p < 0.05). Anti-PD-L1 treatment alone did not improve survival compared with the controls. The GK-1 peptide in combination with anti-PD-L1 showed significantly longer survival (34 days) compared with the other groups (23–27 days). This means an increase; survival increased 47.82% in the mice treated with GK-1+anti-PD-L1, 21.7% in mice treated with GK-1 alone, and 6.08% in those mice treated with anti-PD-L1 only. Tumor size was evaluated by log γ regression, taking into account the initial tumor day by means of an offset. Differences in the rates of tumor size were observed in the GK-1/anti-PD-L1 combination group compared with the other groups, mainly in two long-surviving mice, which had lower tumor volumes beginning at day 10. Statistical significance was observed at day 27 and continuing to the end of the study (p < 0.05). Rates of tumor size increase for the longest and second-longest surviving mice were 83% (95% CI 80–86) and 86% (95% CI 83–90), respectively (Figs. 2 and 3). These data indicate that mice treated with GK-1 in combination with anti-PD-L1, when affective, lived longer and had smaller tumor masses compared with controls.
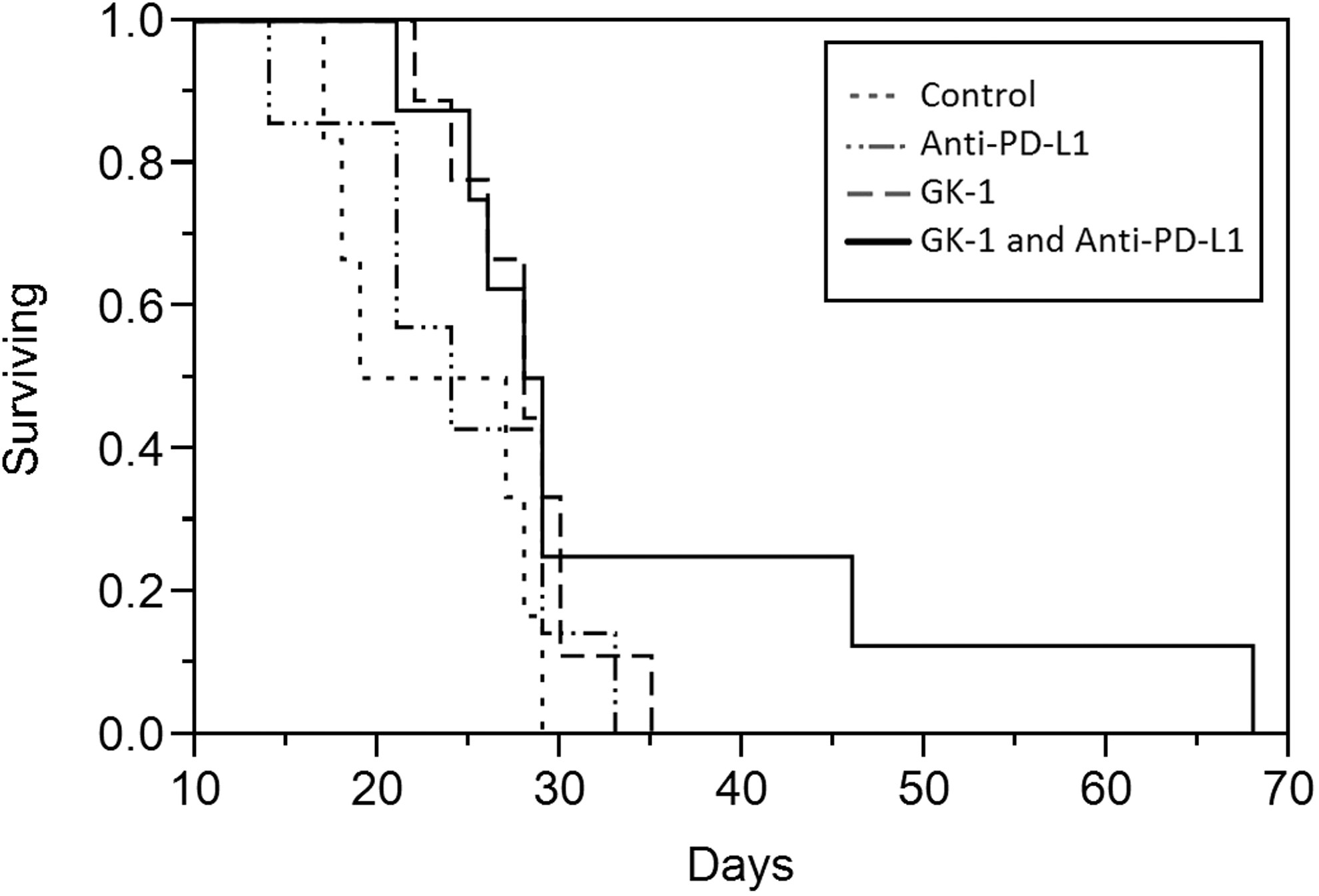
Therapeutic antitumor activity of the synthetic parasite-derived peptide GK-1 in combination with the anti-PD-L1 antibody against established melanoma. C57BL/6 mice were injected with 2 × 105 B16-F10-luc2. Tumor-bearing mice (20 mm3) were separated in four different treatment groups: GK-1 (n = 9), anti-PD-L1 (n = 7), GK-1/anti-PD-L1 (n = 8), and control (n = 6). Kaplan–Meier survival curves. Cox regression showed combined therapy mice had a lower hazard of death than did control mice (p < 0.05). Differences between GK-1 mice and control mice (p < 0.05) and between anti-PD-L1 mice and control mice (p > 0.05) were explicable by chance. Two long-surviving combined therapy mice accounted for the difference.
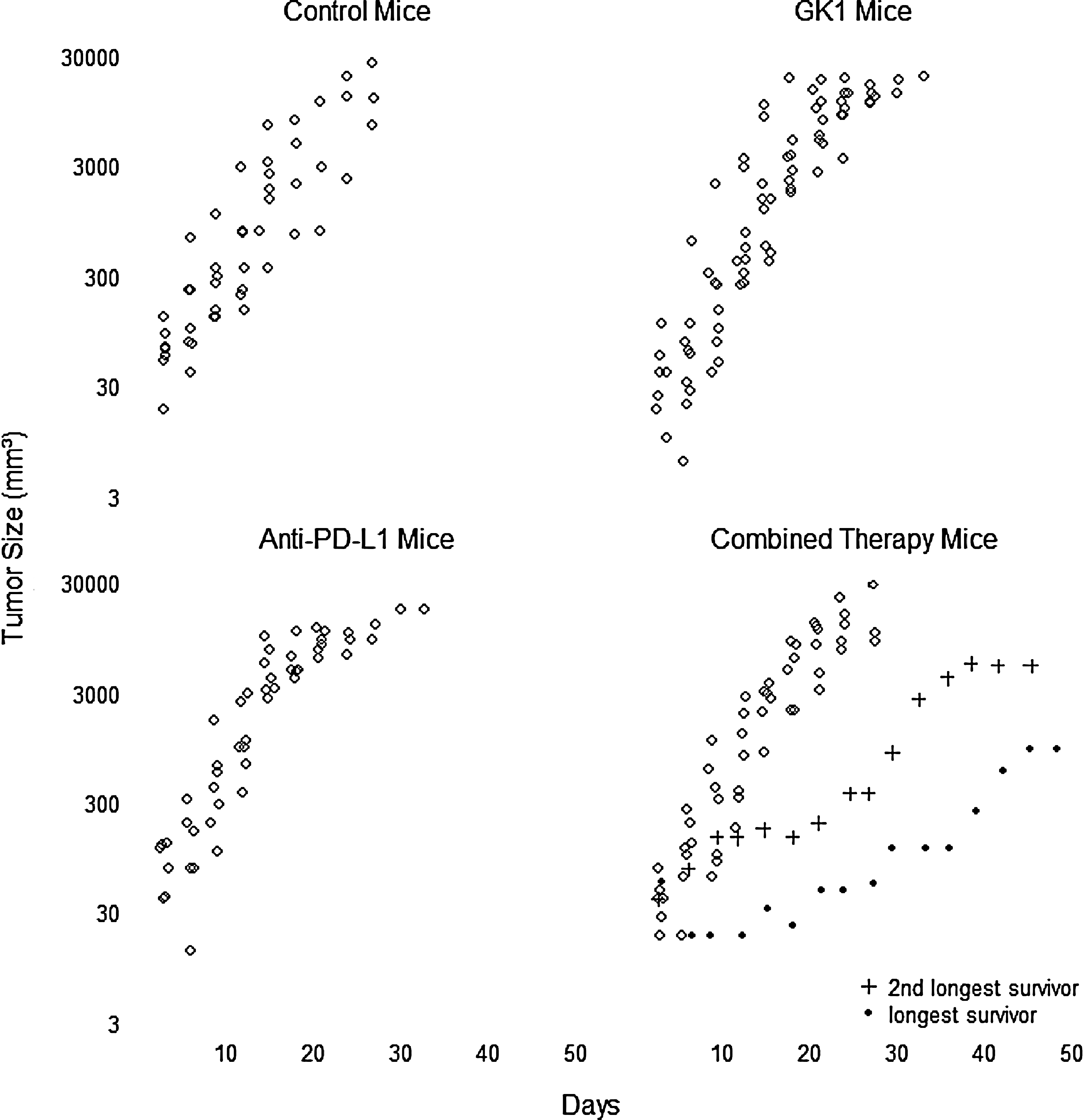
Murine melanoma tumor growth. Tumor volumes after treatment initiation (20 mm3) per group were measured daily with calipers; the two long-surviving combined therapy mice had lower tumor volumes at day 10, continuing to the end of the study. Log γ regression failed to show differences among groups in general. Tumor growth after 27 days of treatment was significantly decreased in the GK-1/anti-PD-L1 group compared with controls or treatments alone (p < 0.05).
Serum cytokine evaluation
To elucidate the serologic effects of GK-1, anti-PD-L1, and GK-1/anti-PD-L1 treatments, blood samples were collected to evaluate the cytokine profiles in the four groups at days 0, 14, and at euthanization. On day 14, 24 mice had levels of each cytokine; however, failure to detect cytokines was noted at a rate of IFN-γ, 13 (54%), IL-4, 8 (33%), and IL-5, 14 (61%). Two-tailed Fisher exact tests found only two comparison differences not explicable by chance: (i) in combined therapy, TNF-α levels were 0.008 (95% CI 0.002–0.014) pg/mL lower than in controls, and (ii) in anti-PD-L1 therapy, IL-2 levels were 0.006 (95% CI 0.001–0.011) pg/mL higher than in controls. At euthanization, the GK-1/anti-PD-L1 group showed significant decreases in IL-4, IL-5, IL-6, and IL-10 when compared with controls, whereas other cytokines (IL-1b, IL-2, IL-12p70, KC/GRO, IFN-γ, and TNF-α) were unaffected when compared with controls (Fig. 4).
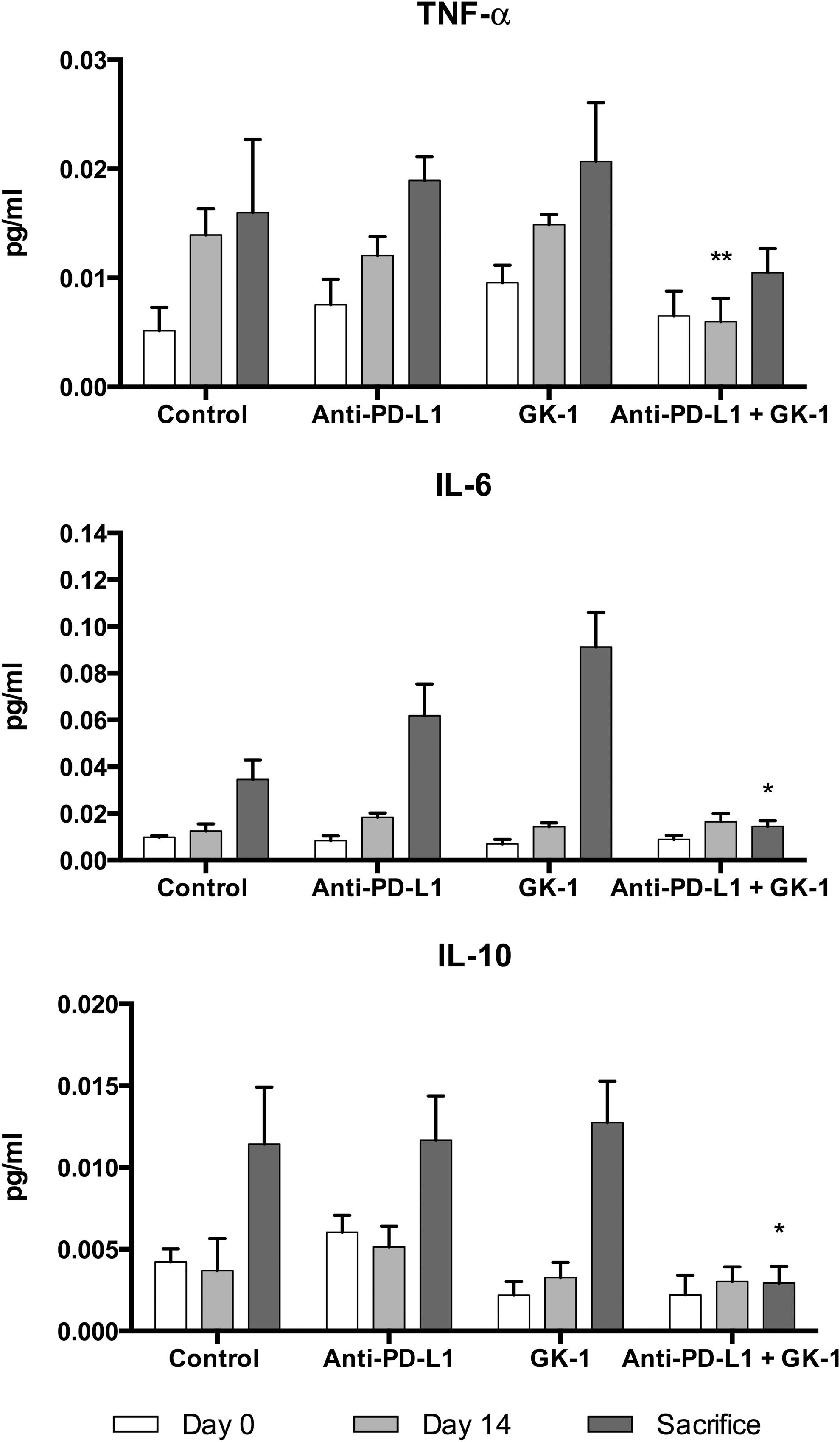
Peripheral blood cytokine evaluation. Blood samples were collected to evaluate the proinflammatory effects in the four groups at days 0, 14, and at sacrifice day. On day 14, TNF-α levels in the anti-PD-L1 and GK-1 therapy group were significantly lower compared with control mice. At sacrifice, the combined treatment group demonstrated significant decrease in cytokine production in IL-6 and IL-10. Data were analyzed using Student's t-test. *p < 0.05 when compared with control.
Histopathological analysis
Two mice from the GK-1, anti-PD-L1, and GK-1/anti-PD-L1 combination and control groups euthanized between days 25 and 30 were randomly selected for histopathological analysis. Lung sections from mice exposed to therapy, regardless of the compound, more often had pulmonary inflammation, although there were insufficient mice in this categorical assessment to make any definite statements among the groups (Table 1). For each mouse, total spleen lymphoid follicles, spleen size, total mitoses, and necrosis ratio were analyzed, no metastases were observed in either of the groups.
0, none; 1+, mild; 2+, moderate; 3+, severe; LymphMac, lymphocytes and macrophages; PD-L1, programmed cell death ligand 1; PMN, neutrophils.
Discussion
In the present work, the authors investigated the immunotherapeutic properties of combined GK-1 peptide and the antibody against the PD-L1, compared with GK-1, anti-PD-L1, or control in a subcutaneous melanoma mouse model. The results demonstrated that GK-1/anti-PD-L1-treated mice significantly increase the mean survival time (34 days) when compared with the control (23 days) mouse group. Mice treated with the GK-1/anti-PD-L1 combination had smaller tumor masses compared with the controls.
Several studies on advanced melanoma patients suggest that combining therapies targeting tumor mechanisms of immune evasion (e.g., removing suppressive cell types through PD-1/PD-L1 neutralization) with activation of normal immune cell functionality (i.e., T cell activation) may provide optimal benefits for patients. 22,23 The precise mechanism by which the GK-1/anti-PD-L1 combination treatment improved survival remains to be determined. However, the serum cytokine profile elicited by the combination treatment suggests a decrease of protumoral Th2 cytokines and stimulating a Th1 cytokine profile. Cytokines as immunomodulatory molecules have a potent antitumoral effect in mouse models of cancer. 24 Certain immunogenic human neoplasms, such as melanoma, are susceptible to modulation by immune cytokines. 25 There is a consensus that the induction of Th1 profiles or T cell release of cytokines such as IFN-γ and TNF-α is essential to the development of an effective antitumor immune response. 26,27
Recently, it has been demonstrated in a preclinical study that melanoma treatment with anti-PD-1 alone had no effect on tumor growth or survival, in part, due to the large size (1 cm diameter) of the tumors before treatment; however, anti-PD-L1 in combination with BRAFi led to significantly delayed tumor growth and improved survival relative to either therapy alone. 18 Similarly, in the study, anti-PD-L1 alone did not improve survival of mice with a 20 mm3 tumor size before treatment. However, the authors' group has shown the survival benefit of GK-1 treatment alone with the initial tumor volume of 20 mm3. 11 In the present work, GK-1 alone also improved survival when compared with the control and anti-PD-L1 alone.
Despite the favorable survival rate in GK-1/anti-PD-L1 combination-treated mice, some of which had smaller tumor masses, no significant differences were observed in tumor bioluminescence among mouse groups. Although bioluminescence is a convenient noninvasive tool for tumor assessment, many factors such as instrumentation, imaging parameters, animal hair, and depth of the tumor, can influence its sensitivity. Melanoma is a superficial tumor, but tends to grow rapidly with an increase of necrosis, hypoxia, and hypoperfusion, all of which have significant effects on bioluminescent signal intensity. A probable explanation for the bioluminescence results is the intense necrosis observed in the pathology assessment of melanoma in all groups of mice.
Previously, it was reported that GK-1 alone induces a significant decrease of serum IFN-γ in mice with experimental melanoma, whereas cytokine evaluation of lung with metastases demonstrated a significant increase of IFN-γ and IL-12p70 along with a significant decrease of IL-17, IL-4, IL22, IL23, and IL12p40, and a marginal decrease of IL-1β, compared with the untreated control group. 12 In contrast, PD-1 blockade shift antigen induced cellular reactivity toward a proinflammatory Th1/Th17 response, as evidenced by enhanced production of IFN-γ, IL-2, TNF-α, IL-6, and IL-17 and reduced production of the Th2 cytokines IL-5 and IL-13 in patients with prostate and advanced melanoma. 28
IFN-γ has been linked to PD-L1 expression in melanoma, 29 leading a possible adaptive immune resistance mechanism in which tumor cells under attack from IFN-γ-secreting T cells develop cell surface PD-L1 expression, thereby evading elimination. 29,30 Unfortunately, in the study, the authors failed to detect IFN-γ in the serum and no intratumor cytokine evaluation was performed. However, the decrease serum level of TNF-α at day 14 and IL-4, IL-5, IL-6 and IL-10 at euthanization in the GK1/anti-PD-L1 combination group is relevant and may partially explain the observed effects because some cytokines of Th2 response as IL-4 and IL-5, and anti-inflammatory or inflammation mediators as IL-10 and IL-6, respectively, have tumor-promoting activity. 31 The antimelanoma effect of the GK-1/anti-PD-L1 combination observed in the present study could be mediated by decreasing the protumor Th2 response. If this is the case, more antitumor effects could be induced as T cell proliferation, Th1 cytokine production, antigen presentation, and lymphokine-activated killer cell cytotoxicity. 32 Accordingly, IL-4 and IL-10 gene knockout mice with melanoma display significant suppression of tumor growth. Furthermore, in immunocompetent mice, neutralization of IL-4 decreased the metastatic ability of a highly metastatic melanoma cell line. 33 In humans, elevated levels of IL-4 and IL-10 in the peripheral blood lymphocytes, tumor-infiltrating lymphocytes, and/or serum have been associated with poor prognosis in squamous cell carcinoma, lymphoma, and pancreatic cancer, 34 –36 suggesting that these typical Th2 cytokines play a promoting role of tumor growth in vivo.
The significant decrease of IL-10, along with IL-4, IL-5, and IL-6, may be important for explaining the increased survival rate in the GK-1/anti-PD-L1 mouse group because melanoma cells increase IL-10 secretion as melanoma progresses, increasing prognostic implications of transition from the radial to the vertical growth phase and the developing risk of regional and visceral metastases. 32 Moreover, increased serum IL-10 has been shown to be a negative prognostic factor for responsiveness to treatment as well as to the overall survival in patients with melanoma and solid tumors, particularly lung, gastrointestinal, and renal cell cancer. 37
It has been shown that IL-6 promotes the development and progression of spontaneous melanoma, but the lack of IL-6 leads to a decrease in both melanoma incidence and size in transgenic mouse melanoma model. 38 In melanoma patients, Moretti et al. demonstrated a protective factor in patients with elevated IFN-γ, but progression was associated with IL-6, IL-10, and IL-12. 39 Additionally, patients with metastatic melanoma had higher serum IL-6 than healthy individuals and patients with elevated pretreatment serum IL-6 had a shorter overall survival rate compared with patients with normal serum. 40 The promoting tumor activity of IL-6 is related to apoptosis inhibition and tumor angiogenesis induction, favoring a larger tumor burden, resistance to chemotherapy and immunotherapy, and shorter median survival rate.
The authors also observed decreased serum IL-5 in the GK-1/anti-PD-L1 mouse group. In humans, elevated serum IL-5 has been detected in patients with advanced stage nonsmall cell lung cancer and their increased expression in breast carcinomas has been linked to higher rates of development of distant metastasis and poor prognosis.
The potential antimelanoma synergism effect activated by the GK-1/anti-PD-L1 combination can be partially explained by the mechanism of action of both molecules. GK-1 is a Th1 response inductor both in vitro and in vivo as it increases IFN-γ and IL-2, but not IL-4 and IL-10. 10,41,42 Moreover, GK-1 has high hydrophobic areas alternating with hydrophilic areas, which offer interesting possibilities for water and lipid interaction that may facilitate the peptide's interaction with B and T membrane-bound receptors. In this regard, GK-1 electronic polarity, adequately positioned anchor motifs, and similarities to motifs reactive with class I MHC molecules may explain the peptide's ability to induce a CD8+ proliferative response. 41,42
It is noteworthy that when PD-L1 signaling is reduced in T cells, these cells proliferate extensively in vitro and produce increased levels of IFN-γ and IL-17, suggesting an enhanced proinflammatory phenotype. Likewise, in the absence of PD-L1 silencing, as few as 104 activated DCs are sufficient to prime potent CD8+ T cell responses, which can control tumor growth. By improving the potency of DCs, in conjunction with the removal of PD-L1-based regulation, a significantly enhanced T cell immunity is capable of controlling tumor growth in a therapeutic setting. 43
The method of experimental tumor generation in animals, that is, spontaneous versus transplanted disease, is an area of controversy among immunotherapy researchers. 44 In this study, the authors elected to use a transplantable tumor model to facilitate the immunotherapeutic study, it is well know that spontaneous tumor models could potentially produce different immune-editing and host tolerance mechanisms. 45 In an elegant study performed by Lechner et al, tumor growth rate correlated indirectly with MHC class I expression and overall immunogenicity of the tumor model; B16 model showed a faster growth, meaning low immunogenic model and a marked absence of tumor-infiltrating effector cells. 45 These data are in agreement with the results, in which the control groups failed to show cell infiltration in the lung and tumor during histological evaluation; in contrast, GK-1 and anti-PD-L1 treatments showed increased cell infiltration.
Recently, it has been reported that intramelanoma administration of a novel TLR7/8 dual agonist, termed 3M-052, synergized with anti-CTLA-4 and anti-PD-L1, even when checkpoint blockade alone was ineffective. Treated tumors exhibited CCL2 chemokines and infiltration of M1 phenotype–shifted macrophages, which along the probably induced tumor-specific T cells, could be essential for the antitumor activity of 3M-052. 46 The results also demonstrated that anti-PD-L1 blockade alone was less effective than GK-1 treatment, but combination of anti-PD-L1+GK-1 effectively increases survival in an apparent mutual potentiation and whose mechanisms should be investigated.
Finally, the mouse model of melanoma used in the present study correlates with human Stage IIb melanoma (T4a, N0, M0, T4a >4.0 mm: w/o ulceration). According to the tumor-node-metastasis classification system of the American Joint Committee on Cancer treatment, this melanoma stage includes primary wide excision, followed by adjuvant clinical trials or observations. 11 Peritumor and systemic therapy combination was evaluated, which is relevant as a preliminary approach. When intralesional therapies are combined with systemic therapies as with T-VEC and ipilimumab, studies have shown augmented response rates in humans. 46 As stated recently by Agarwala, current Phase II/III trials are under development with newer intralesional agents, Allovectin-7 (velimogene aliplasmid), plasmid IL-12, talimogene laherparepvec (T-VEC), and PV-10, which have shown particular promise for their low toxicity, easy administration, and likely systemic immune effects. 46
Footnotes
Acknowledgments
The authors wish to acknowledge the contribution of Texas Tech University Health Sciences Center Clinical Research Institute and Dr. Diana Torres García for their assistance with this research. TTUHSC at the Permian Basin Regional Dean's Office provided financial support to J.V.-A.
Disclosure Statement
No competing financial interests exist.