
Abstract
Overexpression of extracellular matrix metalloproteinase inducer (EMMPRIN) accelerates tumor invasion and metastasis via activation of matrix metalloproteinases (MMPs) and urokinase-type plasminogen activator (uPA) expression. The authors were interested in whether the scFv-M6-1B9 intrabody against EMMPRIN that retains EMMPRIN in endoplasmic reticulum could be a potential tool to suppress cervical cancer invasion through inhibition of uPA. The chimeric adenoviral vector Ad5/F35-scFv-M6-1B9 was transferred into human cervical carcinoma HeLa cells to produce the scFv-M6-1B9 intrabody against EMMPRIN. Cell surface expression of EMMPRIN, the membrane-bound uPA, the enzymatic activity of secreted uPA, and the invasion ability were analyzed. The scFv-M6-1B9 intrabody successfully diminished the cell surface expression of EMMPRIN and the membrane-bound uPA on HeLa cells. uPA activity from tissue culture media of EMMPRIN-downregulated HeLa cells was decreased. The invasion ability of HeLa cells harboring scFv-M6-1B9 intrabody was also suppressed. These results suggested that the scFv-M6-1B9 intrabody might represent a potential approach for invasive cervical cancer treatment. The application of scFv-M6-1B9 intrabody in animal experiments and preclinical studies would be investigated further.
Introduction
Extracellular matrix metalloproteinase inducer (EMMPRIN) is a cell surface glycoprotein belonging to the immunoglobulin superfamily. Its expression is often elevated in many forms of cancer such as breast cancer, hepatoma, esophageal carcinoma, colorectal carcinoma, ovarian carcinoma, and cervical cancer. 1 –3 Overexpression of EMMPRIN has been shown to accelerate the growth and invasion of cancer cells 4,5 and possesses the ability to induce the synthesis of matrix metalloproteinases (MMPs) in cancer and stromal cells. 6 –8 EMMPRIN interacts with MMPs and other molecules such as urokinase-type plasminogen activator (uPA) in the cancer progression process. 9
Downregulation of EMMPRIN by small interfering RNA (siRNA) reduced invasion and metastasis in cancer cells such as pancreatic cancer, 10 nonsmall lung cancer, 11 melanoma, 12 gastric cancer, 13 endometrial cancer, 14 ovarian cancer, 15 and cervical cancer 16 via disruption of MMP-2 and MMP-9 expression. Therefore, EMMPRIN has been suggested to be a potential molecular target for developing the therapeutic strategies to inhibit tumor cell dissemination.
An intrabody is an antibody designed to express inside the cells. Intrabodies can target proteins to various subcellular compartments through the fusion with localization signal sequences. 17 –20 Intrabodies could be useful tools not only for modulating or neutralizing the function of target molecules inside the cell 21,22 but also for therapeutic approaches in cancer. 23 An intrabody against EMMPRIN was successfully generated in 293A cells by using the recombinant scFv antibody fragments of the murine monoclonal antibody, clone M6-1B9 (scFv-M6-1B9 intrabody) in this laboratory. 24 This scFv-M6-1B9 intrabody reacted specifically to EMMPRIN and interrupted the transportation of EMMPRIN to the cell membrane.
The scFv-M6-1B9 intrabody was first introduced into cervical cancer HeLa cells, using adenovirus serotype 5 (Ad5)-based vectors and successfully reduced the cell surface expression of EMMPRIN. 25 Recently, the chimeric adenoviral vector Ad5/F35-harboring gene encoding the scFv against EMMPRIN (Ad5/F35-scFv-M6-1B9) was successfully constructed in this laboratory to enhance infectivity in coxsackievirus and adenovirus receptor (CAR)-negative cells. 26 The scFv-M6-1B9 intrabody produced by this adenoviral vector attenuated the cell surface expression of EMMPRIN, cell proliferation, invasion, and metastasis, and subsequently induced apoptosis in the colorectal adenocarcinoma cell line Caco-2. 26,27
As mentioned previously, the scFv-M6-1B9 intrabody blocked the expression of EMMPRIN on the cell surface of cervical cancer HeLa cells 25 and colorectal cancer Caco-2 cells. 26 However, disruption of metastatic enzymes MMP-2 and MMP-9 and their inhibitors tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2 was not observed in these studies. 25,26
The uPA system is a multifunctional system involved in cellular development and cancer progression, including angiogenesis, migration, and invasion. 28,29 This system significantly correlates to tumor aggressiveness and poor outcome. 29 uPA binds to its receptor (uPA receptor [uPAR]) on the cell surface and then triggers the conversion of plasminogen into plasmin. uPA is considered to be a critical trigger for plasmin generation during cell migration and invasion 28 and also plays an important role in tumor invasion and metastasis. 30,31
The uPA system has been shown to be upregulated by EMMPRIN. 9 The authors were thus interested in whether the scFv-M6-1B9 intrabody could inhibit cancer invasion through interruption of uPA expression and activity. In this study, human cervical carcinoma cell line HeLa was used as a representative model. Ad5/F35-scFv-M6-1B9 was used to transduce into the human cervical carcinoma cell line HeLa to produce the scFv-M6-1B9 intrabody. Cell surface expression of EMMPRIN and membrane-bound uPA expression, uPA activity, and the invasive behavior of HeLa cells were analyzed.
Materials and Methods
Cell lines
The embryonic human kidney cell line (293A) (Invitrogen, Carlsbad, CA) and the cervical carcinoma cell line (HeLa) (kindly gifted from Prof. Dr. Andre Lieber, University of Washington, Seattle, WA) were maintained in Dulbecco's modified Eagle's medium (DMEM) (Gibco, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere of 5% carbon dioxide (CO2) at 37°C.
Preparation of adenoviruses (Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, and Ad5/F35-GFP)
The adenovirus expressing scFv intrabody against EMMPRIN (Ad5/F35-scFv-M6-1B9) and adenovirus expressing scFv intrabody against the irrelevant protein (Ad5/F35-scFv-irrelevant) for intrabody control were previously constructed in this laboratory. 26 Adenovirus not expressing scFv intrabody (Ad5/F35-GFP) for transduction control was a generous gift from Dr. Saw-See Hong, University of Lyon 1, France. Adenoviruses were generated and propagated as described elsewhere. 24,32 Titers were determined by plaque titration in 293A cells. 32
Analysis of uPA by Western blot analysis
The HeLa cells were transduced at an multiplicity of infection (MOI) of 100 pfu/cell of Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, or Ad5/F35-GFP and harvested after 48 hours. Membrane and cytoplasmic proteins were extracted by the ProteoJET™ Membrane Protein Extraction Kit (Fermentas, St. Leon-Rot, Germany). Afterward, the proteins were electrophoresed on a 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel under reducing conditions and then electrotransferred onto a polyvinylidene-fluoride (PVDF) membrane. Membranes were blocked with 5% skim milk in phosphate-buffered saline and incubated with 1 μg/mL anti-uPA monoclonal antibody (Calbiochem, Darmstadt, Germany). After washing, peroxidase-conjugated goat anti-mouse immunoglobulin antibody (KPL, Gaithersburg, MD) was added onto the membranes. Finally, the immunoreactive bands were detected using an enhanced chemiluminescent (ECL) substrate detection system (Thermo Scientific, Schaumburg, IL).
Analysis of EMMPRIN on cell surface of HeLa by flow cytometry
Five hundred microliters of 1.6 × 105 cells/mL HeLa were transduced with Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, or Ad5/F35-GFP. After 48 hours, HeLa cells were washed with fluorescence-activated cell sorting (FACS) diluent and were blocked with human AB serum for 30 minutes on ice. Then, 25 μL of EMMPRIN culture supernatant antibody (M6-1B9) (kindly provided by Prof. Dr. Watchara Kasinrerk, Chiang Mai University, Thailand) was added in blocked cells for 30 minutes on ice. After washing, 25 μL PE-conjugated F(ab′)2 fragment of sheep anti-mouse immunoglobulin antibody was added and incubated for 30 minutes on ice. Finally, cells were washed 3 times and resuspended with 1% paraformaldehyde in FACS diluent. The fluorescence intensity of the stained cells was measured by flow cytometry.
uPA activity by casein–plasminogen zymography
The uPA activity was assessed from the ability of uPA to digest its substrates, casein and plasminogen. HeLa cells (4 × 105 cells/mL/well) were transduced by Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, or Ad5/F35-GFP. After 48 hours, cells were washed twice with DMEM without FBS. Cells were cultured in a serum-free medium for 48 hours. The supernatants were collected and centrifuged at 11,600 g for 10 minutes at 4°C. The serum-free conditioned medium was electrophoresed on SDS-PAGE containing 2 mg/mL casein and 10 μg/mL plasminogen under nonreducing conditions. After electrophoresis, gels were washed twice with 2.5% Triton X-100 for 30 minutes at 37°C and incubated in digestion buffer (50 mM Tris, 10 mM CaCl2, 150 mM NaCl, 0.02% NaN3, and pH 7.4) with gentle shaking at 37°C for 16 hours. Proteins on the gel were stained with 0.2% Coomassie Brilliant Blue. The zones of uPA activity were shown by negative staining (clear zone) and analyzed by Image Quant TL image analysis software.
Matrigel invasion assay
The invasion of cells was analyzed using a Falcon BioCoat Matrigel chamber (Becton Dickinson) with 8-μm pore size. HeLa cells (8 × 104 cells/500 μL/well) were plated into 24-well culture plates and incubated at 37°C, 5% CO2 for 24 hours. Cells were transduced by MOI of 100 pfu/cell of concentrated crude Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, or Ad5/F35-GFP and incubated at 37°C, 5% CO2. After 48 hours, cells were harvested and suspended to a final concentration of 5 × 104 cells/mL in serum-free media. Then, 500 μL of cells were plated into the upper side of a Matrigel chamber. The lower side of the chamber was given 750 μL of 10% FBS-DMEM. The cells were then incubated at 37°C, 5% CO2 for another 24 hours. The noninvading cells on the upper surface of the chamber were removed with a cotton swab, and cells on the lower surface were stained with Wright-Giemsa and counted under a brightfield microscope.
Statistical analysis
All data are expressed as mean ± standard deviation (SD). Comparisons between groups were carried out using an analysis of variance (one-way ANOVA). p-Value <0.05 was considered statistically significant.
Results
Cell surface expression of EMMPRIN in Ad5/F35-scFv-M6-1B9-transduced HeLa cells
To assess the effect of scFv-M6-1B9 intrabody on the cell surface expression of EMMPRIN, HeLa cells were transduced by Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, and Ad5/F35-GFP at 100 MOI and analyzed for the surface expression of EMMPRIN at 48 hours after transduction by flow cytometry. The expression of green fluorescent protein (GFP) in transduced HeLa cells is illustrated (Fig. 1A). HeLa cells that were transduced with Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, and Ad5/F35-GFP at 100 MOI showed similar GFP expression (Fig. 1A). This result indicates that all Ads had essentially identical infectivity for HeLa cells.
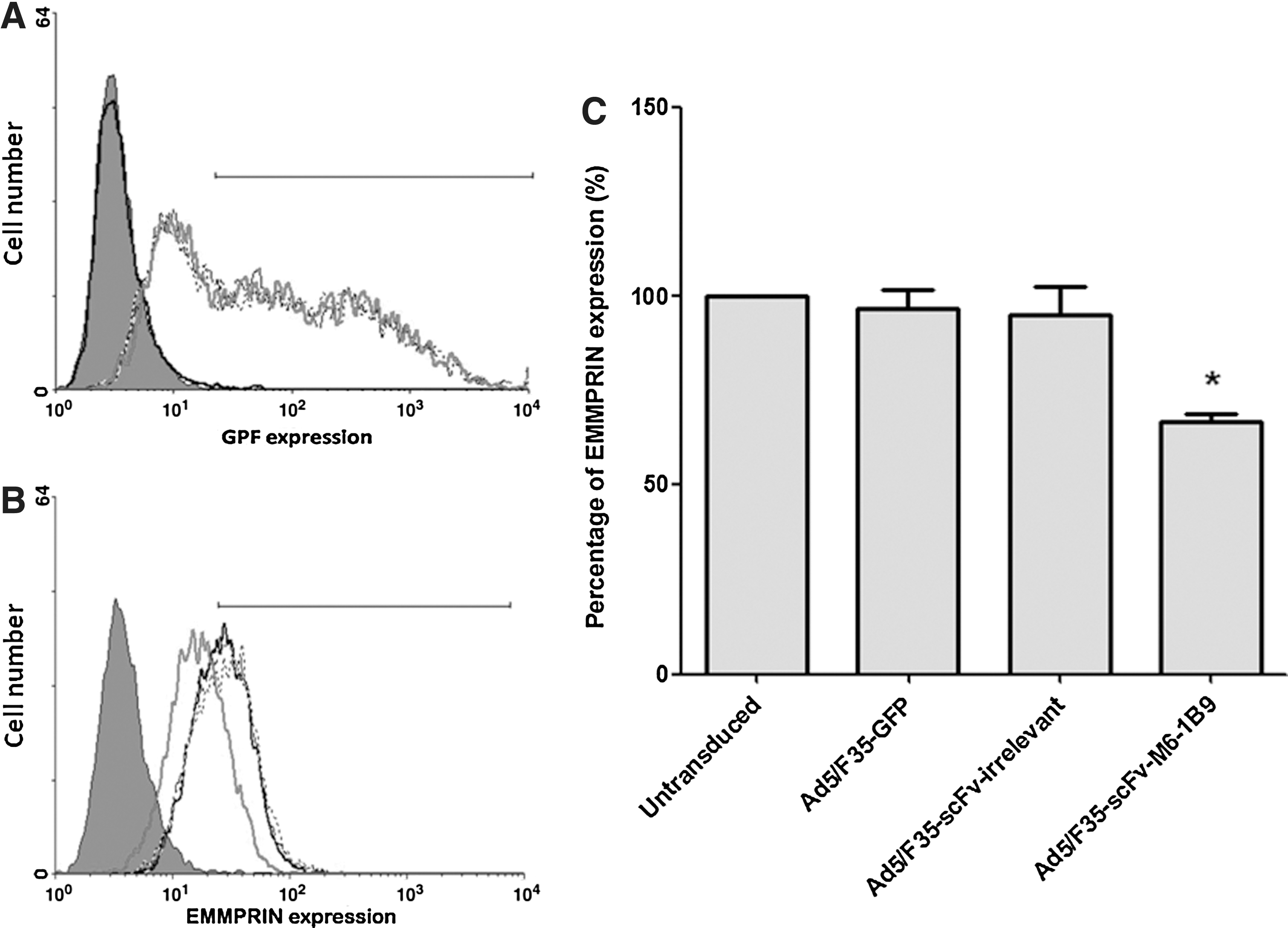
Downregulation of EMMPRIN on HeLa cells by scFv-M6-1B9 intrabody. HeLa cells were transduced by Ad5/F35-scFv-M6-1B9, Ad5/F35-scFv-irrelevant, and Ad5/F35-GFP at 100 MOI.
EMMPRIN cell surface expression in transduced HeLa cells was examined at 48 hours after transduction. Cell surface expression of EMMPRIN was significantly reduced in Ad5/F35-scFv-M6-1B9 transduced HeLa cells compared to Ad5/F35-scFv-irrelevant transduced HeLa cells, Ad5/F35-GFP transduced HeLa cells, and untransduced HeLa cells (Fig. 1B; p < 0.05). The fluorescence intensity of conjugate control HeLa cells is shown in solid shade. Data represent the average percentage of EMMPRIN expression ± SD of three independent experiments, as shown in Figure 1C. This result indicates that scFv-M6-1B9 intrabody generated by Ad5/F35-scFv-M6-1B9 could inhibit EMMPRIN expression on the surface of HeLa cells.
Membrane-bound uPA in EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody
To determine the membrane-bound uPA in EMMPRIN-downregulated HeLa cells, the membrane and cytoplasmic fractions of transduced HeLa cells were electrophoresed on 12% SDS-PAGE and electrotransferred onto a PVDF membrane. The presence of uPA in the membrane and cytoplasmic fractions was detected by Western blot analysis. β-actin and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were used as loading controls for membrane and cytoplasmic fractions, respectively. The membrane-bound uPA was decreased on Ad5/F35-scFv-M6-1B9 transduced HeLa cells compared to Ad5/F35-scFv-irrelevant transduced HeLa cells, Ad5/F35-GFP transduced HeLa cells, and untransduced HeLa cells (Fig. 2A).
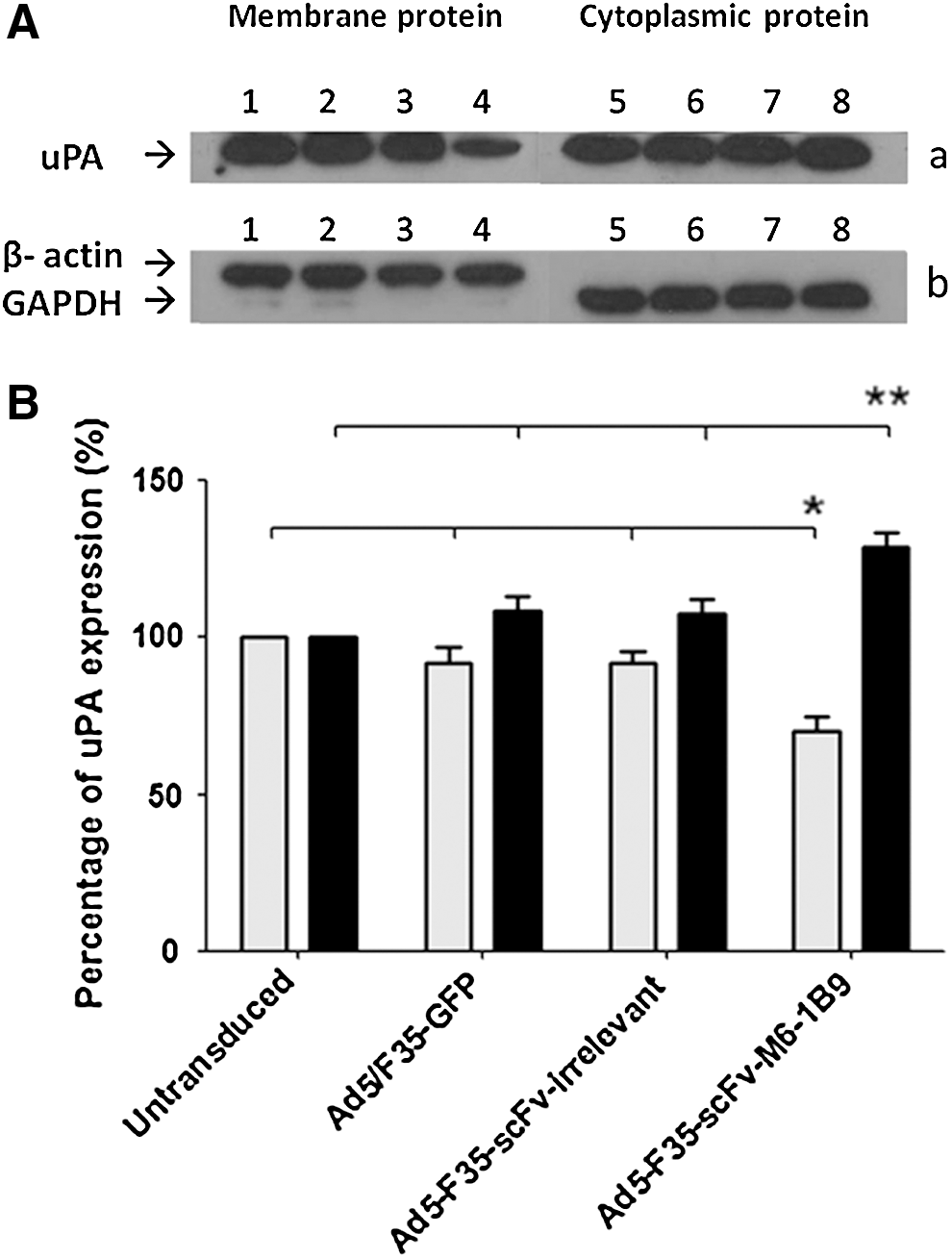
uPA expression in protein extracts from EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody.
As shown in Figure 2B, the membrane-bound uPA from Ad5/F35-scFv-M6-1B9 transduced HeLa cells was significantly decreased (70%) compared to Ad5/F35-scFv-irrelevant transduced HeLa cells (92%), Ad5/F35-GFP transduced HeLa cells (91%), and untransduced HeLa cells (100%) (p < 0.05). In addition, cytoplasmic uPA in Ad5/F35-scFv-M6-1B9 transduced HeLa cells was significantly increased (129%) compared to Ad5/F35-scFv-irrelevant transduced HeLa cells (107%), Ad5/F35-GFP transduced HeLa cells (108%), and untransduced HeLa cells (100%) (Fig. 2A, B; p < 0.05). These results indicate that scFv-M6-1B9 intrabody reduced the membrane-bound uPA and resulted in accumulation of the uPA inside the HeLa cells.
uPA activity in culture supernatant from EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody
Casein–plasminogen zymography was performed to demonstrate that the scFv-M6-1B9 intrabody was involved in the regulation of uPA activity in HeLa cells. One role of uPA is to convert a zymogen plasminogen into active plasmin. Active plasmin in turn can cleave extracellular matrix proteins such as casein. As shown in Figure 3, the uPA activity in culture supernatant from Ad5/F35-scFv-M6-1B9 transduced HeLa cells was significantly decreased compared to that from Ad5/F35-scFv-irrelevant transduced HeLa cells, Ad5/F35-GFP transduced HeLa cells, and untransduced HeLa cells (100%) (Fig. 3A; p < 0.05). This result indicates that uPA activity in tissue culture media of EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody was significantly attenuated.
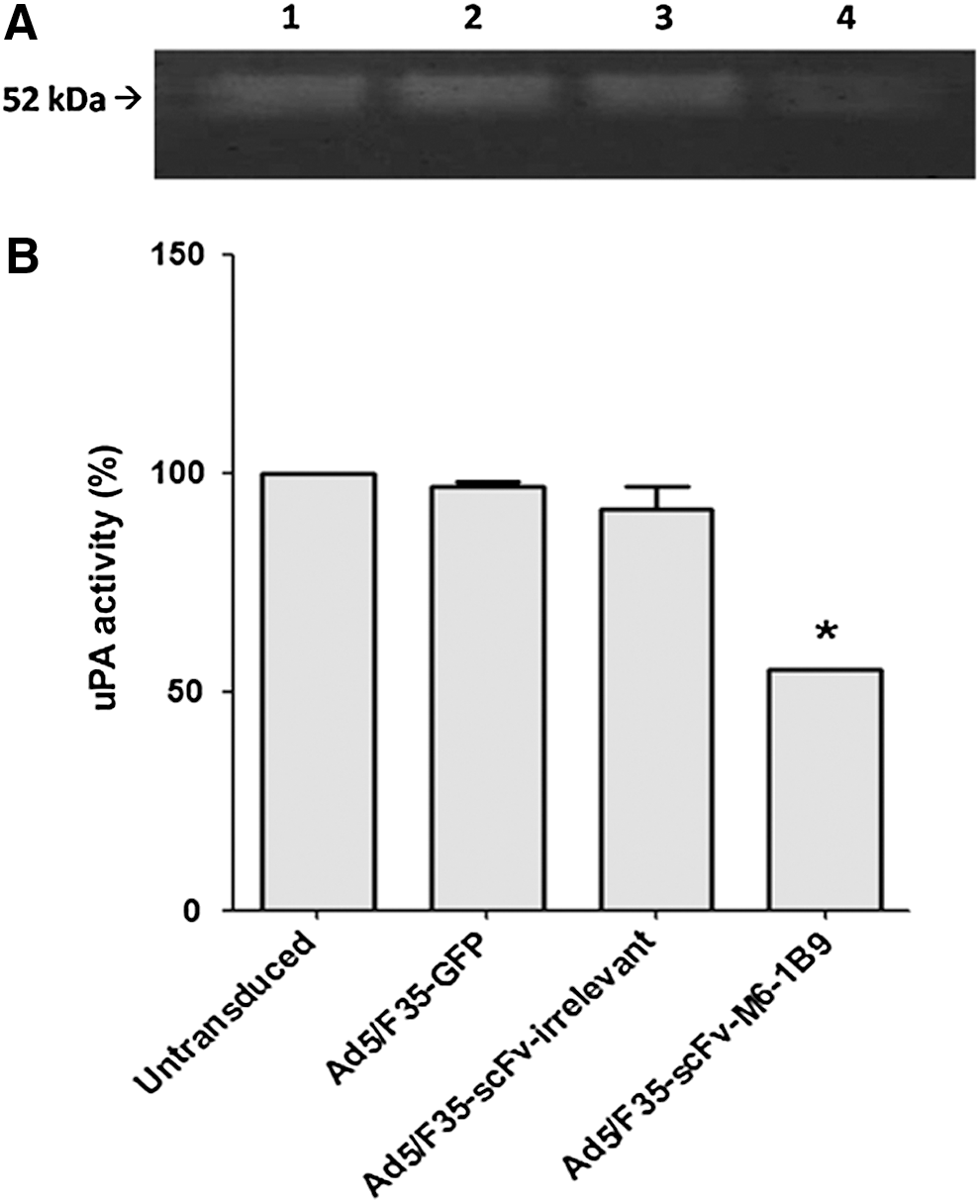
uPA activity in culture supernatant from EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody.
Invasion ability of EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody
To determine the invasion ability of EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody, HeLa cells were transduced by Ads and replated into the upper side of a Matrigel chamber. The number of invading cells on the lower side of the chamber indicated that invasion ability of Ad5/F35-scFv-M6-1B9 transduced HeLa cells was also significantly decreased (38%) compared to Ad5/F35-scFv-irrelevant transduced HeLa cells (94%), Ad5/F35-GFP transduced HeLa cells (100%), and untransduced HeLa cells (100%) (Fig. 4; p < 0.05). This shows that invasion ability of HeLa cells carrying the scFv-M6-1B9 intrabody was decreased when compared to controls.

Invasion ability of EMMPRIN-downregulated HeLa cells expressing scFv-M6-1B9 intrabody. HeLa cells were transduced by Ads and replated into the upper side of a Matrigel chamber. The number of invading cells on the lower side of the chamber was counted to indicate invasion ability.
Discussion
EMMPRIN is widely distributed on the cell surface and involves in a variety of cellular mechanisms in various cell types, such as leukocytes, endothelial cells, fibroblasts, and cancer cells. 33 Its expression is often elevated on the surface of malignant tumor cells as in breast cancer, hepatoma, esophageal, cervical, colorectal, and ovarian squamous cell carcinomas. 34 Previous studies have suggested that overexpression of EMMPRIN correlates with the progression of cervical cancer and serves as a marker for the evaluation of the recurrence probability in cervical cancer patients. 5 Several studies have shown that decreased expression of EMMPRIN can reduce the progression of cancer cells.
It has been previously reported that downregulation of EMMPRIN can be accomplished through RNAi technology. 13,35 Downregulation of EMMPRIN by transfection with siRNA resulted in decreased invasion activity in a cervical squamous carcinoma cell line, 36 prostate cancer cells, 35 and laryngeal carcinoma Hep2 cells. 37 EMMPRIN siRNA transfection of the human breast epithelial cells downregulated EMMPRIN expression, and a clear decrease in secreted uPA activity was also observed. 9
The authors were interested in whether the scFv-M6-1B9 intrabody against EMMPRIN could be used as an alternative tool to downregulate EMMPRIN expression and suppress cancer invasion. Recently, they have shown that the scFv-M6-1B9 intrabody generated by recombinant antibody fragments, scFv, of the murine monoclonal antibody, clone M6-1B9, using Ad5 gene transfer reacted specifically to EMMPRIN. 24 The transport of EMMPRIN to the cell membrane of 293A cells was successfully restricted. 24 The scFv-M6-1B9 intrabody was first introduced into cervical cancer HeLa cells, using Ad5-based vectors, and successfully reduced the cell surface expression of EMMPRIN. 25
The chimeric adenoviral vector Ad5/F35 harboring the gene encoding the scFv against EMMPRIN (Ad5/F35-scFv-M6-1B9) was subsequently constructed in this laboratory to overcome the ineffective activity of Ad5-based vector in the low CAR expressing cancers. 26 The scFv-M6-1B9 intrabody produced by this adenoviral vector reduced the expression of EMMPRIN and demonstrated its effect on α3β1-integrin and MCT1 functions, which contributes to inhibition of aggressive phenotype in the Caco-2 colorectal cancer cell line. 26 However, the expression of metastatic enzymes MMP-2 and MMP-9 and their inhibitors TIMP-1 and TIMP-2 was not different. 26
The invasiveness of multiple cancer cell types depends on the modulated expression of proteolytic enzymes that digest the extracellular matrix and disengage cell–matrix and/or cell–cell interactions. One of such enzymes, the serine protease uPA, converts enzymatically inactive plasminogen into the widely acting protease plasmin. Plasmin then degrades several extracellular matrix (ECM) components and also activates proenzyme forms of matrix metalloproteases. 38
Cervical cancer is another type of cancer accompanied by high expression of uPA. 39 Therefore, uPA is a particularly important component of cancer invasion and metastasis. In this study, the scFv-M6-1B9 intrabody expressed by Ad5/F35 gene transfer vectors was demonstrated to attenuate the expression of EMMPRIN on the surface of HeLa cells. These results indicated that the scFv-M6-1B9 intrabody successfully suppressed the expression of EMMPRIN on the surface of HeLa cells, as in the previous report using Ad5-based vectors. 25 The authors further investigated the membrane-bound uPA in EMMPRIN-downregulated HeLa cells using Ad5/F35-based vectors in this study.
Downregulation of EMMPRIN cell surface expression on HeLa cells expressing scFv-M6-1B9 intrabody led to a decrease in the membrane-bound uPA. The biological activity of uPA in culture supernatant of EMMPRIN-downregulated HeLa cells was significantly decreased compared to controls. A downregulation of uPA activity was revealed in blocking oral tumor cells with anti-EMMPRIN antibody. 40 Moreover, previous studies have shown that invasion and metastasis of tumor cells were strongly decreased in uPA-deficient mice. 41 The authors further investigated the invasion ability of EMMPRIN-downregulated HeLa cells by Matrigel invasion assay. The invasion ability of EMMPRIN-downregulated HeLa cells with the scFv-M6-B9 intrabody was also decreased in this study.
Taken together, these data indicated that the scFv-M6-1B9 intrabody decreased the membrane-bound uPA, reduced the uPA enzymatic activity in tissue culture media, and inhibited the invasive ability of EMMPRIN-downregulated HeLa cells.
Intrabody and RNAi are two technology platforms for the ablation of protein function, intrabody knockout of protein function at the post-translational level, and RNAi-mediated gene silencing at the post-transcriptional level. 42 It has been reported that more than 1000 genes involved in diverse cellular functions were nonspecifically stimulated or repressed in expression profiling of mammalian tissue culture cells treated under standard conditions with conventional 21-bp siRNAs. 43 In addition, the mutation in codon 121 of the uPA gene resulted in a proline to leucine exchange in 10 out of 22 tumor tissues from ovarian cancer patients. 44 This is a formidable barrier to therapeutic applications of RNAi in humans. RNAi cannot bind to target RNA in which only a point mutation exists.
The use of intrabodies would circumvent this problem, if the mutation does not cause conformational change of the epitope recognized by the intrabody. Intrabodies can block protein expression and function in specific cellular compartments. 45 Endoplasmic reticulum targeting scFv intrabody against transferrin receptor strongly induces cell cycle G1 phase arrest and apoptosis of human breast cancer MCF-7 cells, and consequently, significantly suppresses proliferation of MCF-7 cells. 46 Blocking the transcriptional activity of NF-κB using anti-p65 nuclear targeting intrabody inhibited glioblastoma cell angiogenesis, invasion, and tumor growth via the downregulation of vascular endothelial growth factor (VEGF), MMP-9, uPAR, and uPA. 47 Anti-human telomerase reverse transcriptase nuclear-targeting intrabody specifically neutralized telomere synthesis activity, resulted in inhibiting the viability and proliferation of human cervical adenocarcinoma (HeLa) and hepatoma (HepG2) cells. 48
Recently, animal experiments in a xenograft model of human lung cancer have shown that scFv M97, type IV collagenase transfection exerted marked inhibition on the tumor formation rate and tumor growth, and significantly prolonged survival time of nude mice. 49 Thus, the scFv-M6-1B9 intrabody would be able to be applied in animal experiments and clinical studies in the future.
In conclusion, these results indicated that in addition to reducing the expression of EMMPRIN in the human cervical carcinoma HeLa cells, the scFv-M6-1B9 intrabody decreased the membrane-bound uPA and uPA activity in culture supernatant, and suppressed the invasion ability of the human cervical carcinoma HeLa cell. This study suggested that the scFv-M6-1B9 intrabody might represent a potential approach for invasive cervical cancer treatment. The application of scFv-M6-1B9 intrabody in animal experiments and preclinical studies would be investigated further.
Footnotes
Acknowledgments
This research was supported by grants from the Thailand Research Fund the Commission on Higher Education and the Chiang Mai University, Chiang Mai, Thailand, the Faculty of Associated Medical Sciences, Chiang Mai University, the NSTDA Research Chair Grant, National Sciences and Technology Development Agency (Thailand), the UICC American Cancer Society Beginning Investigators Fellowship funded by American Cancer Society, and the Center of Biomolecular Therapy and Diagnostic, Thailand. The authors thank Ms. Sirikwan Sangboonruang for her technical assistance.
Disclosure Statement
No competing financial interests exist.