
Abstract
Background:
Osteosarcoma (OS) is a frequent primary bone malignancy. MicroRNA-497-5p (miR-497-5p) has been recently reported to be downregulated in angiosarcoma, but its potential role in OS remains unclear.
Methods:
Quantitative real-time polymerase chain reaction was used to determine miR-497-5p expression in OS tissues and cell lines. The effect of miR-497-5p on cell growth, cell cycle progression and apoptosis were measured using CCK-8 and flow cytometry assays, respectively. The effect of miR-497-5p on ADP-ribosylation factor-like protein 2 (ARL2) was determined by luciferase reporter assay and western blot.
Results:
We found miR-497-5p was significantly downregulated in OS tissues and cells. Overexpression of miR-497-5p in OS cells inhibited tumor cell proliferation and induced a significant G0/G1 cell cycle arrest and apoptosis in vitro. Additionally, we found ARL2 was a new target of miR-497-5p. Moreover, ARL2 and P-p53 protein levels were significantly downregulated by treatment with miR-497-5p mimics in OS cells.
Conclusions:
Taken together, our findings revealed miR-497-5p may be a tumor suppressor in OS and serve as a promising therapeutic target for OS.
Introduction
Osteosarcoma (OS) is a frequent primary bone malignancy in adolescence and older adulthood. 1 Usually, OS originate from the malignant transformation of human mesenchaymal stem cells into osteoblasts. 2 The annual incidence of OS is about 3/1,000,0003 and occurs more commonly in the metaphysics of femur, tibia, and humerus for young patients. 4 It is known that many OS patients are diagnosed at later stages with local dissemination in the same extremity, most often involving lung. 5 Amputation, which was once considered as the only strategy for this disease, however, with the rapid development of therapeutic of options, such as effective preoperative neoadjuvant and postoperative adjuvant treatments have reduced the amputation rates, also improved to some extent the quality of life. 6 But, prognosis remains poor with far worth at <30% survival in OS patients with recurrence or metastatic. 7 Hence, identification of tumor-specific molecules and mechanism of action in OS will provide a better understanding of tumorigenesis and development, and may supply a critical target for improving the treatment of OS.
MicroRNAs (miRs) constitute a class of small noncoding RNAs, with nucleotide sequences of length ranging from 19 to 24 nt. 8 They function as negative regulators of gene expression with 5′-end of miR often conferring their ability to bind the 3′-UTR of the target mRNA. 9 So far, more than 700 miR genes exist in the human genome, and about more than 30% of human genes are controlled by miRNAs. 10 miRs have been shown to participate in a variety of biological and pathologic events, such as cell cycle, cell death, differentiation, invasion, proliferation, stress responses, and others. 11 Accumulating evidence shows that deregulation of specific miRs associates with the occurrence and development of tumor including chronic lymphocytic leukemia, 12 breast cancer, 13 lung cancer, 14 ovarian cancer, 15 and OS. 16 miRs are thought to play important roles in tumor suppression or carcinogenesis, 17 and also impacts on clinical outcome in patients. 7 To date, the biological function of several miRs has been identified in OS. For example, miR-34 family involving in the OS oncogenesis though regulation of Notch signaling pathway. 16 Sarver et al. 18 reported that downregulation of miRNA clusters at the 14q32 locus promotes the biological behavior of OS, and the representative of 14q32 miRNAs miR-382, 134, and 544 have prognostic and predictive significance in OS. miR-143 has been involved in OS lung metastasis via affecting the expression of matrix metalloprotease-13. 7 MicroRNA-497-5p (miR-497-5p), belongs to the miR497–195 cluster of miR-15 family, exhibits the highest similarity to miR-15a and miR-15b. 19 It has been proposed that miR497–195 cluster exert regulatory function in differentiation of osteoblast though disturbance of BMP signaling. 19 Previous study has shown that miR-15a and miR-16-1 function to induce apoptotic cell death and cell cycle arrest in OS. 20 miR-15b was reported to regulate multidrug resistance in OS. 21 Hence, miR-497-5p may also implicate in OS, but has never been identified.
ADP-ribosylation factor-like protein 2 (ARL2) is a family member of monomeric G proteins that involved in the folding of tubulin peptides and actually shown to cooperate with miR-214 to effect colon cancer 22 and cervical cancer carcinogenesis. 23 Previous study showed that ARL2 is a target of miR-15b, and upregulation of miR-15b decreased transcription and translation of ARL2, and thus suppresses ADP/ATP exchange and ATP synthesis. 24 However, whether there is a relationship between miR-497-5p and ARL2 in OS is unknown.
In this study, we undertook quantitative real-time polymerase chain reaction (qRT-PCR) to determine miR-497-5p expression pattern in OS tissues and their adjacent normal tissues, and among normal osteoblastic cell line and five OS cell lines. Then, miR-497-5p mimics were applied to upregulate miR-497-5p expression in MG-63 and U2OS cells. CCK-8 and flow cytometric assays were used to explore the biological action of miR-497-5p in OS. Finally, dual luciferase reporter assay was performed to verify whether ARL2 is a direct target of miR-497-5p.
Materials and Methods
Tissue sample collection
A total of 30 pairs of fresh OS and adjacent noncancerous tissues (located >3 cm from the tumor) were collected from routine therapeutic surgeries at our department between 2014 and 2016. Before surgical resection, no patient received radiotherapy or chemotherapy. Collected samples were immediately stored in liquid nitrogen at −80°C until use. All patients had given written informed consent and this study was approved by the Ethics Committee of People's Hospital affiliated to Hubei University of Medicine.
Cell lines and transfection
Human OS cell lines, MG-63, U2OS, SAOS-2, SOSP-9607, G292, and a noncancer osteoblastic cell line (hFOB) were all purchased from American Type Culture Collection (ATCC, MD). Cells were cultured in Dulbecco's modified Eagle medium (DMEM; Invitrogen, Carlsbad, CA) and supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY). All cells were maintained in a humidified atmosphere containing 5% CO2 at 37°C.
The miR-497-5p mimics, small interfering RNA for ARL2 (siARL2) and their corresponding negative control sequences (NC and siNC, respectively) were designed and synthesized by RiboBio (Guangzhou, China). For cell transfection, MG-63 and U2OS cells were seeded in six-well plates at a density of 2 × 105 cells per well in 1 mL of complete medium. To overexpress miR-497-5p, MG-63 and U2OS cells were transfected with miR-497-5p mimics For ARL2 knockdown, MG-63 and U2OS cells were transfected with siARL2 or siNC, respectively. All transfection was performed using Lipofectamine 2000 (Invitrogen) according to the manufacturer's recommendations. After 48 hours transfection, the expression levels of miR-497-5p or ARL2 were determined and a series of functional assays were performed in OS cell lines.
Quantitative real-time PCR
Total RNA was extracted from tissues or cells with TRIzol reagent (Life Technologies, NY) according to the manufacturer's instructions. Then, 10 ng of total RNA was converted into complementary DNA using miR reverse transcription kit (Life Technologies). The PCR amplification for miR-497-5p was performed by using an ABI PRISM 7700 real-time PCR system (Applied Biosystems, MA) following the manufacturer's information. Quantitative PCR was conducted at 95°C for 10 minutes followed by 40 cycles of 95°C for 15 seconds and 60°C for 60 seconds. The PCR primers for miR-497-5p were 5′-CCTTCAGCAGCACACTGTGG-3′ (forward) and 5′-CAGTGCAGGGTCCGAGGTAT-3′ (reverse) and those for U6 were 5′-CTCGCTTCGGCAGCACA-3′ (forward) and 5′-AACGCTTCACGAATTTGCGT-3′ (reverse). All reactions were run in triplicate, and miR-497-5p expression was normalized to U6 small nuclear RNA using the 2−ΔΔCT method.
Cell proliferation assay
Cell proliferation was determined using CCK-8 Kit (Dojindo Laboratories, Japan) according to the instructions from the manufacturer. Briefly, transfected cells were seeded in the 96-well at 1 × 104/well and cultured for 24 hours. Then, each well was added CCK-8 reagent and incubated for 2 hours. The optical density at the wavelength of 450 nm was acquired by a microplate reader (168–1000 Model 680; Bio-Rad).
Flow cytometric analysis of cell cycle and apoptosis
MG-63 and U2OS cells were collected and washed twice with PBS, and then underwent cell cycle and apoptotic analysis using flow cytometry. For cell cycle analysis, cells were fixed in 70% ethanol overnight and resuspended in 500 μL PBS containing 50 mg/mL RNase (Sigma-Aldrich). After incubation for 30 minutes at 37°C, staining for DNA content was performed with 50 mg/mL propidium iodide in the dark. Finally, cell cycle distribution was analyzed using fluorescence-activated cell-sorting (FACS) Flow Cytometer with ModFit LT Software (BD Biosciences, San Jose, CA). For cell apoptotic analysis, cells were stained with an Annexin V APC/7-AAD Apoptosis Detetion Kit I (BestBio, Shanghai, China) for 30 minutes at room temperature in the dark. Stained cells were analyzed using FACScan Flow Cytometer (BD Biosciences) with Cell Quest software. Tests were repeated in triplicate.
Dual luciferase reporter assay
For luciferase assay, the 3′-UTR of ARL2 containing the miR-497-5p binding sites and its corresponding mutated sequence were cloned into pRL3 luciferase promoter vector (Promega, Madison, WI), which were named as wild-type (WT) or mutant (MUT) constructs, respectively. Then, cells were seeded in a 24-well plate and cotransfected with miR-497-5p mimics or NC and the vectors carrying WT 3′-UTR or ARL2 MUT using the transfection reagent (Lipofectamine 2000; Invitrogen). Luciferase activity was measured after 24 hours using a Dual-Luciferase Reporter Assay System (Promega). Data were expressed as the ratio of Renilla luciferase to firefly luciferase activities.
Western blot analysis
Total proteins were extracted from cell lines using a RIPA lysis buffer. After quantification by BCA protein assay kit (Beyotime, China), protein was separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto polyvinylidene fluoride membranes (Millipore). After blocking with nonfat milk, the membrane was washed three times with 0.1% Tween-20 (TBST) and then incubated with antibodies against ARL2 (1:200; Abcam), P-p53 (1:500; SAB) or GAPDH (1:5000; Santa Cruz) followed by the HRP-conjugated secondary antibody. The chemiluminescent signals were visualized by ECL reagent (Applygen, Beijing).
Statistical analysis
All experiments were performed at least three times and all quantitative data are presented as mean ± standard deviation. Statistical analysis was performed using SPSS software version 16.0. Statistical significance was considered at p < 0.05 using the Student's t-test.
Results
miR-497-5p was downregulated in OS tissues and cell lines
First, we determined the expression of miR-497-5p in OS tissues and cell lines by qRT-PCR. As shown in Figure 1A, the expression of miR-497-5p mRNA was significantly downregulated in OS tissues compared with the adjacent noncancerous tissues (p < 0.001). For in vitro analysis, we also compared miR-497-5p expression in several OS cell lines (MG-63, U2OS, SAOS-2, SOSP-9607, and G292) and normal osteoblastic cell line (hFOB). Consistently, we found miR-497-5p expressed at lower levels in the above OS cell lines compared with that in hFOB (Fig. 1B). Moreover, MG-63 and U2OS cells had the lowest expression level of miR-497-5p, which thus were chosen for further studies.

Downregulation of miR-497-5p in human OS tissues and cell lines.
miR-497-5p overexpression inhibited OS cell proliferation in vitro
To explore the role of miR-497-5p in OS, MG-63 and U2OS cells were stably transfected with miR-497-5p mimics to increase its expression. After transfection, we found the expression of miR-497-5p was significantly upregulated in both MG-63 and U2OS cells compared with NC groups (Fig. 2A, p < 0.001), which indicated that overexpressed miR-497-5p cell models have been successfully constructed. Subsequently, CCK-8 assay revealed that cells overexpressing miR-497-5p showed a decreased growth rate and viability at the time points of 72 and 96 hours (Fig. 2B, p < 0.05, p < 0.01), which suggested that upregulation of miR-497-5p suppressed OS cell proliferation.
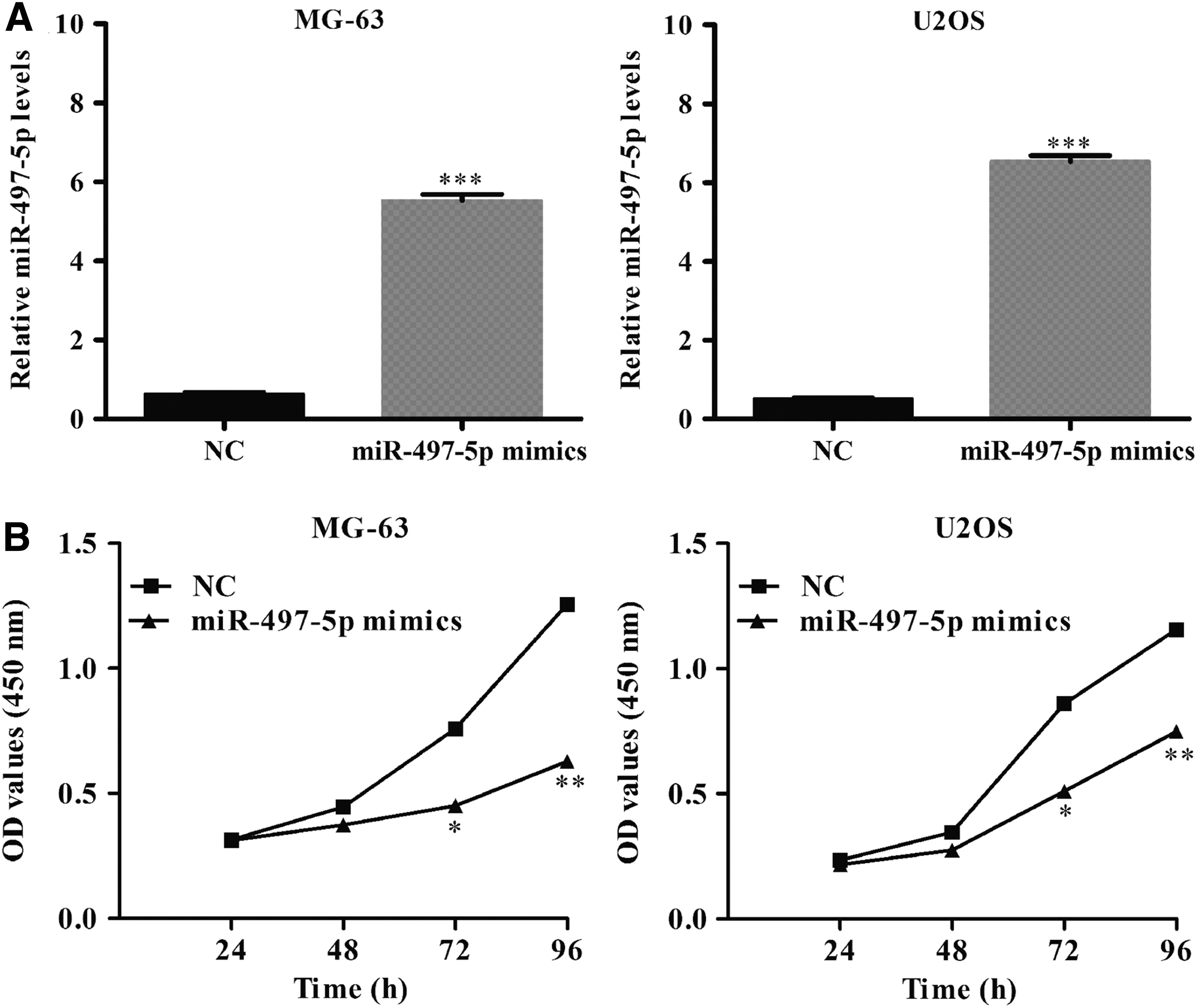
Upregulation of miR-497-5p suppresses OS cell proliferation in vitro.
miR-497-5p overexpression induced cell cycle arrest and apoptosis
We then examined the cell cycle distribution and apoptosis in OS cells after transfected with miR-497-5p mimics. As shown in Figure 3A, B, the percentage of cells in G0/G1 phase was significantly increased from 56.31% ± 0.22% in NC group to 65.80% ± 0.11% in miR-497-5p mimics (p < 0.001), while cells in S (p < 0.01) and G2/M phase (p < 0.05) was obviously decreased in MG-63 cells after transfection. Similarly, we found miR-497-5p mimics transfection also increased the percentage of cells in G0/G1 phase (p < 0.001), while decreased cells in G2/M phase (p < 0.001) in U2OS cells compared with NC groups. These data demonstrated miR-497-5p may suppress cell proliferation through blocking the G1 to S phase transition, means G0/G1 phase arrest. Furthermore, cell apoptotic assay (Fig. 3C, D, p < 0.01, p < 0.001) further indicated that miR-497-5p overexpression promoted cell apoptosis, including early apoptosis (Annexin V+/7-AAD−) and late apoptosis (Annexin V+/7-AAD+), which might be another cause of suppressive cell proliferation by miR-497-5p mimics transfection.

Upregulation of miR-497-5p induced cell cycle arrest and apoptosis in OS cells. MG63 and U2OS cells were transfected with miR-497-5p mimics or NC, respectively. Cell cycle distribution was examined by propidium iodide staining using flow cytometry
miR-497-5p directly targets ARL2 in OS cells
To understand the underlining mechanisms of miR-497-5p in OS, we used bioinformatical approach to identify the potential target of miR-497-5p. As shown in Figure 4A, ARL2 contains a putative miR-497-5p binding site in the 3′-UTR of its mRNA by using Targetscan database prediction. We then constructed 3′-UTR sequences containing WT or MUT of miR-497-5p into the pRL3 luciferase promoter vector and cotransfected with the miR-497-5p mimics or NC into MG-63 and U2OS cells. As expected, miR-409-3p significantly decreased the luciferase activity, while luciferase activity was not significantly changed in the MUT binding site (Fig. 4B, p < 0.001). In addition, transfection of miR-497-5p mimics into MG-63 and U2OS cells caused a significantly decreased level of ARL2 expression (Fig. 4C). Interestingly, we found the expression of P-p53 was remarkably upregulated in MG-63 and U2OS cells after transfection of miR-497-5p mimics (Fig. 4D). These data demonstrated that ARL2 is directly targeted by miR-497-5p in OS cells.
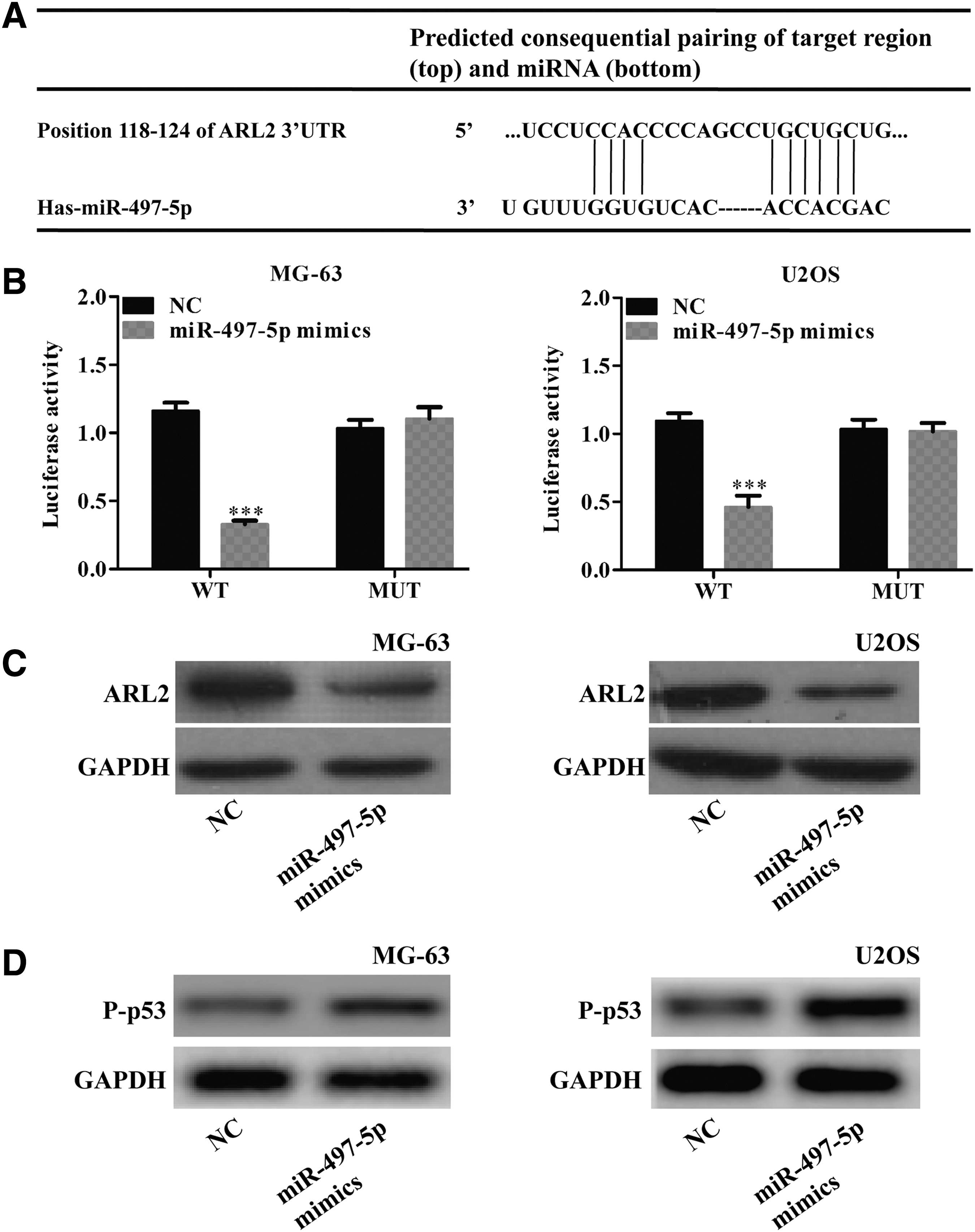
Identification of ARL2 as a target of miR-497-5p in OS cells.
Knockdown of ARL2 inhibited cell proliferation and promoted apoptosis in OS cells
Next, we performed loss-of-function assays to investigate the functional role of ARL2 in OS cells. As shown in Figure 5A, the expression of ARL2 protein was remarkably downregulated in MG-63 and U2OS cells after transfection with siARL2. In functional assays, CCK-8 showed that knockdown of ARL2 significantly suppressed cell proliferation in both MG-63 and U2OS cells (Fig. 5B, p < 0.05, p < 0.01). Moreover, the apoptosis rate of MG-63 and U2OS cells was significantly increased by transfection with siARL2 in comparison with siNC transfection (Fig. 5C, p < 0.001).

Knockdown of ARL2 suppresses cell proliferation and induced apoptosis in OS cells.
Discussion
OS is a common bone tumor with relatively high incidence occurring in young people. 25 miRs alterations are reported to regulate various oncogenes and tumor suppressor genes expression. 26 In this study, we have shown for the first time that miR-497-5p is markedly decreased in OS tissues and cells. miR-497-5p overexpression in MG-63 and U2OS cells led to an inhibition of cell proliferation, induction of G0/G1 cell cycle arrest, and apoptosis. These results indicate that miR-497-5p is a tumor suppressor in OS. Furthermore, ARL2 was identified as a direct of miR-497-5p and knockdown of ARL2 presented similar suppressive effects of miR-497-5p in OS cells. We thus suppose that miR-497-5p suppresses OS cell growth might though directly targeting ARL2.
There is a correlation between expression of specific miRNAs and ARL2 in the progression of some serious illnesses like cancer and cardiac disease. For example, miR-214 blocks growth and invasion in colon cancer 22 and cervical cancer 23 through downregulation of ARL2. Previous study showed that miR-497-5p exhibits the highest similarity to miR-15a and b. 19 miR-15b can regulate the deterioration of mitochondrial function in cardiomyocytes via targeting ARL2 3′-UTR. 24 Nishi et al. 24 also showed that ARL2 mRNA and protein expression were decreased in miR-15b-overexpressing cardiomyocytes. In this study, miR-497-5p upregulation inhibited the ARL2 3′-UTR luciferase reporter activity and downregulated protein expression of ARL2. These results suggest that the regulating and binding behaviors of miR-497-5p to ARL2 in OS are similar to miR-15b in cardiomyocytes. Interestingly, overexpression of miR-15a impedes proliferation, induces apoptosis and G0/G1 cell cycle arrest in OS SOSP-9607 cells. The same phenomenon was observed in miR-497-5p-overexpressed OS MG-63 and U2OS cells, indicating that miR-497-5p and miR-15a possesses a similar biological roles in OS, possibly due to their highly similarity. Notably, the percentage of cells in S phase was significantly decreased in MG63 cells, but has no significant change in U2OS cells, which might be the different cell subtypes and sensitivities under transfection.
ARL2 is implicated in the conformational of tubulin peptides and impact on microtubule dynamics and cell cycle progression in breast cancer cells. 27 ARL2, can influence phosphorylation of p53 at Ser15, results in microtubule sequestration of p53 in breast cancer MCF7 cells, through PP2A-mediated mechanism. 28 p53 is considered as a tumor suppressor gene that impedes development of tumorigenicity via induction of cell cycle arrest and apoptosis. 29 What's more, the expression of phosph-p53 (P-p53, Ser15) is associated with p53 apoptotic activity. 30 Both Ser15 mutation and Ala substitution blocks p53-dependent apoptotic activity, 31 and relieves the suppression of cell cycle progression that mediated by p53. 32 In glioma cells, p53 apoptotic activity was restrained, partially due to the p53 Ser15 mutation. 33 Previous study has shown that the phosph-p53 (Ser15) was increased in breast cancer MCF7 cells with decreased content of ARL2. 28 Here, miR-497-5p overexpression reduced ARL2 protein expression but elevated P-p53 protein expression in OS MG-63 and U2OS cells. We thus supposed that the expression of P-p53 were upregulated in OS cells might be ascribed to decreased ARL2 expression levels. These studies collectively suggest that miR-497-5p overexpression led to activation of p53-dependent apoptosis and induction of G0/G1 cell cycle arrest in OS cells.
Microtubules are an important component of cytoskeleton formed from tubulin polymers and play a key role in mitosis and cell migration. 34 In recent years, microtubules-targeted drugs like taxol are widely applied for treating various kinds of human cancers such as breast cancer, lung cancer, prostate cancer, and so on. 35 Azarenko et al. 36 showed that sulforaphane strongly inhibited proliferation and mitosis though suppression of microtubule dynamic instability in breast cancer MCF-7 cells. Griseofulvin depresses the dynamic instability of the microtubules, and contributes to reduced mitosis in HeLa cells. 37 Beghin et al. 27 pointed out that the microtubule dynamic instability was obviously boosted in highest ARL2 expression but hampered in lowest ARL2 expression level compared with control breast cancer MCF7 cells. In the current study, reduction of microtubule dynamic instability might have occurred due to the downregulation of ARL2 in the miR-497-5p-overexpressed OS cells, thus contributing to induction of G0/G1 mitotic arrest. The identification of miR-497-5p in OS may provide a potential target in antimitotic anticancer therapies.
In conclusion, overexpression of miR-497-5p inhibits proliferation and induces apoptosis in OS cells, possibly though direct targeting of ARL2. Our findings on miR-497-5p are encouraging and provide important insights into the mechanism of OS occurrence and development.
Footnotes
Disclosure Statement
There are no existing financial conflicts.