
Abstract
Background:
MicroRNA-516b (miR-516b) has been recently reported to be downregulated in nonsmall cell lung cancer (NSCLC). However, its clinical significance and biological function in NSCLC remain to be clarified.
Materials and Methods:
Quantitative real-time polymerase chain reaction (qRT-PCR) was used to detect the expression of miR-516b in 82 paired fresh primary tumor tissues and NSCLC cell lines. The association of miR-516b expression with clinicopathological factors and prognosis was statistically analyzed by SPSS 21.0 software, Kaplan–Meier method, and Cox regression analyses. Cell Counting Kit-8, colony formation, flow cytometric, Transwell migration, and invasion assays were used to evaluate the proliferation, cell cycle, apoptosis, migration, and invasion of NSCLC cells after miR-516b mimics or negative control of mimics transfection.
Results:
The expression level of miR-516b was found to be significantly lower in NSCLC tissues and cell lines than in corresponding normal tissues and cells. Decreased miR-516b expression was significantly associated with tumor size (p = 0.004), Tumor Node Metastasis (TNM) stage (p = 0.016), and shorter overall survival (p = 0.0039). Multivariate analysis suggested that miR-516b was an independent risk factor for NSCLC (hazard ratio = 2.435, 95% confidence interval: 1.423–2.457; p = 0.003). Furthermore, overexpression of miR-516b could inhibit NSCLC cell proliferation, cell cycle progression, migration and invasion, and promoted cell apoptosis. The qRT-PCR results indicated that overexpressing miR-516b reduced the mRNA expression of CDK2, MMP-2, and MMP-9, whereas increased BAX mRNA expression in NSCLC cells. Their protein expression levels presented similar trends, as confirmed by Western blotting.
Conclusions:
Findings in this study demonstrated for the first time that miR-516b expression might be a novel diagnostic and prognostic factor, as well as a promising target for NSCLC.
Introduction
Lung cancer is the most common cause of tumor-related death worldwide in males and females, which has been classified into two main types: small cell lung cancer and nonsmall cell lung cancer (NSCLC). 1 As the main subgroup of lung cancer, NSCLC could account for 85% of all lung cancers diagnosed. 2 Even though recent advances have been made in clinical and experimental studies for the NSCLC, the 5-year survival rates still remain relatively poor for advanced NSCLCs. 3,4 Currently, several prognostic factors, including Tumor Node Metastasis (TNM) classification, tumor size, and node, are the basis for prognostic management for NSCLC. However, sufficient information is still scarce about biological tumor progression. Therefore, novel biomarkers urgently need to be identified for better understanding of the molecular mechanisms involved in the NSCLC development and to improve the prognostic prediction of NSCLC.
Currently, RNA interference, including small interfering RNAs (siRNAs) and microRNAs (miRNAs; miRs), is a good way to study the expression of certain genes with fewer side-effects and greater target specificity than conventional medicines, and has been successfully used in related studies of bacteria, animals and humans. 5 Compared with siRNAs, miRs, including miR mimics (agonist) and anti-miR (antagonist), are a simple and efficient tool to study the gain-of-function or loss-of function assays in cells because it has not complicated operation of constructing vector. 6 miRs are a class of single-stranded, noncoding small RNAs with 18–25 nucleotides, 7 which have been reported to act as key regulators of diverse biological processes, including embryonic development, cell proliferation, and endocrine homeostasis. 8,9 There are accumulating pieces of evidence to suggest that miRs function as oncogenes or tumor suppressors by negatively regulating gene expression through base pairing with the 3′ untranslated region of their target mRNA. 10,11 For example, miR-346 has been demonstrated to be a novel oncogenic miR that facilitates NSCLC cell growth and metastasis in vitro and in vivo. 12 On the contrary, Zhang et al. found that miR-143 was able to suppress NSCLC cell proliferation and invasion by inhibiting the effects of epidermal growth factor receptor (EGFR). 13 Notably, a recent study by Hu et al. found that miR-516b was significantly downregulated in NSCLC tissues using integrative miR data analysis. 14 But to date, the clinical significance and function attributes of miR-516b in NSCLC have not been investigated. Therefore, this study was to investigate the expression of miR-516b in NSCLC and its clinical significance. Furthermore, the effects of miR-516b on proliferation, apoptosis, migration, and invasion of NSCLC cells were also evaluated using miR mimics, which will help us to have a better understanding of the functions of miRs in NSCLC tumorigenesis.
Materials and Methods
Clinical tissues sample and cell lines
A total of 82 paired fresh primary NSCLC tumor tissues and corresponding normal tissues were obtained for quantitative real-time polymerase chain reaction (qRT-PCR) analysis. These specimens were all collected from patients with NSCLC at Tai Zhou Central Hospital, Zhejiang, China, which fulfilled the following criteria: (1) histologically diagnosed primary NSCLC, (2) no history of familial synchronous malignancy, and (3) available clinical and follow-up data. The clinical stage was evaluated according to the seventh edition of the International Union Against Lung Cancer (UICC, 2009). All participants provided written informed consent and their clinical data are summarized in Table 1. This study was approved by the ethics committee at the local hospital.
Bold values indicate significant differences.
Four human NSCLC cell lines (SPCA-1, A549, H69, and H1299) and one normal human bronchial epithelial cell line, BEAS-2B, were all purchased from Cell Bank of the Chinese Academy of Sciences. All cell lines were cultured in RPMI 1640 medium with 10% fetal bovine serum (FBS; Gibco) in a sterile incubator containing 5% CO2 at 37°C. The medium was changed at least once in 3 days.
RNA extraction and qRT-PCR analysis
Total RNA was extracted from tissues and cell lines using Trizol reagent (Invitrogen, Carlsbad, CA) and reversed transcribed using the GoScript™ reverse transcription kit (Promega) according to the manufacturer's instructions. TaqMan miR probes (Applied Biosystems, Foster City, CA) were used to quantify miR-516b. The relative expression level of miR-516b, CDK2, BAX, MMP-2, and MMP-9, was calculated and statistically compared using the 2−ΔΔCt method after being normalized with the U6 snRNA or GAPDH expression. The primer sequences used in this study are listed in Table 2. Each sample was performed in triplicate.
Western blot analysis
After cell transfection, A549 cells were harvested and the protein was extracted from cells according to the previously reported method. 15 Then the lysed protein was quantified using a BCA protein assay kit and separated by a denaturing SDS-PAGE. The gels were transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA) and blocked with 5% nonfat milk for 2 hours. Subsequently, the membranes were incubated with the primary antibodies against CDK2, BAX, MMP-2, and MMP-9 (1:500; Proteintech, Proteintech Group) and GAPDH (1:5000; Proteintech, Proteintech Group) overnight at 4°C. After incubation with antirabbit secondary antibody at room temperature for 2 hours, proteins were observed using enhanced chemiluminescence kit (Amersham Pharmacia Biotech, Little Chalfont, United Kingdom). GAPDH was used as an internal control.
Oligonucleotide transfection
The miR-516b mimics and miR negative control (NC) of mimics were purchased from Applied Biosystems. For cell transfection, A549 cells were seeded in 12-well plates and incubated overnight, then transiently transfected with miR-516b mimics or NC using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) following the manufacturer's instruction. The nontreated cells were used as control group.
Cell viability assay
Cell Counting Kit-8 (CCK-8; Dojindo) was used to determine the viability of A549 cells according to the manufacturer's instructions. In brief, cells (2000 cells per well) were seeded in 96-well plates and incubated at 37°C after transfection. After 48 hours transfection, cells were added to 10 μL CCK-8 solution and incubated for 2 hours at 37°C. The absorbance of each well at 450 nm was measured using a spectrophotometer. Cell viability was assessed daily for 4 days consecutively and each sample was performed in triplicate.
Colony formation assay
For the colony formation assay, A549 cells were seeded in six-well plates after transfection. After culture for 6 days to form natural colonies, cells were fixed in 4% paraformaldehyde for 10 minutes and stained with crystal violet. The stained colonies (at least 50 cells per colony) were recorded with a digital camera and counted using imaging analysis software (Wimcology, Wimasis).
Cell cycle and apoptosis analysis
Flow cytometry (FACScan) was performed to analyze the effect of miR-516b on cell cycle progression and apoptosis in A549 cells. For cell cycle, cells were harvested at room temperature after 48 hours transfection. Then cells were trypsinized, fixed with 70% ice-cooled ethanol, and stained with 100 μL of 100 μg/mL propidium iodide. Subsequently, the percentage of cells in different cell cycle phases (G0/G1, S, and G2/M) was analyzed by flow cytometer (BD). Apoptosis was detected by Annexin V−APC/7-AAD apoptosis detection kit (KeyGEN Biotech, Nanjing, China) following the manufacturer's instructions. For cell apoptosis, cells were categorized into viable cells (Annexin V−/7-AAD−), dead cells (Annexin V−/7-AAD+), early apoptotic cells (Annexin V+/7-AAD−), and late apoptotic cells (Annexin V+/7−AAD+). The apoptotic percentage in miR-516b mimics group was compared with the control or NC group. Each sample was analyzed in triplicate.
Cell migration and invasion assay
To further confirm whether miR-516b affected NSCLC metastasis, the migration and invasive ability of the cells were investigated using Transwells (Corning Costar Corp., Bedford). For migration assay, A549 cells (50,000) from different groups (control, NC, and miR-516b mimics) were seeded into the upper chambers. Approximately 400 μL RPMI 1640 medium containing 10% FBS was added to the bottom chambers. After incubation for 48 hours at 37°C, cells that migrated to the bottom surface of the chamber were fixed with 2% paraformaldehyde for 20 minutes, and stained with 0.1% crystal violet for 30 minutes. Then, the cells were observed and counted under a light microscope for at least three random fields. For invasion assay, the upper chambers were first covered with 1 mg/mL Matrigel and cells were analyzed according to a protocol similar to that of the migration assay. Each sample was repeated three times.
Statistical analysis
The expression of miR-516b mRNA in NSCLC tissues and corresponding noncancerous tissues was analyzed using Mann–Whitney U-test. The Chi-square test was used to analyze the association between miR-516b expression and clinicopathological parameters of NSCLC. The Kaplan–Meier method was applied to estimate the differences in overall survival rate using log-rank test. The Cox regression analysis was used for multivariate analysis. For cellular experiments, data were expressed as means ± standard deviation. All statistical analyses were performed using SPSS 21.0 software and a p-value less than 0.05 was considered to be statistically significant.
Results
Associations between miR-516b expression and clinicopathological features in NSCLC
To investigate the role of miR-516b in NSCLC, the expression of miR-516b was first determined in 82 paired NSCLC tissues and normal tissues using qRT-PCR. As shown in Figure 1A, the expression of miR-516b was clearly found to be downregulated (0.545 ± 0.32) in tumor tissues in comparison with the paired noncancerous tissues (1.642 ± 0.226). To further explore the potential clinical significance of miR-516b in NSCLC patients, a total of 82 NSCLC patients were divided into miR-516b low-expression group (n = 24) and mid/high-expression group (n = 58) according to the median expression of miR-516b in tissues (0.545). The associations between clinicopathological features and miR-516b expression are summarized in Table 1. The expression of miR-516b was positively associated with tumor size (p = 0.004), TNM stage (p = 0.016), and tissue source (p = 0.0139). However, it has no correlations with gender, age, histology, smoking history, and lymph node (LN) metastasis. Based on these findings, it was speculated that miR-516b might play a vital role in NSCLC progression.

Expression of miR-516b was significantly downregulated in NSCLC tissues and cell lines.
To roundly confirm the miR-516b expression in NSCLC in vitro, the expression of miR-516b was detected in several NSCLC cell lines (SPCA-1, A549, H69, and H1299) and normal human bronchial epithelial cell line BEAS-2B. As shown in Figure 1B, it was observed that the miR-516b expression was clearly downregulated in four NSCLC cell lines compared with normal cell line BEAS-2B. Notably, A549 cell line was chosen for further analysis for expressing the lowest levels of miR-516b expression among four NSCLC cell lines.
Decreased expression of miR-516b predicted poor prognosis in patients with NSCLC
Considering that the level of miR-516b expression was significantly correlated with tumor size and TNM stage, it was predicted that miR-516b might affect the prognosis of NSCLC patients. Thus, the Kaplan–Meier method with log-rank test analysis was used. As shown in Figure 2, the median overall survival for all patients was 20 months. Patients in the high expression group achieved significant survival prolongation compared with those in the low expression group (median overall survival, 23.5 months vs. 17.0 months; p < 0.05).
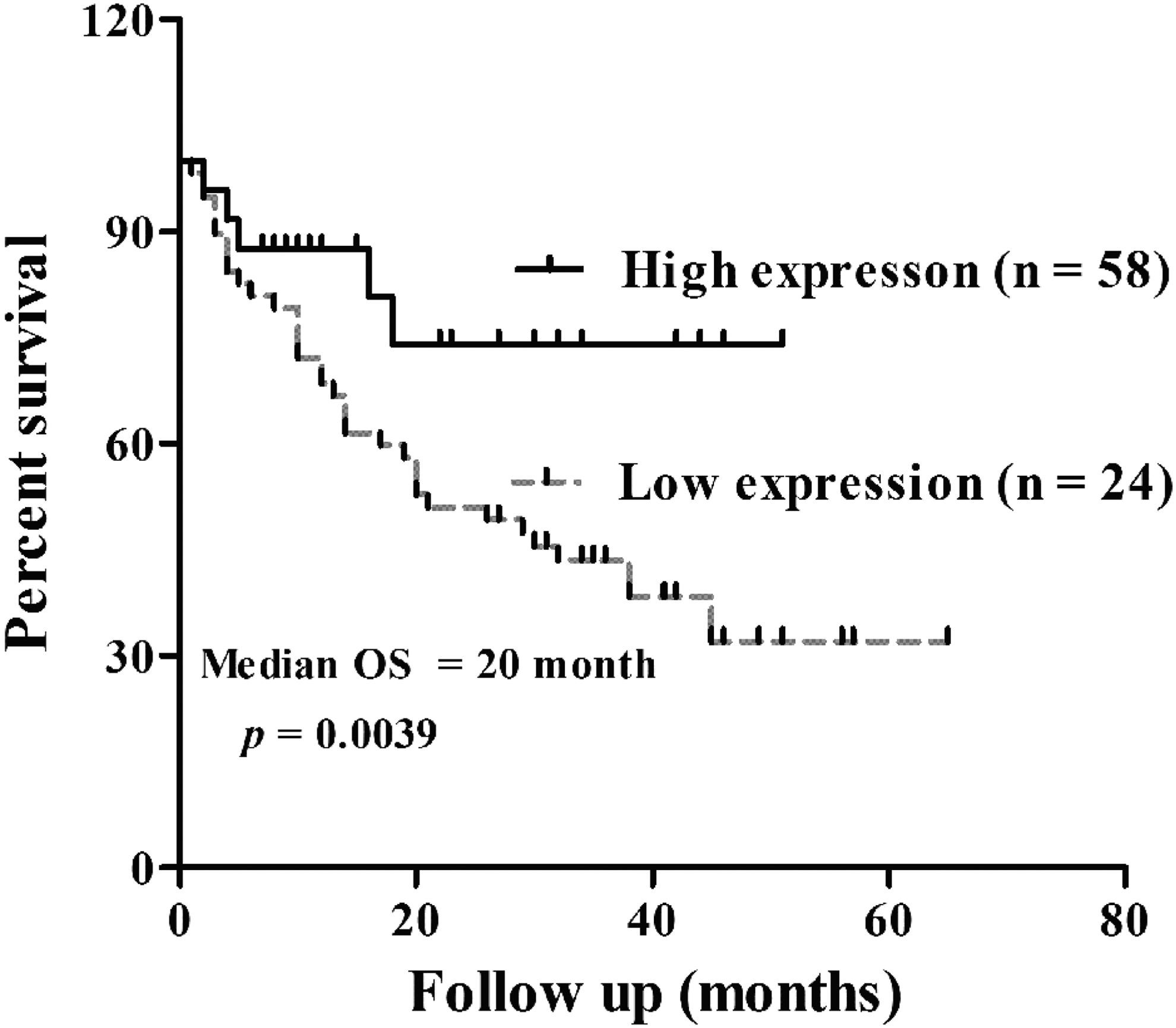
Decreased expression of miR-516b predicted poor prognosis in patients with NSCLC. Overall survival curves for two groups defined by low and high expression of miR-516b in patients with NSCLC (p = 0.0039, log-rank test). NSCLC, nonsmall cell lung cancer.
To determine whether miR-516b could be used as an independent risk factor for poor prognosis of NSCLC patients, miR-516b expression levels and clinicopathological factors were further analyzed by Cox's univariate and multivariate hazard regression model. As shown in Table 3, tumor size (hazard ratio [HR] = 1.234, 95% confidence interval [CI]: 0.687–1.756; p = 0.006), TNM stage (HR = 1.310, 95% CI: 0.968–2.635; p = 0.034), distant metastasis (HR = 1.246, 95% CI:1.347–3.241; p = 0.056), and tissue miR-516b expression level (HR = 2.435, 95% CI: 1.423–2.457; p = 0.003) were significantly correlated with overall survival in univariate analysis. Furthermore, multivariate analysis revealed that miR-516b expression, tumor size, and TNM stage were independent prognostic factors for overall survival of NSCLC patients (p = 0.007, p = 0.013, p = 0.032, respectively). Therefore, Cox regression analysis suggested that miR-516b expression has a significant correlation with poor prognosis of NSCLC patients as an independent factor.
Bold values indicate significant differences.
CI, confidence interval; HR, hazard ratio.
Overexpression of miR-516b suppressed cell viability and proliferation in A549 cells
To further assess the biological role of miR-516b in NSCLC cells, miR-516b mimics or NC mimics were transfected into A549 cells. As shown in Figure 3A, the expression level of miR-516b was significantly upregulated in miR-516b mimics transfected cells compared with NC transfected cells (p < 0.001), as determined by qRT-PCR analysis. CCK-8 assay indicated that overexpression of miR-516b remarkably decreased optical density (OD) value from 3.545 ± 0.087 to 1.834 ± 0.015 on day 4. Obviously, the OD value was reduced from 4.412 ± 0.143 in NC mimics group to 2.403 ± 0.140 in miR-516b mimics group on day 5 (Fig. 3B, p < 0.001), which indicated cell viability was significantly impaired after miR-516b overexpression. In addition, colony formation assay showed smaller and fewer colonies formed in miR-516b mimics transfected cells compared with NC transfected cells or controls (65 ± 2 vs. 128 ± 4 or 127 ± 3, Fig. 3C, p < 0.001). Collectively, these results demonstrated that miR-516b might play a suppressive role in NSCLC cell proliferation in vitro.

Overexpression of miR-516b inhibits A549 cell viability and proliferation in NSCLC.
Overexpression of miR-516b affected cell cycle progression and apoptosis in A549 cells
To determine whether proliferation inhibition by miR-516b has an effect on cell cycle distribution, DNA content was analyzed by flow cytometry in cells transfected with miR-516b mimics. As shown in Figure 4A, the percentage of cells in G0/G1 phase was significantly increased from 52.14% ± 0.47% in the NC mimics group to 65.90% ± 0.31% in the miR-516b mimics group, whereas cells in G2/M phase were obviously decreased from 19.96% ± 0.91% in the NC mimics group to 7.91% ± 0.32% in the miR-516b mimics group in A549 cells (p < 0.01). Furthermore, it was also observed that more cells were Annexin V+/7-AAD− and Annexin V+/7-AAD+, which represent early apoptosis and late apoptosis, respectively, after miR-516b mimics transfection (Fig. 4B, p < 0.001). These results indicated that miR-516b inhibited cell cycle progression at G0/G1 phase and promoted early and late cell apoptosis in NSCLC cells.
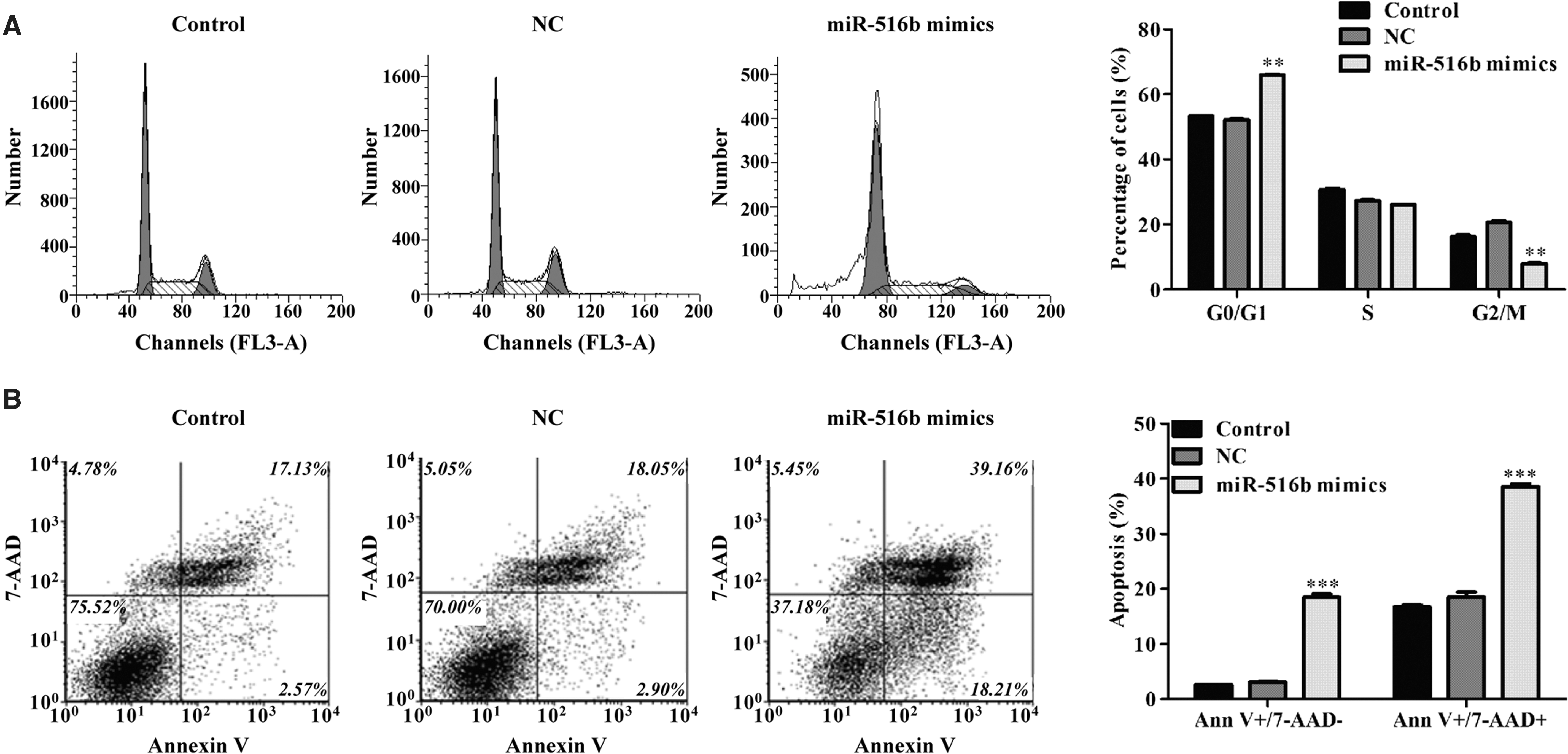
Overexpression of miR-516b induced cell cycle arrest and apoptosis in NSCLC.
Overexpression of miR-516b inhibits cell migration and invasion in A549 cells
To evaluate whether miR-51b affected the metastasis of NSCLC cells, Transwell migration and invasion assays were performed in A549 cells after being transfected with miR-516b mimics or NC mimics. As shown in Figure 5A, Transwell migration assay showed a significant reduction of cell migration in miR-516b mimics-transfected cells compared with the NC mimics or control (45 ± 4 vs. 107 ± 5 or 110 ± 3, p < 0.05). Moreover, Matrigel invasion assay also confirmed the significantly inhibitory effect of miR-516b mimics on A549 cell invasion (Fig. 5B, p < 0.05).
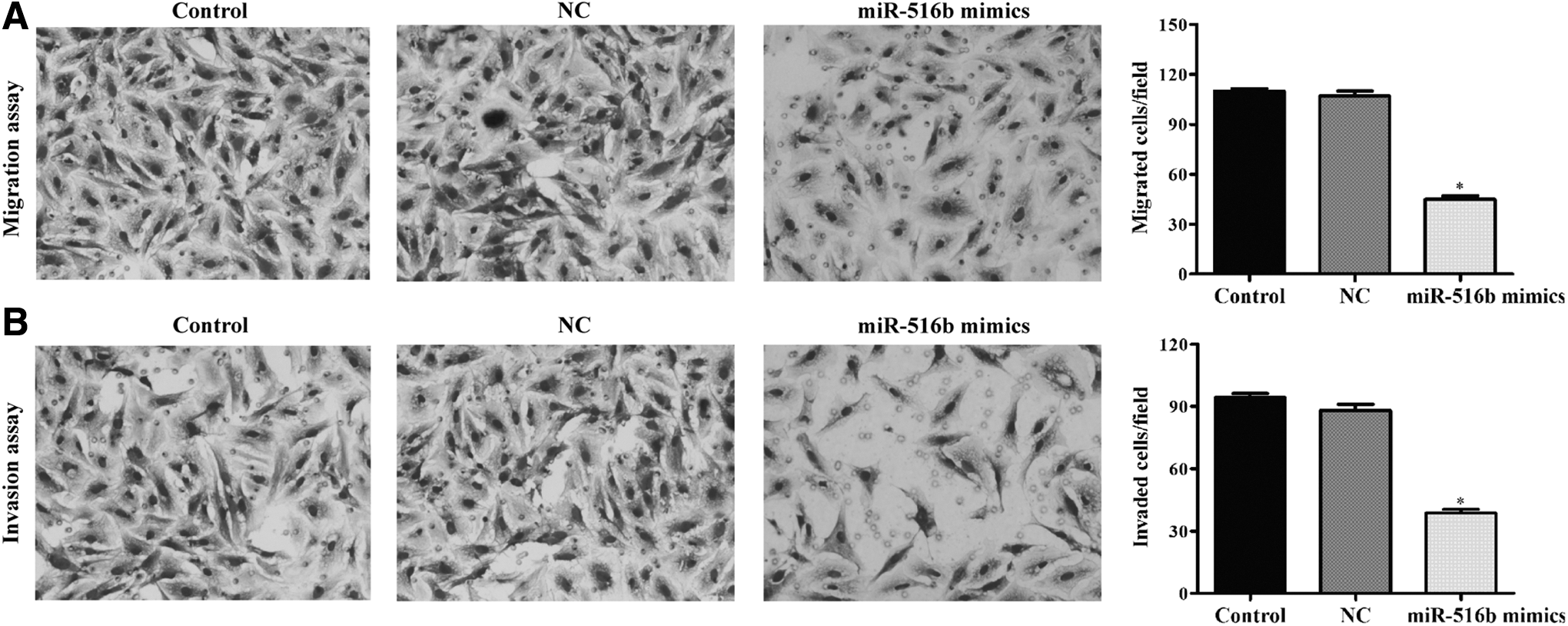
Overexpression of miR-516b suppressed cell migration and invasion ability in NSCLC.
Overexpression of miR-516b regulated markers associated with cell cycle, apoptotic, and migration
Furthermore, the expression alterations of some regulators and markers were detected. As shown in Figure 6A, the expression level of CDK2 mRNA, associated with G1-S transition, was decreased in the miR-516b mimics group. BAX, as a proapoptotic marker, was significantly increased after miR-516b overexpression. Besides, the expression levels of MMP-2 and MMP-9 mRNA were both reduced in A549 cells following miR-516b overexpression. Consistently, Western blotting demonstrated that overexpression of miR-516b downregulated the protein expression of MMP-2, MMP-9, and CDK2, whereas upregulated BAX expression in A549 cells (Fig. 6B).
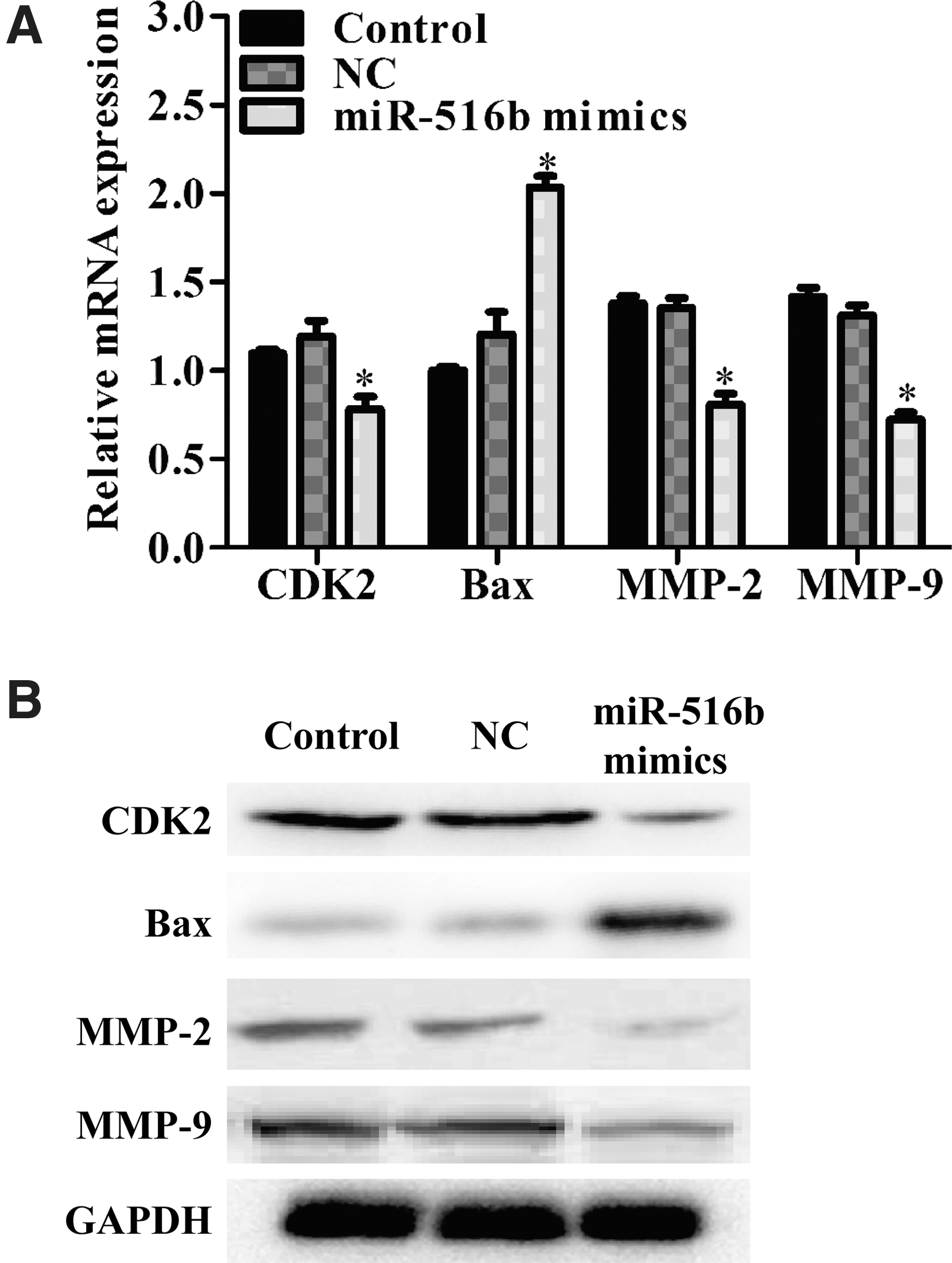
Effects of miR-516b transfection on NSCLC. miR-516b overexpression reduced CDK2, MMP-2, and MMP-9, whereas increased BAX mRNA
Discussion
NSCLC is a malignant tumor that accounts for ∼85% of lung cancers, which is mostly diagnosed in late stages, leading to poor survival. 16 The molecular and cellular mechanisms of NSCLC remain poorly understood. In this study, it was found that miR-516b was significantly downregulated in NSCLC compared with adjacent normal tissues, implying a possible involvement of miR-516b in NSCLC development. The results shown here have been supported by a previous study, which has revealed that miR-516b might be a tumor suppressor in lung cancer. 14 Consistently, the expression of miR-516b was found to be lower in aggressive tumors than in nonaggressive primary tumors in melanoma. 17 Furthermore, miR-516b has been identified as specific miR in fetal growth restriction pregnancy 18 and facioscapulohumeral muscular dystrophy. 19
In addition, decreased miR-516b expression was significantly associated with tumor size (p = 0.004), TNM stage (p = 0.016), and tissue source (p = 0.0139). Moreover, it was observed that the NSCLC patients with lower tissue miR-516b expression had shorter overall survival rate, and miR-516b expression was a significant independent prognostic factor for NSCLC patients. These findings indicated that low miR-516b could predict poor outcomes in NSCLC patients.
To know more about the biological function of miR-516b in NSCLC in vitro, the authors first found it was commonly downregulated in NSCLC cell lines compared with normal lung cells, which was consistent with the mentioned results in NSCLC tissues. Then, in vitro functional assays demonstrated that upregulation of miR-516b expression in A549 cells significantly inhibited cell proliferation, cell cycle progression, migration, and invasion, and promoted cell apoptosis. CDK2 has been reported to regulate the G1 to S phase transition during the late G1 phase. 20 Bax is a representative member of the Bcl-2 family that promotes apoptosis. 21 In this study, miR-516b could decrease the CDK2 expression CDK2 and increase the Bax expression, which suggested that upregulation of miR-516b suppressed cell cycle progression might via downregulating CDK2 and promoted apoptosis by upregulating BAX. In addition, MMP-2 and MMP-9 are two of the most studied members of the matrix metalloproteinase family, which play central roles in cell invasion and metastasis. 22,23 In line with these pieces of evidence, overexpression of miR-516b was found to downregulate both MMP-2 and MMP-9 mRNA and protein expression, further demonstrating that upregulation of miR-516b may inhibit invasion and metastasis in NSCLC.
These results suggest that miR-516b might serve as a useful diagnostic and prognostic biomarker for NSCLC. Notably, no significant difference was observed between miR-516b expression and LN metastasis, but cellular experiments indicated that miR-516b plays a suppressive role in cell migration and invasion in A549 cells, which might be ascribed to small sample sizes. To the authors' best knowledge, this is the first report regarding the clinical significance and biological function of miR-516b in NSCLC.
In summary, novel pieces of evidence present that miR-516b is downregulated in NSCLC patients and serve as an independent prognostic factor for the poor survival of NSCLC patients. Even though more cellular and animal model studies are needed to investigate the function of miR-516b in the development of NSCLC, the findings of this study support a novel role for miR-516b in NSCLC progression and also lay the foundation for further studies of the role of miR-516b in NSCLC.
Footnotes
Disclosure Statement
No competing financial interests exist.