
Abstract
Cisplatin resistance is still one of the main reasons for failure of clinical therapy for cervical cancer. But the underlying molecular mechanisms involved in cisplatin resistance of cervical cancer have still remained unclear. Recent studies reported that long noncoding RNAs (lncRNAs) are novel nonprotein-coding transcripts, which might play a key role in cancer biogenesis and prognosis. One of the lncRNAs, urothelial cancer associated 1 (UCA1), has been shown to promote different types of cancer cell proliferation, migration, and invasion. This study showed that overexpression of UCA1 confers cisplatin resistance by promoting cancer cell proliferation and inhibiting apoptosis. In addition, knockdown of UCA1 remarkably decreased cisplatin resistance in cervical cancer cells. Moreover, results also indicated that UCA1 was involved in signaling pathways modulating cell apoptosis and proliferation. UCA1 suppressed apoptosis by downregulating caspase 3 and upregulating CDK2, whereas enhanced cell proliferation by increased level of survivin and decreased level of p21. This study reports for the first time that UCA1 might play an important role in the cisplatin resistance in cervical cancer, and also explain partially how UCA1 promotes cisplatin resistance in cancer cells. These results provide evidence to support that UCA1 can be used as a potential target for a novel therapeutic strategy for cervical cancer.
Introduction
Cervical cancer is one of the most common cancers, which still remains the fourth leading causes of cancer death in females worldwide. 1 Despite strategies for prevention and treatment have rapidly developed in the past decades, the prognosis of patients with advanced or recurrent cervical cancer remains very poor, and survival rate within 1 year is only 10%–20%. 2 Currently, chemotherapy is a widely used treatment for patients with advanced or recurrent cervical cancer, because cervical cancer is extremely chemosensitive. Cisplatin is the most commonly used chemotherapeutic agent. It is a small molecule platinum compound and was first found to inhibit the growth of bacteria and later identified as a potent anticancer drug, which has been shown to be one of the most effective drugs for treating advanced or recurrent cervical cancer. 3 It has been demonstrated that multiple signaling pathways are involved in the molecular mechanisms of anticancer effect of cisplatin. 4,5 . Among these mechanisms, its ability to crosslink with the purine bases on the DNA molecule, interfering with DNA repair, eventually leading to severe DNA damage and inducing apoptosis and necrosis in cancer cells, has been well established. However, cisplatin resistance is still the biggest challenge in the improvement of the long-term curative effect, and the mechanism of cisplatin resistance in cervical cancer is not yet clear.
Recent investigations have reported that long noncoding RNAs (lncRNAs) are involved in different types of pathological conditions, such as Mendelian disorders, cardiovascular disease, neurological disorders, and cancer. 6 LncRNAs are transcripts greater than 200 nucleotides, which are transcribed by RNA polymerase II but do not encode proteins. It has been shown that lncRNAs modulate a number of physiological processes including gene transcription, cell proliferation, cell differentiation, and chromosome inactivation. 7,8 In addition, increasing number of studies have suggested that lncRNAs might be the potential drivers in the development of cancers and can be used as clinical biomarkers for the prognosis of cancers. 9
Urothelial cancer associated 1 (UCA1) is an lncRNA that has been given more and more attention in recent years. It has been indicated that UCA1 is highly expressed in a broad range of cancer tissues and plays a crucial regulatory role in the pathogenesis of cancer development, including tumors growth and metastasis. 10 Thus, UCA1 might be considered as a therapeutic target or biomarker for cancers. 11 Moreover, it has been proven that UCA1 is associated with cisplatin resistance in cancer cells. 12 However, the underlying molecular mechanisms for the regulatory roles of UCA1 in different types of cancer are still unknown, also there are very few studies on the effect of UCA1 expression in cervical cancer.
In this study, the functional role of UCA1 expression was evaluated in the chemosensitivity of cervical cancer cells to cisplatin. It was suggested that UCA1 expression promotes cisplatin resistance in cervical cancer cells through signaling pathways regulating cell apoptosis and proliferation, which are involved in a balancing expression of caspase-3, p21, CDK2, and survivin. This study indicates for the first time that UCA1 plays a crucial regulatory role in the cisplatin resistance in cervical cancer cells, and also explains partially how UCA1 promotes cisplatin resistance in cervical cancer cells. These results provide evidence to support that UCA1 can be used as a potential target for a novel therapeutic strategy for cervical cancer.
Materials and Methods
Cell culture
The HeLa cell line was cultured in Dulbecco's modified Eagle's medium (DMEM) with 10% (v/v) fetal bovine serum (FBS), 100 U/mL penicillin, 100 mg/mL streptomycin, and 2 mM glutamine. Cells were incubated in a 5% carbon dioxide incubator at 37°C. At 70%–80% confluence, cells were splitting according to standard procedures. In certain groups, cells were treated with cisplatin (DDP) (0.5, 1.0, 2.0, 4.0, 8.0, and 16.0 μg/mL) for 24 hours. IC50 was calculated and then cells were harvested in DMEM with 1% FBS for preparation of the following biochemistry analysis. Antibody of caspase-3 (#9665; CST), survivin (#2808; CST), p21 (#ab109520; Abcam), and CDK2 (#ab32147; Abcam) was selected.
Establish the cisplatin resistance model in HeLa cells
To study the effect of UCA1 in cisplatin resistance of cervical cancer cells, the progressively increasing DDP concentration method was utilized to establish a cisplatin resistance HeLa cell model. Cells were treated with progressively higher concentrations of DDP (0.1, 0.2, 0.4, 0.6, 0.8, 1.0, and 2.0 μg/mL). After 2 weeks screening, cisplatin resistance HeLa cells (Hela/DDP) were obtained. HeLa/DDP cells were later treated with higher concentration of DDP (2, 4, 8, 16, 32, and 64 μg/mL). IC50 was calculated and cells were harvested for preparation of the following biochemistry analysis.
RNA extraction and real-time polymerase chain reaction
The total RNA from HeLa cells was extracted by using Trizol reagent (Invitrogen, CA). The first-strand cDNA was synthesized from 1 μg of total RNA using the Reverse Transcription System Bestar quantitative PCR reverse transcription kit (Bestar, Shanghai, China) according to the manufacture's instruction. Real-time polymerase chain reaction (PCR) was carried out with an ABI 7500 Real-Time PCR system. Each assay was performed in triplicate, and GAPDH was used as the endogenous control gene. The primer sequences used were as follows: UCA1, 5′-CCACACCCAAAACAAAAAATCT-3′ (forward) and 5′- TCCCAAGCCCTCTAACAACAA −3′ (reverse); GAPDH, 5′- TGTTCGTCATGGGTGTGAAC-3′ (forward) and 5′- ATGGCATGGACTGTGGTCAT-3′ (reverse). The relative amount of UCA1 to GAPDH was calculated using with a 2−ΔΔCt method and normalized using GAPDH cDNA as an internal control.
UCA1 knockdown and overexpression
The siRNA–UCA1 (5′-GAGCCGAUCAGACAAACAAUU-3′) and eukaryotic expression vector pcDNA3.0–UCA1 were obtained commercially (Genepharma Company, Shanghai). Transfections were performed using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's instructions.
Determination of cell proliferation by cell counting kit 8 (CCK8) and EdU assay
Cell proliferation was evaluated by cell counting kit 8 (CCK-8) and ethynyl-2-deoxyuridine incorporation assay using an EdU Apollo DNA in vitro kit (RIBOBIO, Guangzhou, China) according to the manufacturer's protocol. HeLa cells were cultured at a density of 5 × 104 per well in a 96-well culture dish. After adherence, different groups of cells were treated with 1 μg/mL DDP for 12, 24, and 48 hours. UCA1–siRNA or UCA1–pcDNA3.0 vector was transfected into HeLa cells before DDP treatment, if necessary. At the end of each time point, CCK-8 solution was added to each well, and then cells were further incubated for 2 hours at 37°C. The absorbance of samples at 450 nm was determined by a multiwell plate reader.
For EdU assay, at the end of each time point, cells were incubated with 100 μL of 50 μM EdU per well for 2 hours at 37°C. Then, the cells were fixed for 30 minutes at room temperature using 100 μL of fixing buffer (4% polyformaldehyde containing PBS). Subsequently, the cells were incubated with 50 μL of 2 mg/mL glycine for 5 minutes followed by washing with 100 μL of PBS. After permeabilization with 0.5% Triton X, the cells were reacted with 1X Apollo solution for 30 minutes at room temperature in the dark. After that, cells were incubated with 100 μL of 1X Hoechst solution for 30 minutes at room temperature in the dark followed by washing with 100 μL of PBS. The cells were then analyzed using flow cytometry.
Assay of apoptosis and cell cycle by flow cytometry
For apoptosis analysis, cell apoptosis utilizing Annexin V and propidium iodide (PI) double staining was determined. HeLa cells treated with different experimental conditions were detached with trypsin–EDTA, washed with PBS, and then resuspended in binding buffer (10 mM HEPES pH7.4, 150 mM NaCl, 5 mM KCl, 1 mM MgCl2, and 1.8 mM CaCl2) containing FITC-annexin V (1 g/mL) and further incubated for 20 minutes. Ten minutes before the end of incubation, PI (10 g/mL) was added to this cell suspension to stain necrotic cells. Cells were analyzed with an FACS flow cytometer equipped with an excitation laser at 488 nm. The PI was collected through a 575 nm band pass filter. For cell cycle analysis, cells were stained with PI staining solution (10 μg/mL RNase A and 50 μg/mL PI) at 37°C for 30 minutes in the dark. Cell cycle distribution was analyzed using flow cytometry provided with the Cell-Quest software.
Western blot analysis
For Western blot analysis, frozen cells were sonicated on ice twice for 5 seconds in 50 mM lysis buffer (pH 7.4, 3.1 mM sucrose, 1 mM DTT, 10 μg/mL leupeptin, 10 μg/mL soybean trypsin inhibitor, 2 μg/mL aprotinin, and 0.1% Triton X-100). Homogenates were centrifuged at 10,000 g at 4°C for 20 minutes and the supernatants were collected. The total protein concentration was measured using the Bradford protein assay (Bio-Rad, Hercules, CA). Protein lysates (30 μg) were separated using 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene fluoride (PVDF) membrane. After blocking with 1% bull serum albumin (BSA), the PVDF membrane was incubated overnight with the primary antibody as follows: monoclonal antibodies against caspase-3 (#9665; CST), survivin (#2808; CST), p21 (#ab109520; Abcam), and CDK2 (#ab32147; Abcam) solute in PBS-T with 1% BSA. Membranes were washed in PBS-T (10 minutes) for three times and then probed with the appropriate secondary antibody. Membranes were developed using Versa Doc 5000 and band densities were measured with Quantity One 4.6 software (Bio-Rad). Equal protein loading was additionally verified by measurement of GAPDH level with mouse monoclonal antibody.
Statistical analysis
Statistical calculations were performed using Prism 6 (GraphPad, San Diego, CA). Data are presented as the mean ± SEM. Student's t tests were used for comparisons between two groups, and one-way analysis of variance was used for comparisons among multiple groups. *p < 0.05, **p < 0.01 indicated statistical significance.
Results
The expression level of UCA1 in HeLa cells and HeLa/DDP cells
To investigate the cisplatin resistance mechanism in cervical cancer cells, the cisplatin (DDP)-resistant HeLa cell line was established by using progressively increasing dose of DDP and high-dose stimulation. The HeLa/DDP cells with high cisplatin resistance were successfully obtained. The value of IC50 for DDP in HeLa/DDP cells is 8.03 μg/mL, which is much higher than that in wild type HeLa cells, DDP IC50 = 1.74 μg/mL (Tables 1 and 2).
The relative expression level of UCA1 was determined by quantitative real-time PCR in HeLa cells. HeLa cells were treated with 0.5 or 1.0 μg/mL DDP for 48 hours. UCA1 expression in the DDP-treated group significantly decreased by 31% and 53%, respectively, in comparison with the control group (Fig. 1A).
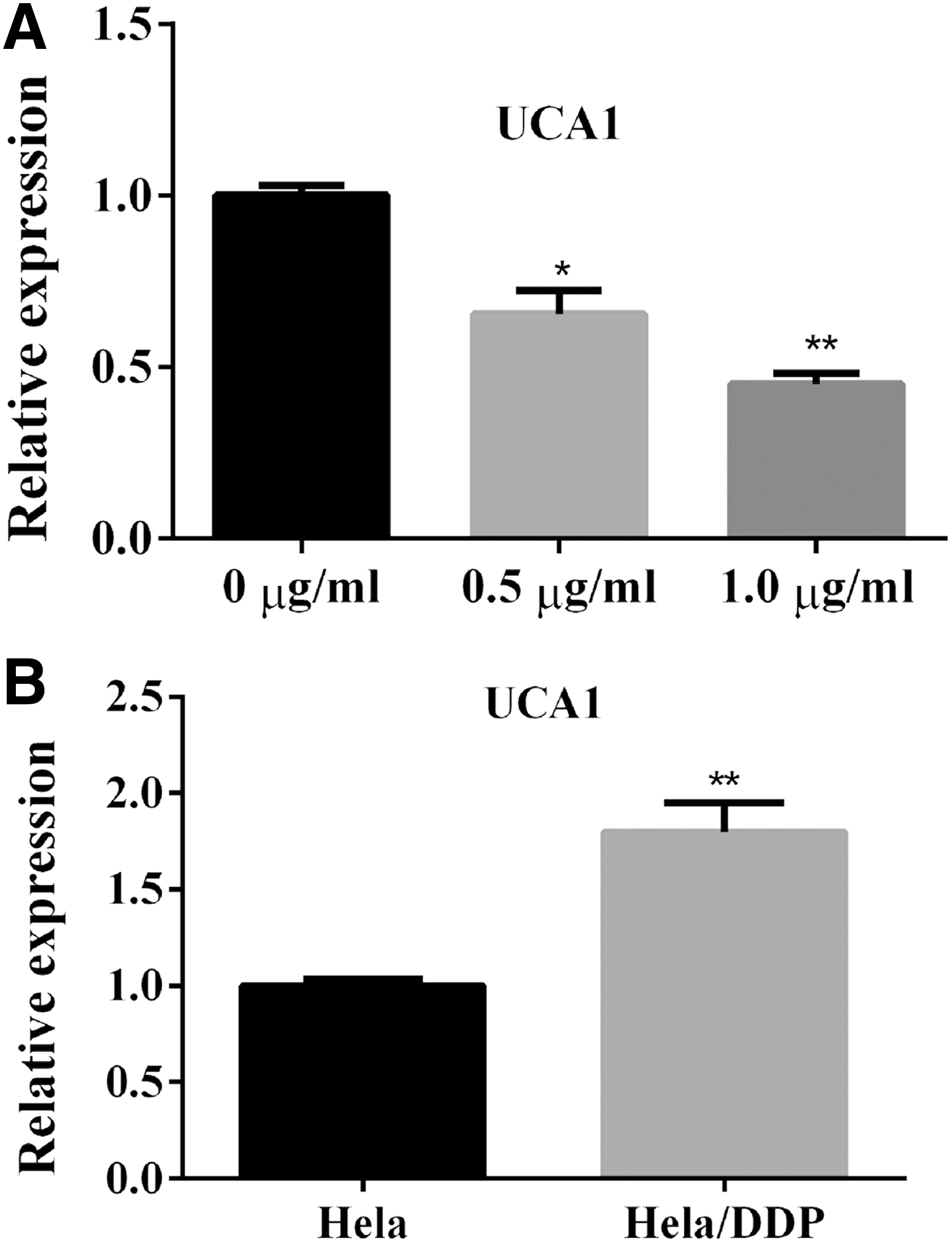
The effect of DDP treatment on the expression of UCA1. The relative expression level of UCA1 was determined by qRT-PCR.
Data in this study have shown that the cisplatin resistance model in HeLa/DDP cells has a much higher IC50 value than the wild type HeLa cells. To investigate whether UCA1 is associated with cisplatin resistance, the relative expression level of UCA1 was also measured in wild type HeLa cells and HaLe/DDP cells after 24 hours DDP treatment. Result from qRT-PCR assay showed that UCA1 expression remarkably increased compared with that of wild type HeLa cells (Fig. 1B).
Overexpression of UCA1 inhibited apoptosis in HeLa cells and protected cell proliferation
Specific plasmid vector pcDNA3.0–UCA1 or pcDNA3.0 empty was transfected into HeLa cells. At 48 hours after transfection, the relative expression level of UCA1 was evaluated by qRT-PCR. In comparison with the empty pcDNA3.0 negative control group, the expression level of UCA1 in the pcDNA–UCA1 group markedly rose fivefold (Fig. 2A).

The effect of overexpression of UCA1 in DDP-treated HeLa cells.
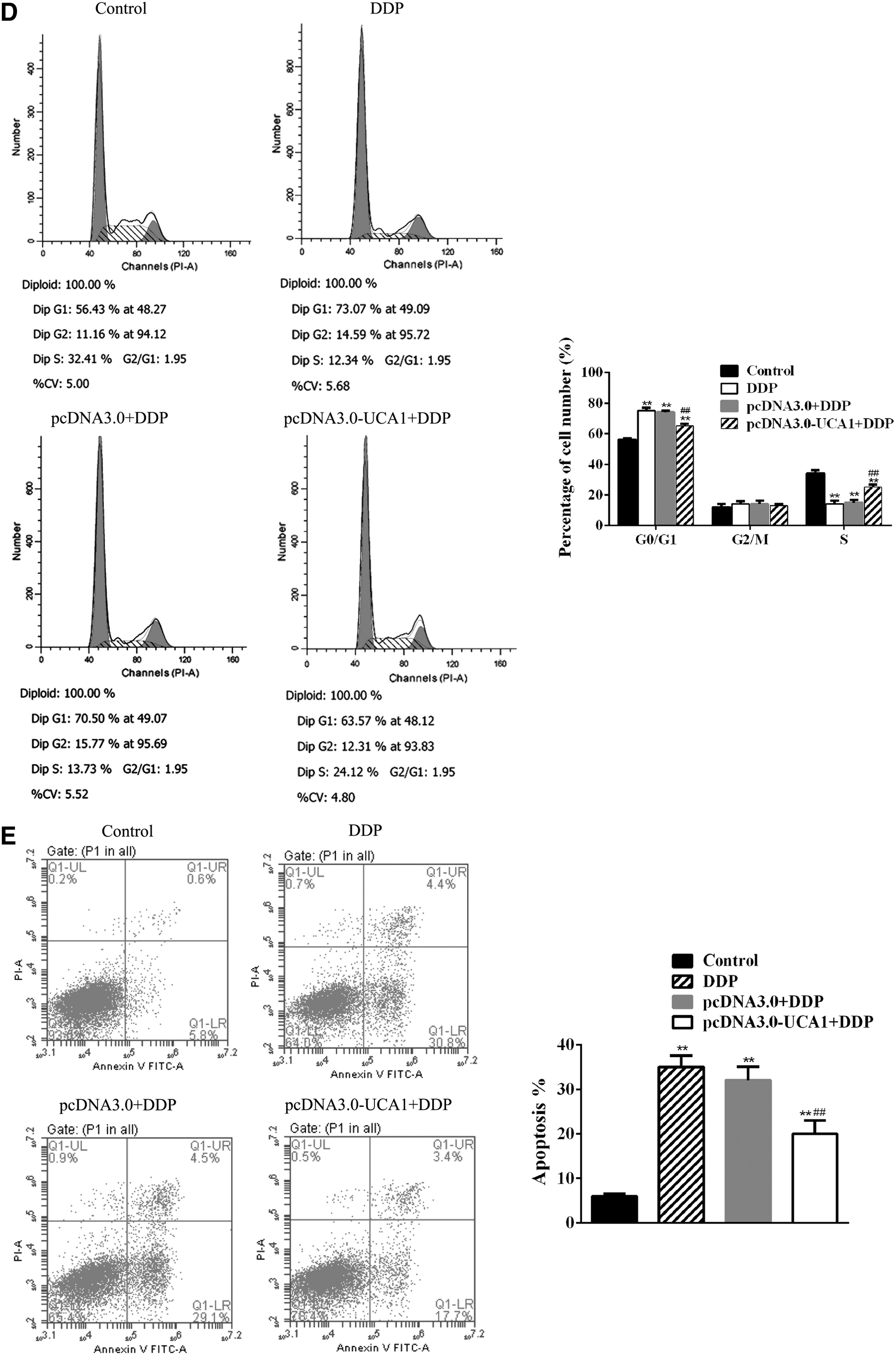
To explore the possible effect of UCA1 overexpression on the growth of cervical cancer cells, cell proliferation activities of HeLa cells were analyzed by CCK-8 assay, EdU assay, and flow cytometry. Results from CCK8 indicated that Pcdna3.0–UCA1-transfected HeLa cells have a better performance in cell viability and proliferation after 24 hours DDP treatment than control HeLa cells and pcDNA3.0 empty transfected HeLa cells (Fig. 2B). Similar results were obtained from both EdU assay and flow cytometry analysis. Cell growth and proliferation were protected in pcDNA3.0–UCA1-transfected HeLa cells after 24 hours DDP treatment (Fig. 2C, D).
Control HeLa cells, pcDNA3.0–UCA1-transfected HeLa cells, and pcDNA3.0 empty transfected HeLa cells were treated with 1.0 μg/mL DDP for 24 hours. Apoptosis rate was analyzed by flow cytometry. In comparison with nontreated HeLa cells, the apoptosis rate was remarkably increased in control HeLa cells after DDP treatment. Similar result was obtained in pcDNA3.0 empty transfected HeLa cells after DDP treatment. However, a significant reduction of apoptosis rate in pcDNA3.0–UCA1-transfected HeLa cells after DDP treatment was observed, which decreased by 33% compared with control HeLa cells and pcDNA3.0 empty transfected HeLa cells (Fig. 2E).
UCA1 overexpression varied the level of apoptosis-related protein and cell survival factors
Quantitative PCR analysis indicated that the expression level of caspase 3 and p21 in control HeLa cells and pcDNA3.0 empty transfected HeLa cells was significantly increased after treatment with DDP in comparison with that in nontreated HeLa cells. Whereas pcDNA3.0–UCA1-transfected HeLa cells had a lower expression of caspase3 and p21 after DDP treatment than the other two DDP-treated groups. However, the expression of survivin and CDK2 in pcDNA3.0–UCA1-transfected HeLa cells remained at a similar level as that of the nontreated HeLa cells, whereas a dramatic drop was observed in control HeLa cells and pcDNA3.0 empty transfected HeLa cells (Fig. 3A). Similar results were also obtained by Western blot (Fig. 3B).

The expression level of caspase-3, p21, survivin, and CDK2.
Knockdown of UCA1 inhibited cell proliferation and induced apoptosis in HeLa/DDP cells
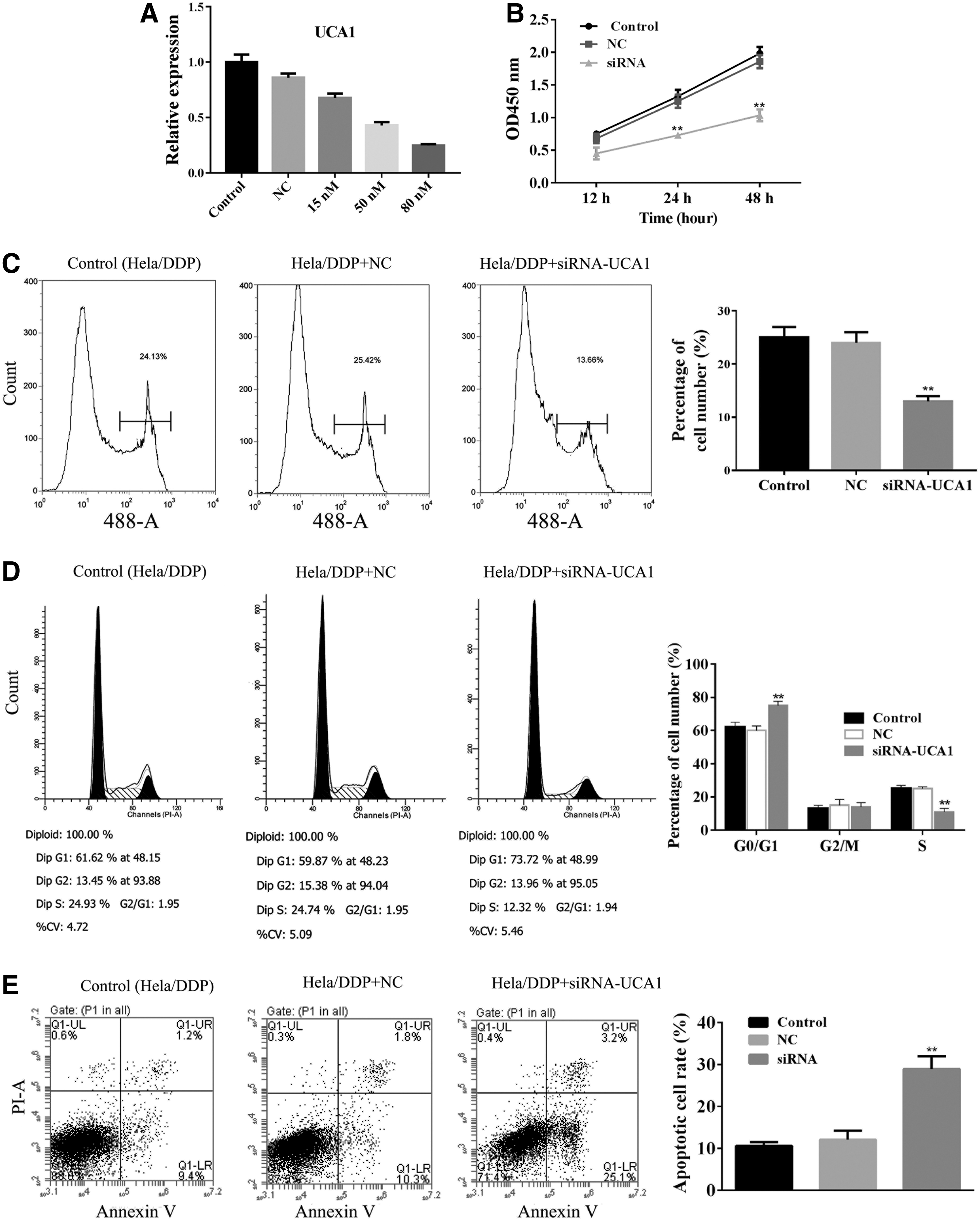
Knockdown of UCA1 expression by UCA1–siRNA was evaluated by qRT-PCR analysis, which showed a significant reduction of UCA1 expression in HeLa/DDP cells (Fig. 4A).
The effect of UCA1 knockdown in HeLa/DDP cells.
The cell proliferation activities of HeLa/DDP cells were also determined by CCK-8 assay, EdU assay, and flow cytometry. CCK-8 revealed that cell proliferation in UCA1–siRNA-transfected HeLa/DDP cells remarkably decreased by ∼50% in comparison with that in control HeLa/DDP cells and NC siRNA-transfected HeLa/DDP cells (Fig. 4B). Similar observations were obtained from both EdU assay and flow cytometry analyses. Cell growth and proliferation were remarkably suppressed in UCA1–siRNA-transfected HeLa/DDP cells (Fig. 4C, D).
Flow cytometry analysis revealed that the apoptosis rate was remarkably increased in UCA1–siRNA-transfected HeLa/DDP cells than in control HeLa/DDP cells and NC siRNA-transfected HeLa/DDP cells (Fig. 4E).
Upregulated apoptosis-related protein and downregulated cell survival factors were induced by UCA1 knockdown in HeLa/DDP cells
The expression levels of caspase-3, p21, survivin, and CDK2 were also evaluated by qPCR analysis. In comparison with control HeLa/DDP cells and NC siRNA-transfected HeLa/DDP cells, the levels of caspase-3 and p21 were significantly increased, whereas the levels of survivin and CDK2 were decreased (Fig. 5A). Similar results were also obtained by Western blot (Fig. 5B).
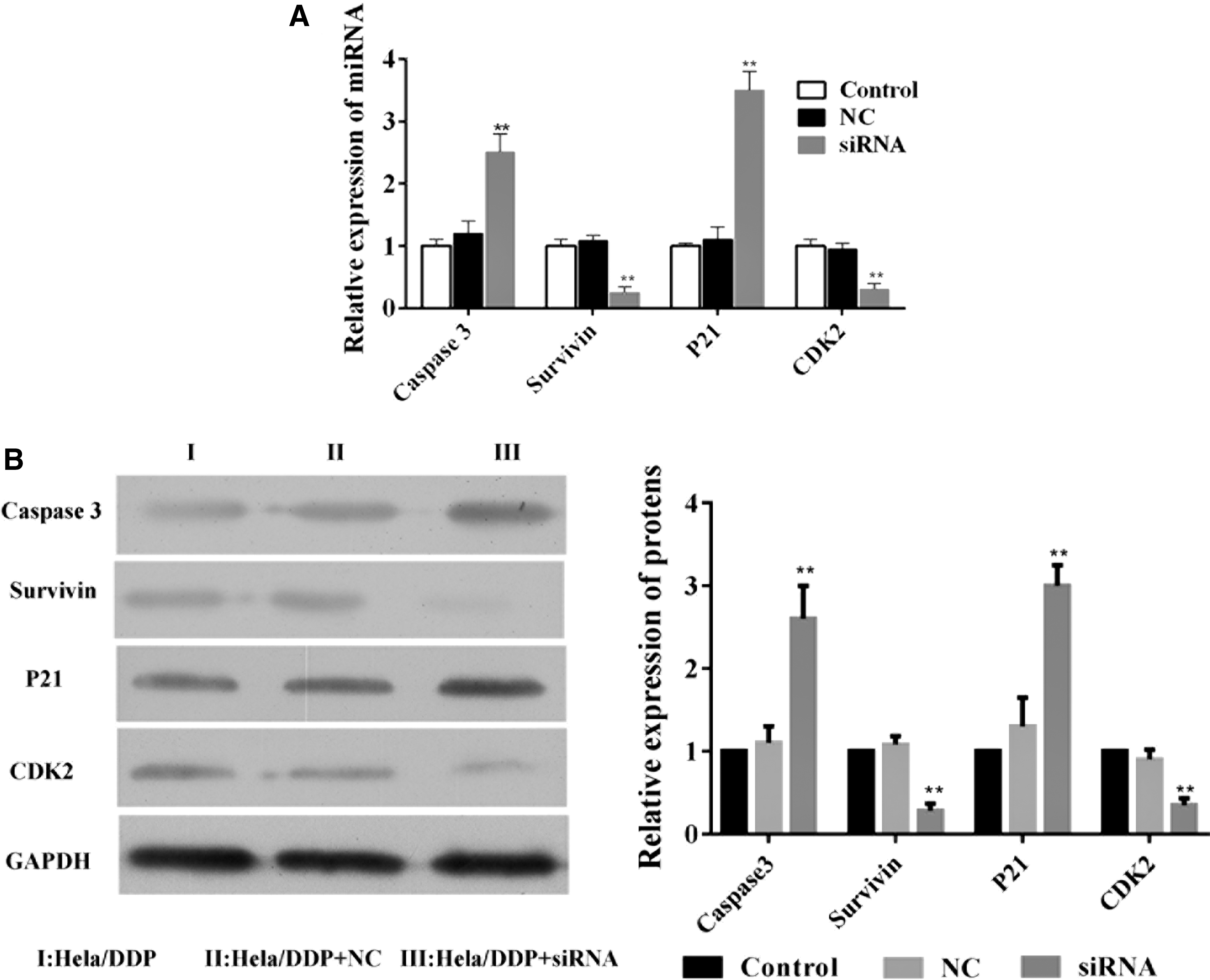
The expression level of caspase-3, p21, survivin, and CDK2.
Discussion
Multiple therapeutic and prevention strategies against cervical cancer have been introduced and widely used, including surgery, 13 chemotherapy, 14 radiation therapy, 15 proton therapy, 16 hormone therapy, 17 RNA interference therapy, and vaccine prevention. 18 However, cervical cancer still remains one of the leading causes of cancer death, especially in developing countries. The traditional strategy of treating advanced cervical cancer is radiation therapy combined with cisplatin chemotherapy. Cisplatin is the most commonly used chemotherapy agent, but cisplatin resistance remains a major unsolved problem, causing relapsed cisplatin-resistant disease and leading to more than 80% of uncured advanced or recurrent cervical cancer. 2 Mechanisms of cisplatin resistance have been proposed by a number of investigations, including cellular uptake and efflux variation, detoxification, and apoptosis inhibition. 19 In this study, by using cisplatin resistance in HeLa/DDP cell model, a different mechanism of cisplatin resistance through lncRNA UCA1 was found, which is crucially associated with the development of cisplatin resistance in cervical cancer cells.
LncRNAs have emerged as a crucial player in multiple types of cancers. Pieces of evidence suggest that different lncRNAs play different roles in cancer development and they might function as tumor suppressors or oncogenes. 20 UCA1 as one of the most studied lncRNAs is commonly considered as encogene. 21 UCA1 has been shown to promote the progression of cancer, including tumor growth, 22 apoptosis inhibition, 23 tumor cell migration, 24 metabolism, 25 and metastasis, 26 and affecting many different types of cancers. 10 However, very few studies have focused on the role of UCA1 in cervical cancer. In this study, the functional role of UCA1 was investigated in HeLa cells and provided evidence supporting that UCA1 expression plays an important role in the pathogenesis of cervical cancer.
This study shows that UCA1 is downregulated in DDP-treated HeLa cells, whereas upregulated in the cisplatin resistance model in HeLa/DDP cells. This phenomenon suggests that DDP as chemotherapy agent can reduce the expression of UCA1 at early stage of cervical cancer treatment, resulting in initial curative effect; whereas after a long-term high-dose treatment, UCA1 expression recovers or even upregulates and cells become cisplatin resistant, leading to cisplatin-resistant diseases and low efficiency of chemotherapy in cervical cancer. Multiple molecular mechanisms of UCA1 enhancing cancer progression have been reported, which are associated with AKT-Bax-Bcl-2 pathway, 27 FGFR1/ERK pathway, 28 TGF-β signaling pathway, 29 and Wnt signaling pathway. 30 In this study, cell proliferation activity assay revealed that UCA1 expression dramatically enhanced cell growth and proliferation but inhibited HeLa cell apoptosis, leading to remarkably increased survival rate in HeLa cells. Western blot analysis showed decreased level of apoptosis-related protein caspase3 and p21, and increased level of cell cycle-related protein survivin and CDK2. These results support that UCA1 does play a functional role in cervical cancer, which at least partially associate with caspase-3/p21 apoptosis pathway and survivin–CDK2 cell cycle cascade.
Recent studies have indicated that UCA1 is implicated in cisplatin resistance in bladder cancer 31 and ovarian cancer. 11 In this study, a cisplatin resistance model was established in HeLa/DDP cells, and UCA1 expression was knocked down using UCA1–siRNA. Cell proliferation activity assay demonstrated that knockdown of UCA1 in HeLa/DDP cells significantly impacted the pathological homeostasis of HeLa/DDP cells, which resulted in more apoptotic cells, suppressing cell growth and proliferation and decreased cell survival rate. Fan et al. reported that UCA1 expression induces cisplatin resistance through activating Wnt signaling pathway, 30 and Wang et al. suggested that cisplatin resistance is related to the expression of serine/arginine-rich protein-specific kinase1. 26 These results provide clear evidence that caspase 3/p21-activated apoptotic cascade and survivin/CDK2-related cell cycle signaling pathway play crucial roles in generating cisplatin-resistant cervical cancer cells.
To the best of our knowledge, this is the first study to determine how UCA1 plays an important role in the progression of cervical cancer and the development of cisplatin resistance through inhibiting apoptotic pathway and enhancing cell proliferation. These results suggest that UCA1 may be considered as a therapeutic target for a novel therapeutic strategy against cervical cancer in the future.
Footnotes
Acknowledgments
This study was supported by a grant from the Academician Workstation of Maternal and Child Health Hospital of Guizhou Province, Talents in the platform of Scientific Cooperation in Guizhou Province ([2016]5603); the National Natural Science Foundation of China (No.81460365); the program for Scientific Cooperation in Guizhou Province ([LH][2015]7377),([LH][2016]7405) and ([LH][2015]7382); the Science and technology program for Guiyang City Health and Family Planning Commission [2016]002; and support by Guiyang City Science and Technology Bureau [20151001]41.
Disclosure Statement
No competing financial interests exist.