
Abstract
Aim:
Considering the central role of dendritic cells (DCs) on the development of an antitumor immune response, in this study we used a murine model to evaluate how DC transfection with drug-treated tumor cell RNA changes their phenotype, and whether transfection enhances the in vivo effectiveness of a DC-based antitumor vaccine.
Materials and Methods:
MC-38 colorectal tumor cells were pretreated with the minimum effective concentration of 5-fluorouracil (5-FU), then their total RNA was extracted and transfected into DCs. These DCs were inoculated into C57Bl/6 mice bearing subcutaneous MC-38 tumor.
Results:
DC transfection with drug-treated tumor RNA increases the percentages of CD40+ (from 37.6% to 61.4%), CD86+ (from 39.8% to 53.4%), and major histocompatibility complex class II+ (from 51.2% to 75.3%) cells, whereas significantly increases the in vivo generation of interferon-γ producer lymphocytes.
Conclusion:
These results reinforce our view that treatment of tumor cells with 5-FU induces transcriptional changes that can be transferred to DCs by RNA transfection, enhancing their ability to stimulate an antitumor response.
Introduction
Colorectal cancer (CRC) ranks in the third place among the most frequent cancer types worldwide and is the second most common cause of mortality among cancer patients in the United States. 1,2 Malignant transformation of intestinal epithelial cells is attributed to genetic factors or exposure to food and environmental oncogenic compounds, 3,4 whereas previous inflammatory diseases such as Crohn's disease and colitis can enhance the development of CRC. 5
Both local and systemic immune responses are triggered to delay tumor progression, being that cytotoxic T cells specifically recognize and kill tumor cells in an major histocompatibility complex (MHC)-restricted manner. 6,7 Development of specific antitumor response is strongly dependent on dendritic cells (DCs) that represent the main antigen presenting cells. Once processing tumor antigens DCs are able to stimulate Th1 lymphocytes, which drive the immune response to the generation of tumor-specific cytotoxic T lymphocytes (CTL). However, in most CRC patients DCs are suppressed or remain poorly differentiated at the tumor site, whose density is three times lower than in normal colonic mucosa. In fact, DCs isolated from tumor mass show low density of activation markers or co-stimulatory molecules, especially when these cells are found in metastatic tumors. 8 In contrast, Schwaab et al. 8 observed that tumors with a high proportion of infiltrating T cells expressing tumor necrosis factor have a higher number of mature DCs, indicating the role of this cytokine in maturation and the antigen presentation function. The DC:CD4 and DC:CD8 interactions at tumor borders are related to a better prognosis and reduced metastasis in CRC patients, reinforcing the view of a central role of DCs in the antitumor immunity. 8,9 Therefore, therapies for CRC patients should be conducted to restore the ability of the immune system to recognize and specifically react against tumor antigens.
Proposals of DC-based therapeutic antitumor vaccines (DC-vax) have been used to achieve this goal. 10 –12 One of the approaches for generating DC-vax is based on transfection of tumor RNA into DCs, as previously reported by Nencioni et al. 13 and Boczkowski et al., 14 who achieved a protective immunity against CRC in a murine model. Our previous studies have shown that pretreatment of tumor cells with low concentrations of paclitaxel (PAC) increases their immunogenicity, thus facilitating the generation of specific cytotoxic T lymphocytes (CTL). 15 In addition, we observed that DCs are modulated in vitro by exposure to ultralow concentrations of antineoplastic agents 16 and that their sensitization with tumor lysate is enhanced by previous incubation with a low concentration of PAC. 16 Thus, although drugs at such concentrations cannot induce apoptosis or necrosis of tumor cells, they are able to modulate the cytokine production in the tumor microenvironment and stimulate the in vitro DC maturation and functions. 16,17 Clinically, some centers use the metronomic chemotherapy schedule, based on the more frequent administration of moderate doses of chemotherapeutic agents. This schedule contrasts with the conventional maximal tolerable dose, usually followed by severe side-effects, including mielosuppression and impaired immune response. 18
Since 5-fluorouracil (5-FU) is the main antineoplastic agent for colon cancer, this project was designed to evaluate whether DC transfection with total RNA of tumor cells, pretreated with the minimum effective dose of 5-FU, is able to prime them for an antitumor response. Our studies showed that transfection increases the CD40, CD86, and MHC class II expression on DCs, and those animals treated with these cells in vitro produced higher levels of interferon (IFN)-γ.
Materials and Methods
Experimental design
Immature DCs were differentiated from bone marrow precursor cells and transfected with total RNA of tumor cells exposed to the minimum effective concentration of 5-FU (DC-FU). These cells were intradermally inoculated in mice bearing MC-38 colon cancer cells (at days 10 and 17). Spleen and tumor mass were excised on the 30th day after tumor implantation for histopathological analysis (tumor mass) and evaluation of specific antitumor responsiveness (spleen lymphocytes).
Animals
C57Bl/6 mice were obtained from the Animal Facility of the University of São Paulo, Ribeirão Preto, Brazil. They were separated into four groups (N = 4) and maintained under controlled temperature and humidity, water and food supply ad libitum, and 12/12-hour light/dark periods at the Department of Pathology, Faculty of Medicine at Botucatu–UNESP. All the proceedings followed the rules of Brazilian College of Animal Experimentation and were approved by the local Ethics Committee (Protocol No. 194-CEEA).
Maintenance of MC-38 cells and treatment with 5-fluorouracil
Murine colorectal MC-38 tumor cells were cultured in complete culture medium (RPMI 1640 supplemented with 10% fetal bovine serum, 2 mM
Tumor RNA extraction
Total RNA was extracted from 4 × 106 MC-38 cells with TRIZOL and isolated in RNeasy TM Maxi anion exchange spin columns (Qiagen) according to the manufacturer instructions. RNA purity and concentration were checked by spectrophotometry under 260/280 nm wavelength (Nanodrop Technologies, Inc., Wilmington, DE).
Generation of myeloid DCs
Bone marrow was collected from the tibiae and femurs of healthy mice for the generation of myeloid DCs. Erythrocytes were lysed with a hemolytic solution, and T and B lymphocytes were then depleted with anti-CD4, -CD8, and -B220 antibodies plus complement. Cells were cultured overnight in complete culture medium on polystyrene plates and nonadherent cells were collected 18 hours later. These cells were seeded in six-well culture plates with 80 ng/mL of recombinant murine granulocyte-macrophage colony-stimulating factor and interleukin (IL)-4 at 37°C in a 5% CO2 chamber. 17 Six days later, immature DCs were collected, washed with fresh RPMI, suspended in the serum-free AIM-V medium, and adjusted to 2.4 × 106/mL. These DCs were transfected with tumor total RNA (5 μg/106cells) by lipotransfection using the TransMessenger Kit (Qiagen) following the manufacturer's instructions. After 24 hours, DC preparations were analyzed by flow cytometry (FACS Calibur) for the markers CD11a, F4/80, MHC class II molecules, CD80, CD86, and CD40 (BioLegend Antibodies).
Subcutaneous development of MC-38 colon cancer cells and treatment with DCs
Mice were subcutaneously inoculated with 4.5 × 105 tumor cells. Ten days later, when tumor site became visible, animals were inoculated with 106 DCs transfected with drug-treated tumor RNA (group DC-FU). As controls, tumor-bearing mice were inoculated with nontransfected DCs (group DC), DCs transfected with RNA of wild-type untreated tumor cells (group WT), or just PBS. Treatment was repeated at the 17th day and tumor size (area) was measured weekly until the 30th day.
In vitro lymphocyte challenge with tumor cells
To investigate the generation of specific antitumor T lymphocytes by tumor-bearing mice under different treatments, we evaluated the ability of their spleen cells to produce IFN-γ after contact with tumor target cells. Thus, 2.5 × 105 spleen cells of animals subjected to different treatments were put onto a monolayer of MC-38 cells (5 × 104) and cocultured for 24 hours, when supernatants were collected and frozen for further analysis. 19 Levels of both IFN-γ and IL-10 were measured by ELISA MAX Deluxe Set (BioLegend), according to the manufacturer's instructions.
Histological analysis
Tumor tissue fragments were fixed with 10% formalin for 24 hours and included in paraffin. Histological 5μm slides were routinely processed and dyed with hematoxylin-eosin for histopathological analysis.
Statistical analysis
Groups were tested for homogeneity of variances (Bartlett test) and analyzed by the ANOVA statistical test using the statistical software SigmaStat. Differences among the groups were considered significant when p < 0.05.
Results
RNA transfection activates DCs
We first observed that gated cell population has a low percentage of CD14+ cells, indicating the differentiation of myeloid cells into DCs. DCs transfected with RNA of 5-FU-treated tumor cells (group DC-FU) showed higher percentage of CD40+ (increased from 34.3% to 54.6%), CD86+ (from 38.5% to 59.3%), and MHC class II+ (from 51.6% to 81.5%) than DC control group as revealed by flow cytometry (Fig. 1). This positive effect shows that exposure of tumor cells to 5-FU, before transfection of their RNA into DCs, increases the expression of these markers, whereas RNA of WT tumor cells did not induce the same level of stimulation (CD40+ 49.5%, CD86+ 53%, MHC class II+ 77.9%) even though the percentage of cells expressing these markers was also higher than that observed in untreated DCs.
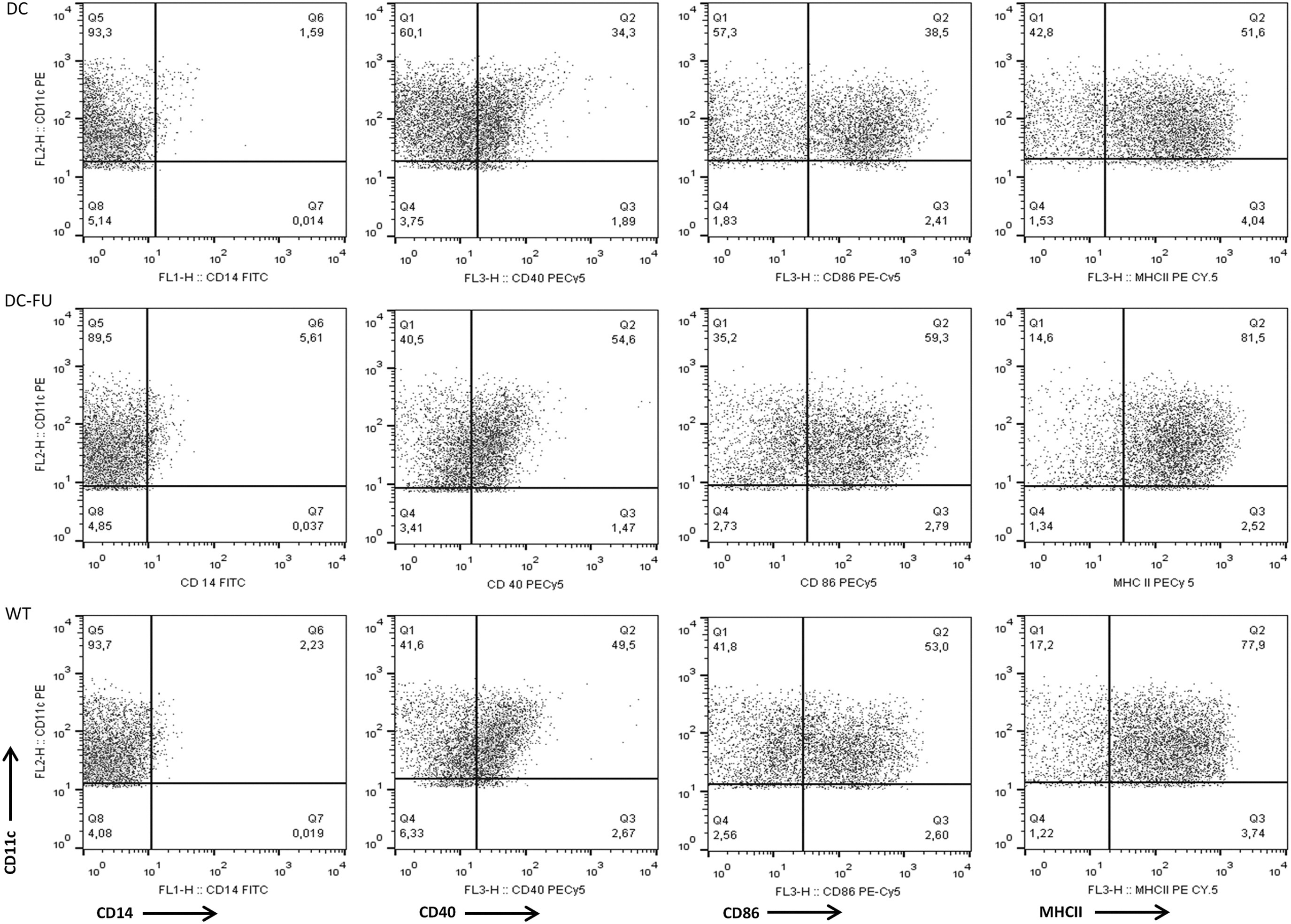
Phenotype of transfected (DC-FU) and nontransfected DCs at the 6th day of culture. Dot plots show double labeling with CD11c and CD40, CD86 and MHC class II of nontransfected control DCs in the upper side, and DCs transfected with RNA of 5-FU-treated tumor cells in the down side. Results are expressed as percentage of positive cells. 5-FU, 5-fluorouracil; DC, dendritic cell.
DC transfection with tumor improves the in vivo generation of specific antitumor lymphocytes
Since tumor-specific lymphocytes are able to produce IFN-γ after a challenge with tumor cells, spleen cells of tumor-bearing mice were in vitro challenged with MC-38 cells and culture supernatants were collected for IFN-γ quantification. We observed that animals inoculated with DC-FU generated a higher number of specific anti-MC-38 T lymphocytes than controls, as evidenced by IFN-γ production (Fig 2A). No relevant changes were observed in the IL-10 production by spleen cells (Fig. 2B).
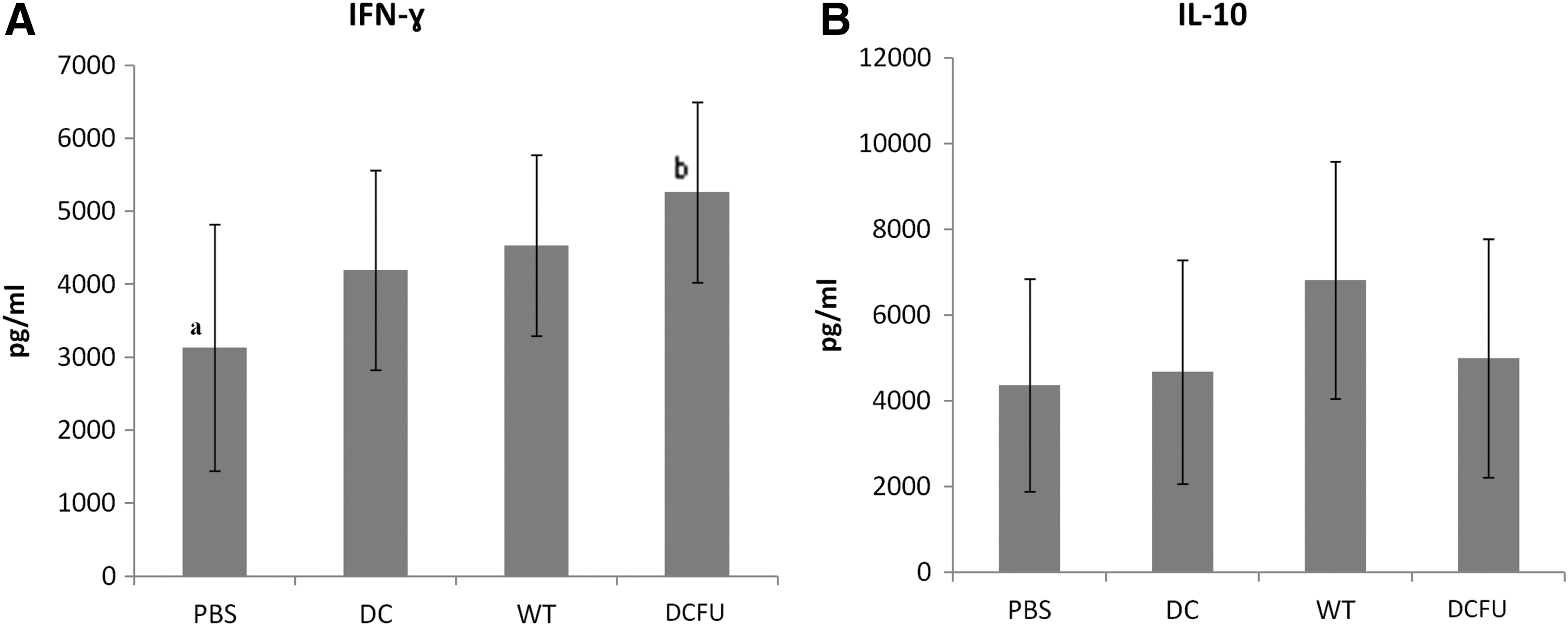
In vitro IFN-γ
DCs inhibit in vivo tumor growth
In vivo effectivity of DCs was evaluated by measuring the tumor growth in each animal once a week for 30 days and indicated that it was significantly decreased by administration of DCs, independently of their transfection with tumor RNA. In fact, tumor growth in the groups WT and DC-FU was lower than in PBS control, but there was no difference among the experimental groups (Fig. 3).
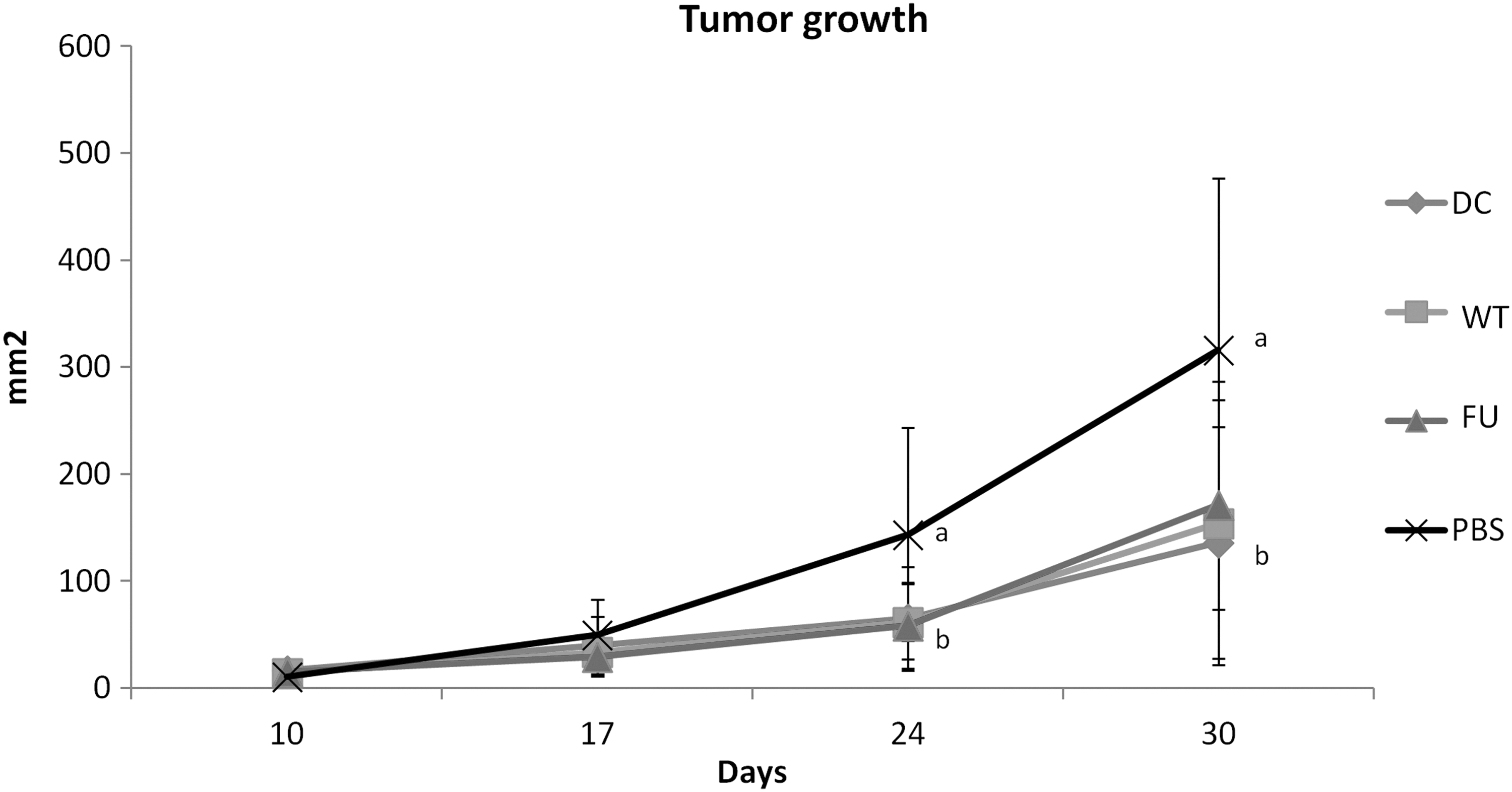
Effect of DC vaccine on tumor growth. Tumor cells were subcutaneously inoculated and animals were treated with DCs transfected with RNA of WT tumor cells, or of 5-FU-treated tumor cells (FU). As control, we inoculated nontransfected DCs (DC) or just PBS. Tumor area was measured once a week for 30 days. Mean ± SD of two experiments performed independently. Statistical analysis by ANOVA and Tukey–Kramer multiple comparisons test, a ≠ b; p < 0.05; N = 8. PBS, phosphate-buffered saline.
Histopathological analysis showed extensive areas of necrosis, ulceration, and diffuse inflammatory polymorphonuclear infiltration in tumor tissue of DCs and DC-FU groups. No significant differences among treatments were observed (Fig. 4).

Histopathological analysis of MC-38 tumor on mice showing absence of significant differences among groups. Edema, apoptotic bodies, and necrotic areas were observed in tumor masses obtained from all groups of mice treated with different preparations of DCs. Color images are available online at
Discussion
We previously observed that low concentration of PAC is able to increase the immunogenicity of colon cancer cells. 15 We also observed that ultralow nontoxic concentrations of selected chemotherapeutics are able to modulate the immune system 17 and increase the maturation and functions of human DCs. 16,17 In accordance with these findings, in this study, we first observed that DCs transfected with RNA of MC-38 cells pretreated with the minimum effective concentration of 5-FU increase the number of cells expressing CD40, CD86, and MHC class II. Upregulation of CD86, MHC class II, as well as CD80 and CD83 was also observed in our previous study with human DCs transfected with RNA of 5-FU-treated HCT-116 colon cancer cells. 19 These markers on the cell surface are necessary to activate and initiate immune response of T cells at the draining lymph nodes. 20 –22 In accordance with this, we observed that modulation of those markers enhanced the ability of DCs to induce both the in vitro allogeneic responsiveness (mixed leukocyte reaction) and the generation of antitumor CTL. 19 In another previous study, we observed that treatment of tumor cells with low concentrations of the antineoplastic drugs PAC and doxorubicin induces the superexpression of several tumor antigens, antigen-processing machinery components, and heat shock proteins. 15 So, we think that if these products are induced in DCs after RNA transfection, they could work as danger signals to induce increased expression of costimulatory molecules.
In addition, spleen cells from animals treated with DC-FU showed higher ability to produce IFN-γ than control, a cytokine that plays a key role in promoting cell-mediated antitumor response. 23,24 Since this cytokine was produced when spleen cells were put in contact with MC-38 target cells, it indicates the in vivo development of specific antitumor response. IL-10 levels were similar for all groups, indicating that none of the DC preparations has regulatory property to hinder the antitumor response.
Taken together, data on phenotype and IFN production suggest that transfection of tumor RNA into DCs facilitates the maturation and activation of these cells, thus improving their antigen-presentation ability. We recently observed similar results in a parallel study with human DCs, supporting our concept that treating tumor cells with drugs before RNA extraction renders DCs more efficient to fight CRC cells. 19
Despite the in vitro effect on specific IFN-producing cells, our in vivo studies showed no difference between transfected and nontransfected DC preparations in antitumor activity. Controversial results between in vitro and in vivo activities of DC vaccine preparations were also reported by He et al., 25 who observed that lysate-pulsed DCs present a poor protective effect against the development of CT26 cells, despite their ability to stimulate CTL activity and to induce INF-γ production. Similar to our results, Kao et al. 26 demonstrated that DC vaccine is able to prevent the growth of inoculated tumor cells and induce a strong in vitro CTL response, but it does not present a therapeutic role, failing to inhibit the growth of pre-existing tumor.
It is possible that such a contrast between in vitro and in vivo findings is because in vivo, the chronic inflammation drives the response to a TH17 profile in the tumor microenvironment, making it difficult for DC to change this profile, whereas these complex interactions do not occur in vitro. In addition, production of reactive oxygen species, proangiogenic factors (e.g., vascular endothelial growth factor), and matrix metalloproteases (which facilitate invasion and metastasis) contributes to a poor host responsiveness. 27 Moreover, chronic inflammation at the tumor site usually triggers regulatory mechanisms such as infiltration of Treg, 28 tolerogenic DCs, 29 and myeloid-derived suppressor cells (MDSCs), 30 establishing a microenvironment favorable for tumor growth.
In our study, we did not investigate the phenotype of tumor infiltrating cells but we observed the presence of cells looking like polymorphonuclear cells (PMN) surrounding the necrotic area of vaccine-treated animals. The activity of PMN is classically considered a tumor-enhancing factor, especially when such cells are associated with chronic inflammatory responses. 31 In addition, there is a morphological similarity between tumor-associated neutrophils and MDSCs, being the latter involved in tumor progression both by direct immunosuppression and through the production of angiogenic factors, matrix-degrading enzymes, and growth factors. 32 Therefore, it is possible that part of infiltrating cells we have found in the tumor microenvironment are MDSCs that produce IL-10 and TGF-β. We consider that despite the generation of specific IFN-γ-producing anti-MC-38 cells in the spleen, secretion of IL-10 and TGF-β by these putative MDSCs in the tumor site could locally modulate TH1 responsiveness and avoid the in vivo immunostimulatory activity of antigen-loaded DCs. 33,34
In conclusion, our results indicate that transfection induces phenotypic and in vitro functional changes in DCs (increased CD40, CD86, and MHC class II expression, and higher ability to induce IFN-γ), but the feasibility of this approach to prepare therapeutic antitumor DC-based vaccines requires more extensive investigation, for instance associating these cells with antibodies or cytokines able to overcome the local regulation of immune response.
Footnotes
Acknowledgments
We are grateful to São Paulo Research Foundation (FAPESP) for the grant 2009/18331-8 (regular research grant). M.R.C., C.P.R., and J.C.L.F. were recipients of FAPESP scholarships (2010/06013-9, 2011/19403-2, and 2011/05258-0, respectively), and C.M.G. and R.K. received scholarships from the Brazilian Council of Research and Development (CNPq: 13991/2013-2 and 303952/2010-5). We also thank Fabiana Zambuzi and Victoria Elisabeth Galvão for technical support.
Disclosure Statement
No competing financial interests exist.