
Abstract
Background:
Suppression of Aurora kinase A (Aurora-A, AURKA) by siRNA of Aurora-A (siAurora-A, siA) has been used in lung tumor treatment. However, the dose and frequency of gene transfection still need to be confirmed further. We imitated multiple administration of solid tumor and attempted to make out the effects of thrice transfection of siAurora-A on cilia generation and apoptosis of SK-MES-1 cells (SK) or A549 cells.
Methods:
The Aurora-A mRNA levels of cells cultured with serum for 6 d or without serum for 2, 4, or 6 d were examined with real-time quantitative PCR; Cells were transfected single or repeatedly with siAurora-A or siControl (siC), their Aurora-A mRNA levels were determined with PCR; Their cilia were examined with immunohistochemistry. Cell viability was measured with the MTT assay. Protein expression was analyzed with western blot.
Results:
Cell viability showed a downward trend along with the prolongation of starvation time to the second, fourth, and even to the sixth day in both types of cells. But, the expression level of Aurora-A mRNA flipped to rise at the sixth day instead of decreasing at the fourth day. Protein expression trend of total Aurora-A in the two groups was consistent with Aurora-A mRNA expression trend. Compared with siC-3 group (transfected three times with siControl), siAurora-A significantly reduced the Aurora-A mRNA expression in siA-3 group (transfected three times with siAurora-A). Similarly, the cell viability of siA-3 group was lower than that of siC-3 group. The cell viability of siC-3 group was higher than that of serum-free-6d group, but, levels of Aurora-A mRNA expression of siC-3 group had no difference with serum-free-6d group. Finally, among groups transfected once or three times or starved for 6 d, there was no significant difference of ciliated cell proportions in both types of cells respectively.
Conclusions:
Repeated siAurora-A transfection decreased Aurora-A expression that resulted in effective suppression proliferation of SK-MES-1 or A549 cells, but did not affect cilia generation.
Introduction
Large amounts of DNA duplicated during the S phase of the cell cycle are segregated into two daughter cells during the M phase. Usually G1 and G2 phases separate S phase and M phase in most cells. Proliferating cells control precisely DNA condition of every phase, especially in the proliferating G1 phase. 1 Cells may stay in stable G0 phase for a long time if conditions of cell proliferation are not ready. However, the mechanism of cell cycle reentry from G0 into G1 has not been well understood. A derived from mother centriole (a centriole which is inherited from mother cell, not the replicated one), static, microtubule-rich primary cilium can be found on the surface of many types of resting cells. 2 Primary cilia may be chemosensors or mechanosensors. Its dysfunction has relationship with some diseases such as brain malformation, obesity, diabetes, and so on. 3,4 Some studies have shown that cilia were lack in most cancer cells. 5 –10 When cells reenter the cell cycle from G0, disassembling of primary cilia is promoted in many types of cells 11 –13 Cilia generation appears to be inversely related to cell cycle progression. However, Cilia still were kept during exceptional cell proliferation. 11,13,14 –17 Therefore, it has been discussed for a long time whether a primary cilium negatively regulates cell cycle.
Recent studies have indicated that the Aurora kinase family's frequent overexpression in human cancers has a relationship with oncogenic transformation. 18 Three Aurora kinases, Aurora-A (AURKA), -B, and -C, have been identified in humans, among which, the overexpression of Aurora-A occurs in some types of cancers 19 –21 including non-small-cell lung cancer. 22,23 Aurora-A localizing to centrosomes and mitotic spindles can drive mitotic entry, centrosome maturation, centrosome separation, and bipolar spindle formation. 24 –27
Aurora-A is considered negatively regulated ciliary dynamics in proliferating cells. 28 On the one hand, Aurora-A disassembles cilia at cell cycle reentry, on the other hand, Aurora-A persistently inhibits primary cilia regeneration during cell proliferation. 28,29 A study showed that ciliary reassembly is suppressed by trichoplein-mediated Aurora-A activation pathway in growing cells. Serum starvation induced transient Ndel1 degradation, subsequent to the disappearance of trichoplein at the mothercentriole. Thus, serum starvation can induce primary cilia assembly by suppressing Ndel1-trichoplein-Aurora-A pathway. 30 Some studies showed deacetylation on axonemal α-tubulin may be related to ciliary disassembly. 31 Histone deacetylase (HDAC6) can be activated by Aurora-A to remove acetylated group of axonemal α-tubulin. But, some studies have opposed the role of α-tubulin acetylation in the assembly or function of primary cilia.
With the exception of some cells possessing primary cilia during cell proliferation, cells begin to retract their primary cilia at cell cycle reentry. It remains controversial whether or not (de)ciliation affects cell cycle progression. 32 Some studies showed that Aurora-A suppressed ciliary formation and knockdown of Aurora-A by Aurora-A inhibitors or siRNA increased ciliary formation in some types of cells. To tumor cells, the inhibition of Aurora-A can induce false mitosis, whereas, to healthy cells, it may only cause cilia assembly and exit of the cell cycle. 28 This different cellular reaction made Aurora-A inhibition become a valid anticancer means, 32 however, some studies should be carried out to confirm its possibility.
On this basis, we are the first to repeatedly transfect siAurora-A, which mimics repeated clinical treatment of solid tumors, to evaluate effects of siAurora-A on suppression of proliferation and ciliary formation of SK-MES-1 cell and effects on ciliary formation of A549 cells.
Materials and Methods
Materials
PCR primers of GAPDH and Aurora-A were obtained from Takara Biotechnology (Dalian, China). Aurora-A: sense primer sequence (Sq.), 5′-GAAGCAATTGCAGGCAACCA-3′; antisense primer Sq., 5′-GAGGGCGACCAATTTCAAAG-3′. GAPDH: sense primer Sq., 5′-CCACCCATGGCAAATTCCATGGCA-3′; antisense primer Sq., 5′-TCTAGACGGCAGGTCAGGTCCACC-3′.
Cell culture
SK-MES-1 cells, human lung squamous carcinoma cells, grew in cell growth medium containing minimum essential medium (MEM). Human lung carcinoma A549 cells, a human alveolar epithelial cell line, grew in growth medium containing DMEM. Both MEM and DMEM were supplemented with 10% fetal bovine serum (FBS), 56 U/mL penicillin-G, and 56 μg/mL streptomycin sulfate. The cells were seeded at an initial density of 100,000 cells in six-well dishes (Costar, NY). Cells were cultured normally until the sixth day. During this process, medium was changed every 2 d. Cells in other groups were cultured without serum until the second, fourth, or sixth day. All cells were recovered respectively to detect indicators.
Transfection with siRNAs
siAurora-A,Sq., 5′-(TC)CCAGCGCATTCCTTTGCAA-3′, was purchased from QIAGEN Sciences, Inc. (Germantown, Germany). Negative control siRNA (siC, catalogue No. 1027310) was purchased from QIAGEN. In all transfections, exponentially growing SK-MES-1 or A549 cells were used. Cells were transfected with siRNA according to siRNA transfection protocol of jetPRIME (Polyplus-transfection, Inc., NY). The timelines of normal single transfection, repeated transfection, and single transfection were showed in Figure 1. The images in Figure 2 and Figure 3 were obtained using a microscope IX53 (Olympus, Tokyo, Japan), equipped with a Q imaging camera (Qimaging, BC, Canada). Images were taken at room temperature. Two or three researchers observed the cells at least three times in each case, and took representative photographs.
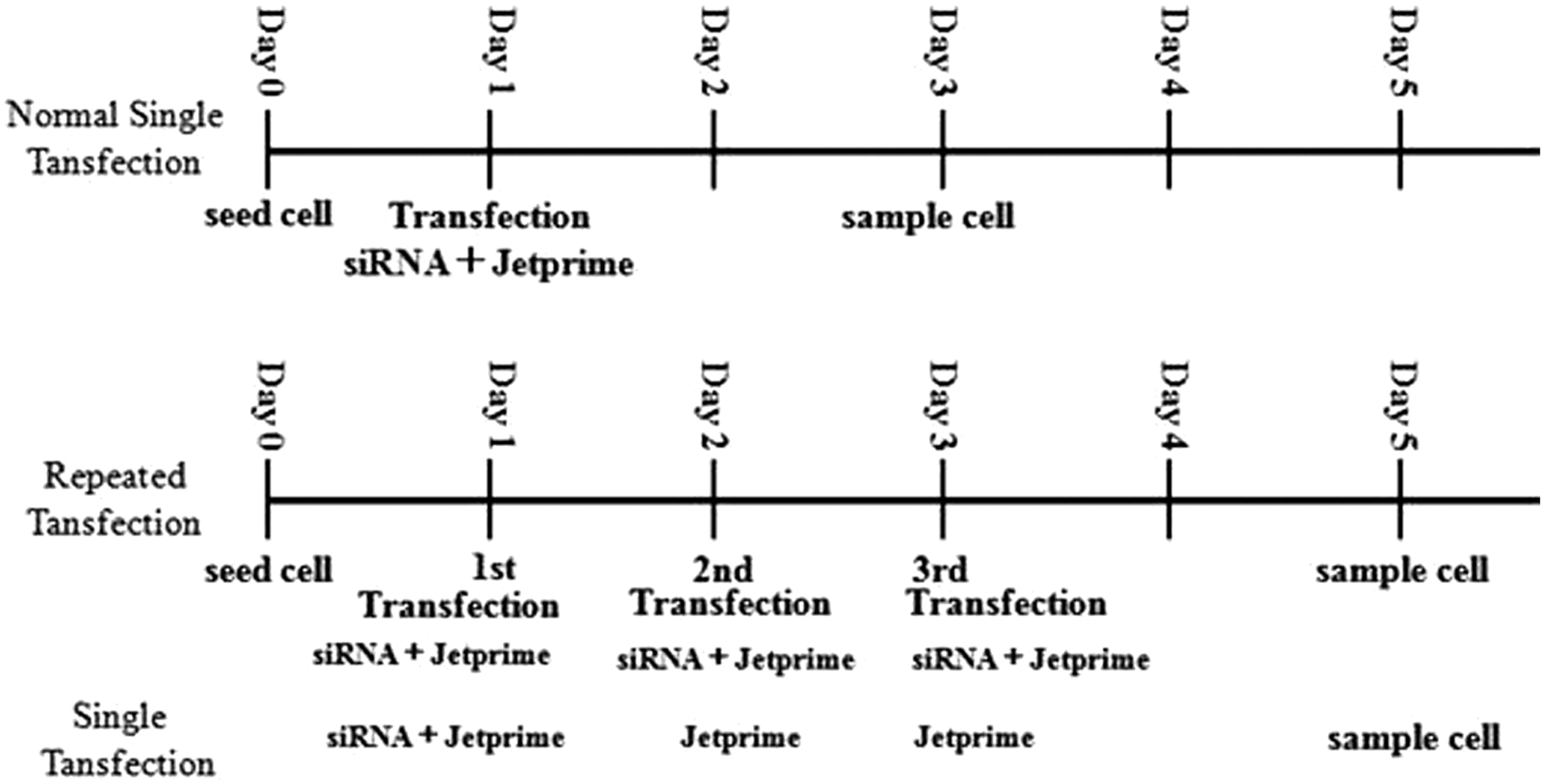
Timeline of the study.
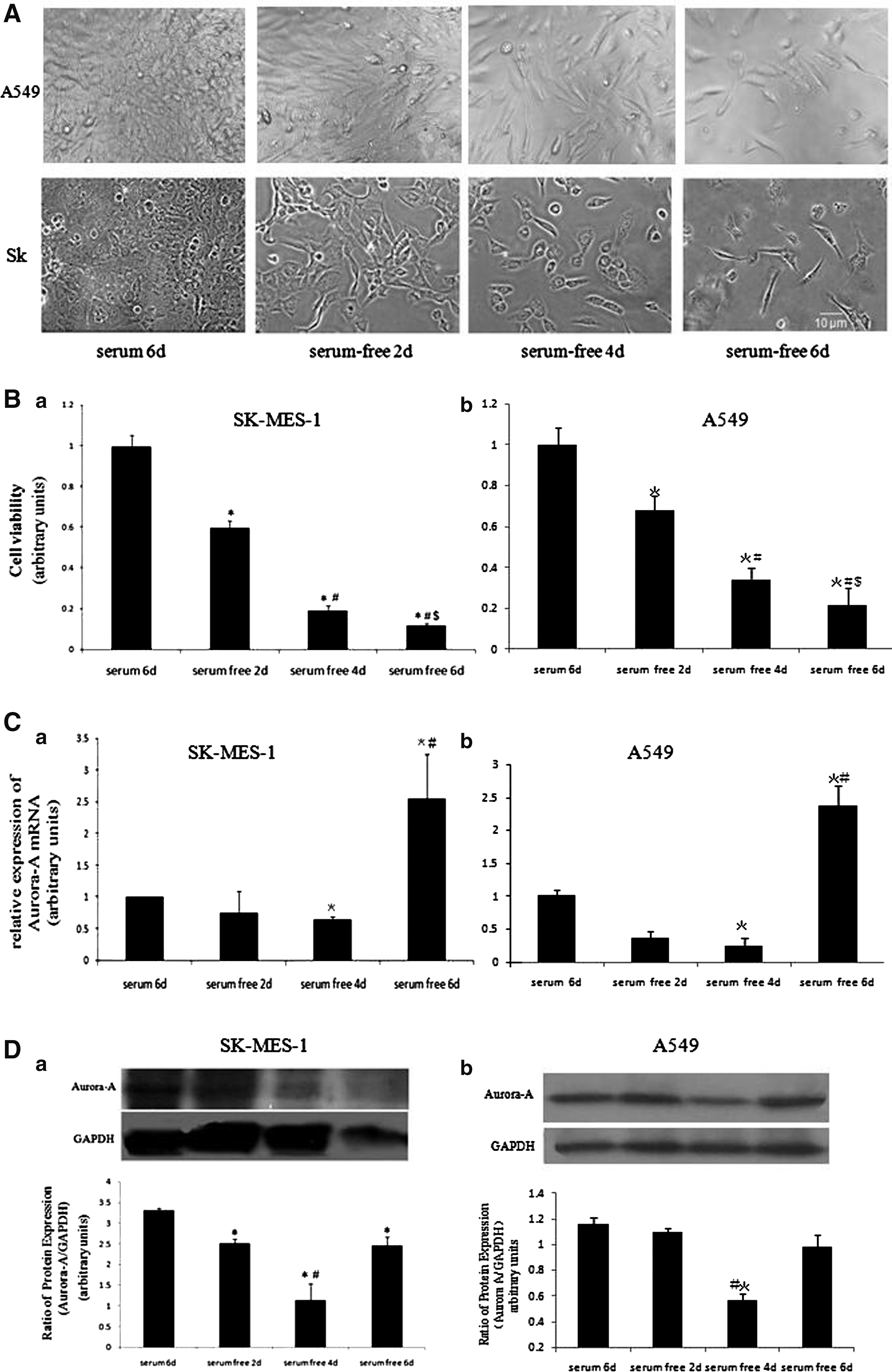
Effects of starvation time on Aurora-A mRNA expression, cell viability, and protein expression. After SK-MES-1 or A549 cells reached 50% confluence, culture was continued with serum-free medium until the second, fourth, and sixth day. Images were taken before cells were sampled respectively to detect Aurora-A mRNA expression. The same experiment was repeated three times.
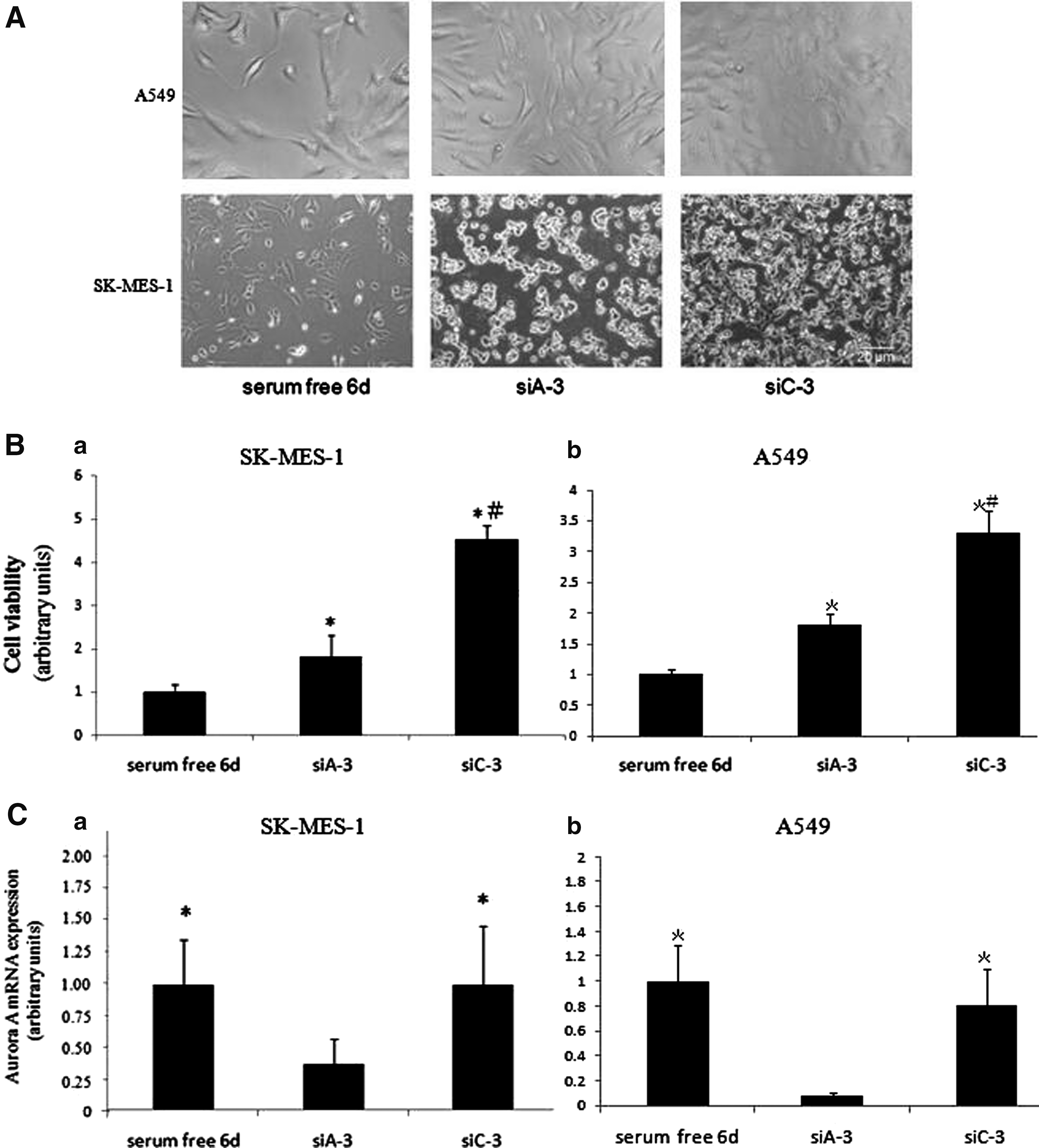
(
Normal single transfection
A total of 100,000 cells were seeded in per well of six-well plate with 2 mL growth medium 24 h before transfection to ensure that cells were 50% confluent at the time of transfection. And, 110 pmoles of siRNA was diluted in 200 μL of jetPRIME buffer for a final concentration of 50 nM/well, and mixed by pipetting up and down. Four microliters of jetPRIME reagent was added, followed by vortexing for 10 s and brief centrifugation. The mixture was incubated for 10–15 min at room temperature. The transfection mixture was added to the cells in serum-containing medium. The plate was gently rocked back and forth, returned to the incubator to incubate for 4–6 h. The transfection medium was changed with cell growth medium. After 48 h, cells were analyzed as required.
Repeated transfection
In the repeated transfection experiments, the first transfection was performed according to normal single transfection protocol like mentioned before. The transfection medium was replaced with cell growth medium after the first transfection and cells were cultured for 24 h, but not 48 h, until the second transfection. The second and third transfections were then carried out just like the first transfetion. But, after the third transfection was performed, cells were cultured continuously in cell growth medium for 48 h and then were analyzed. The cells in the single transfection were transfected with siRNA only once. And in the second and third transfection processes, cells were treated only with the transfection reagent Jetprime, but not siRNA. These experiments were independently performed at least thrice, each in triplicate.
Cell viability
Cell viability was evaluated by the MTT (3–4, 5-dimethylthiazol 2,5-diphenyltetrazolium bromide) assay. Cells were seeded into 96-well plates containing 200 μL MEM for SK cells or DMED for A549 cells with 10% FBS at 0.6 × 104 cells initial density into each well. Before cell viability was detected, the medium was changed to 180 μL fresh MEM or DMEM with 10% FBS to each well. Twenty microliter MTT (5 mg/mL formulated with PBS, pH 7.4) was dropped in each well and cells were incubated for additional 4 h. Then the supernatant was carefully aspirated, subsequently 150 μL DMSO was dropped in each well and shaken for 10 min to fully dissolve the crystals. A wavelength of 562 nm was selected to measure the absorbance of each well in the enzyme-linked immunosorbent monitor (Kehua KHB-ST-360, Shanghai, China).
Real-time PCR
Total RNA was extracted by using RNAiso Plus (TaKaRa) as the lysis buffer. cDNA was synthesized with a Prime Script RT reagent Kit (TaKaRa). Reverse transcription PCR was carried out with the primers mentioned in Materials. Real-time PCR was performed with SYBR Premix Ex Taq II in StepOne AB Applied Biosystems (Life Technologies, CA).
Western blot analysis of protein expression
Sodium dodecyl sulfate polyacrylamide gel ectrophoresis was performed and then samples were electrophoretically transferred to polyvinylidene difluoride membrane. Five percent nonfat milk blocked the membrane for 2 h, then, the blots were incubated with primary antibodies overnight at 4°C. Tris-buffered saline with 0.05% Tween 20 was used to rinse the blots and then the blots were incubated with second antibody for 2 h at room temperature. Immobilon Western Chemiluminescent HRP Substrate (Millipore Corporation, MA) were used to detect Immunoreactive bands, and image analysis program (Scion Corporation, MD) was used to quantify band intensities. Commercial antibodies and their dilution were as follows: mouse anti-Aurora-A diluted at 1:500 (AA921; Beyotime, Nantong, China), Rabbit anti-GAPDH diluted at 1:1000 (BA2913; Boster, Wuhan, China), Goat anti-Rabbit IgG diluted at 1:1000 (BA1054; Boster), Goat anti-Mouse IgG diluted at 1:1000 (A0216; Beyotime).
Immunofluorescence microscopy
Cells were seeded on coverslips (Iwaki Glass Co., Ltd.). The anti-acetylated tubulin (Cell Signaling Technology) was used to stain cilia according to immunostaining protocol. Before fixation, cells were incubated on ice for 30 min, then, methanol at 20°C fixed cells for 10 min. PBS was used to rinse cells at room temperature for 15 min. Subsequently, cells were blocked with 3% BSA/PBS for 30 min followed by incubation with primary antibodies. After PBS-washing three times, cells were incubated with secondary antibodies Alexa Fluor 488 (Invitrogen, CA) for 30 min. Fluorescence images in Figure 4 were obtained by using a microscope IX53 (Olympus, Tokyo, Japan), a Plan Apochromat 100 × /1.40 NA oil immersion lens (Olympus), and Q imaging camera (Qimaging, BC, Canada). Images were taken at room temperature and two or three researchers observed the cells at least three times in each case, and took representative photographs. Images were further analyzed with Photoshop Elements 6.0 (Adobe, CA).

Cilia were probed by immunohistochemical method. Cells were immunostained after single transfection or repeat transfection (three times) with siAurora-A or siControl, or starvation culture for 6 d. Representative photographs were taken under a Plan Apochromat 100 × /1.40 NA oil immersion lens. siA-1: cells were single transfected with siAurora-A; siA-3: cells were transfected three times with siAurora-A; siC-1:cells were single transfected with siControl; siC-3: cells were transfected three times with siControl; serum-free 6d: cells were cultured without serum for 6 d. For A549 cells, other groups include siA-1, siA-3, siC-1, and siC-3. The images of centrsomes were similar. Only one image was employed to represent them.
Statistical analysis
All data are presented as the means ± SD. A statistical comparison between groups was carried out using the Student's t test. p < 0.05 was considered statistically significant.
Results
The prolongation of serum starvation did not cause sustained reduction of Aurora-A mRNA expression
According to cell culture in Materials and Methods section, cells were cultured without serum and then cells were sampled at different time points such as the second, fourth, and sixth day to detect the Aurora-A mRNA expression level by real-time quantitative PCR. Figure 2A showed images of cells cultured with serum for 6 d, without serum for 2 d, 4 d, and 6 d. In Figure 2B, MTT values showed the cell viability in the four groups. We observed, for both SK and A549 cells, the longer starvation time, the lower the cell viability, and the morphology of cells has changed, from the full to slender. The cell viability in the serum-free-6d group was obviously lower than that in the serum-free-2d group. We detected the affection of starvation time on mRNA expression of Aurora-A by real-time PCR. Figure 2C showed as the culture time was prolonged, the Aurora-A mRNA expression level varied. When cells were cultured without serum for 4 d, mRNA expression level was significantly lower than that of cells normally cultured 6 d with serum (p < 0.05). Instead, the mRNA expression level of cells cultured without serum for 6 d in serum-free-6d group was significantly higher than those of the serum-6d groups and serum-free-4d group (p < 0.05 vs. serum-6d, or serum-free-4d group). Protein expression trend of total Aurora-A in the two types of cells was consistent with Aurora-A mRNA expression trend. Protein expressions of total Aurora-A were gradually decreasing over starvation time until the fourth day, but flip to increase at the sixth day. But for A549, the decrease of protein expressions of total Aurora-A in serum-free-2d group was not so significant as that of SK-MES-1 cells and the increase of protein expressions of total Aurora-A in serum-free-6d group was more significant than that of SK-MES-1 cells.
The effect of repeated transfection of siAurora-A on cells
Cells in monolayer culture were repeatedly transfected three times with siAurora-A or siControl, cell viability, and Aurora-A mRNA expression were examined. Figure 3A showed images of cell cultured without serum for 6 d or transfected three-times with siAurora-A or siControl. For the two types of cells, the amount of cell in siA-3 group was obviously lower than that in siC-3 group. In Figure 3B, MTT values showed the cell viability of siA-3 group transfected three times with siAurora-A was lower than that in siC-3 group transfected three times with siControl. Figure 3C showed, three times transfection of siAurora-A in siA-3 group significantly reduced the expression of Aurora-A mRNA, compared with siC-3 group (p < 0.05), but, mRNA expression of Aurora-A in serum-free-6d group did not decrease more than that in siC-3 group.
The effect of siAurora-A on cilia generation of cells
Cilia were detected by immunocytochemistry method. Whatever cells were cultured without serum for 6 d in serum-free-6d group or cells were singly or repeatedly transfected with siRNA-AuroraA or siRNA-Control (Fig. 4), there were no significant differences in proportion of cillia among five groups. Their proportions of generating cilia cell were about 7%–10% for SK-MES-1 cells. Generating cilia cells were not observed in A549 cells singly or repeatedly transfected with siRNA-AuroraA or siRNA-Control, but generating cilia cells were observed in serum-free-6d group of A549 cells and the proportion was very low. One generating cilia cell could be found among about 300 A549 cells and the cilium was short.
Discussion
First, our results showed that cell viability of SK-MES-1 cells and A549 cells in serum-free medium condition significantly decreased with culture time prolonged. Compared with the normal cultured cells in serum-6d group, cell viability in serum-free groups (serum-free-2d group, serum-free-4d group, and serum-free-6d group) gradually declined along culture time extending, consistent with our speculation. Aurora-A mRNA expression levels in serum-free-4d group more significantly declined, compared with that of serum 6d group. Aurora-A was involved in tumor proliferation process. When tumor cells were under starvation status without serum, their growth rate was reduced down, followed dropdown of mRNA expression levels of Aurora-A. Serum starvation induced the disappearance of trichoplein at the mother centriole, subsequently, trichoplein could not active Aurora-A to inhibit primary cilia assembly, 31 so serum starvation induced primary cilia assembly. However, we noted when cells were cultured in serum-free medium for 6 d, cell proliferation was suppressed, but, the mRNA expression level of Aurora-A was reversely increased in the two types of cells.
This phenomenon has been observed in our previous study, 33 when A549 Lung adenocarcinoma cells were transfected repeatedly with siControl to the fifth day, the amount of dead cells increased, but the mRNA expression levels of Aurora-A had reversely risen. We thought it was possible that the self-protection mechanism of tumor cells had been switched on. When the cells suffered crowning calamity, such as under the extreme hunger status or damage status induced by cytotoxicity from the whole transfection system, to avoid persistent decline of Aurora-A mRNA, the ongoing process timely increased the synthesis of Aurora-A mRNA in feedback manner, and thereby ensured tumor cells reenter proliferation status. Just as tumor cells can gain resistance to drug, when the conditions change, and this change reach a certain extent, the tumor cells may gain self-protection ability by gene mutation. This reverse increase of Aurora-A mRNA expression in serum starvation for 6 d case was not what we expected, and serum starvation also would injure normal cells in the course of clinical treatment.
Gene target therapy was still the best choice. Then, in this study, we found that repeated transfection of siAurora-A decreased expression of Aurora-A mRNA in siA-3 group, compared with siC-3 group, in the two types of cells. Actually this part experiment of the study for A549 has been published. 33 To detect the effect of repeated transfection of siAurora-A on SK-MES-1 cells, A549 cells were used to compared with SK-MES-1 cells. The result showed siAurora-A was also able to decrease the expression of Aurora-A mRNA in SK-MES-1 cells, even in a high expression status of Aurora-A mRNA triggered by the cytotoxicity from the whole transfection system on the fifth day. Such result suggests the cytotoxicity from the whole transfection system will not threaten efficacy of generic drugs in future clinical treatment.
Finally, we found occurrence frequencies of cilia involved in cell proliferation were similar to each other group in the two types of cells, although with different administration. Previous studies have reported, blocking of Aurora-A mRNA could improve the ratio of ciliated cells, and the emergence of cilia was able to arrest cell proliferation cycle processes. So in the study, the reduced expression of Aurora-A mRNA induced by siAurora-A should increase the proportion of ciliated cells. But, actually the ratios of ciliated cells in siA-3 group and siC-3 group were not statistically different in the two types of cells. It might be related to proliferation status of cancer cells. Why induction/absorption of a primary cilium affects cell cycle progression becomes a new question. One possible explanation is disassembled primary cilium may serve as templates for centriole duplication in the S phase. On the other hand, there are some exceptions many ciliated protozoans may duplicate their centrioles without deciliation. 34 Therefore, whether entry into the S phase needs the absence of a primary cilium may rely on cells in which deciliated centrioles are a prerequisite for duplication templates.
We noted, there may be the third possibility in SK-MES-1 or A549 cells. Although inhibiting Aurora-A could not successfully induce primary cilia, deciliated centrioles in cells were not able to serve as duplication templates. Their proportions of generating cilia cell in serum-free-6d, siA-1, siA-3, siC-1, and siC-3 groups were about 7%–10% for SK-MES-1 cells. There were no significant differences in proportion of cillia among five groups. Generating cilia cells were not observed in siA-1, siA-3, siC-1, and siC-3 groups for A549 cells, only in serum-free-6d group of A549 cells, one generating cilia cell could be found among about 300 A549 cells and the cilium was short. Why siAurora-A could not induce primary cilium assembly in SK-MES-1 or A549 cells needs further investigation to clarify it. We are the first to detect the relationship between cilia and Aurora-A in SK-MES-1 and A549 cell line. Due to lack of other relevant data, the current results could not be given an explicit explanation. The further studies have to be carried out to confirm the information.
Conclusions
Our results showed that repeated transfection with siAurora-A effectively suppressed proliferation of SK-MES-1 cells. But, the suppression has no relationship with cilia regeneration in the two types of cells. These findings may provide some support when gene therapy-based strategies are designed to patients with lung cancer.
Footnotes
Acknowledgments
This work was supported by the 12th Five-year Project of Medicine (No. BWS12J007, to Z.M.); the Natural Science Foundation of Liaoning Province, China (No. 2011225008, to Z.M.); The fourth batch of Liaoning Province Science and Technology Program, China (No. 2015225020, to Z.M.); China Postdoctoral Science Foundation (No. 2013M542470, to Z.W.); Natural Science Foundation of Liaoning Province, China (No.2015020424, to Z.W.); and the Students' Innovation and Entrepreneurship Training Program of Liaoning Province, China (No. 201610164006, to Z.W.).
Authors' Contributions
Z.M.: conceived and designed the study; Z.W and J.C.: performed the experiments and data acquisition; Z.M., Z.W.: performed statistical analysis; Z.W.: wrote the article; Z.M. and Z.W.: reviewed and revised the article. All authors have read and approved the article to be published.
Disclosure Statement
No competing financial interests exist.