
Abstract
Cancer Biotherapy and Radiopharmaceuticals
officially retracts the paper entitled, “miR-155 Affects Osteosarcoma MG-63 Cell Autophagy Induced by Adriamycin Through Regulating PTEN-I3K/AKT/mTOR Signaling Pathway” by Lin Wang, Bing Tang, Heng Han, Dan Mao, Jie Chen, Yun Zeng, and Min Xiong (Cancer Biother Radiopharm 2018;33(1):32–38; doi: 10.1089/cbr.2017.2306) due to the discovery that the paper was submitted from a paper mill, which is a violation of the journal's standard protocols.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves and does not tolerate any violations of scientific misconduct.
Introduction
Osteosarcoma is a common primary malignant bone tumor. Although chemotherapy improves the survival rate and prognosis, more than 1/3 of patients showed chemoresistance and eventually led to treatment failure. 1,2
Autophagy is the process of autolysosome formation to digest and degrade abnormal proteins and organelles, thus is of great significance in maintaining cell homeostasis, genome stability, and preventing cancer. 3 Autophagy is closely associated with cancer cell occurrence, development, outcome, and treatment effect. It was reported that tumor cell proliferation and apoptosis may change after downregulation of autophagy treated by anticancer drugs. 4 Autophagy level was related to chemosensitivity. PI3K/AKT/mammalian target of rapamycin (mTOR) signaling pathway is mostly explored in regulating cell autophagy. mTOR, as an important effect protein of PI3K/AKT/mTOR signaling pathway, can negatively regulate cell autophagy by inhibiting the formation of ULK complex and blocking endoplasm reticulum membrane fall off to form autophagosome membrane. 5,6
Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) is a cancer suppressor gene with dual activity of proteinase and phosphatase that can restrain PI3K/AKT/mTOR signaling pathway. Therefore, it exhibits a positive regulatory role on cell autophagy. PTEN-PI3K/AKT/mTOR signaling pathway is closely related to the occurrence, progress, prognosis, and chemosensitivity of breast cancer, 7 lung cancer, 8 prostate cancer, 9 and bladder cancer. 10
MicroRNA is a type of small noncoding RNA in eukaryote at the length of 18–22 nt. It regulates cell proliferation and apoptosis by complete or incomplete binding with target mRNA to promote mRNA degradation or inhibit its translation. 6 It was showed that miR-155 significantly increased in osteosarcoma tissue, 11 suggesting its oncogene role in the pathogenesis of osteosarcoma. Bioinformatics analysis revealed the targeted binding site between miR-155 and the 3′-UTR (untranslated region) of PTEN mRNA. This study investigated the role of miR-155 in regulating osteosarcoma cell autophagy, chemosensitivity to Adriamycin (ADM), and PTEN-PI3K/AKT/mTOR signaling pathway.
Materials and Methods
Main reagents and materials
Human osteosarcoma cell MG-63 was purchased from Cell Bank in Chinese Academy of Sciences. Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were bought from Hyclone. RNA extraction reagent TRIzol was obtained from Invitrogen. ReverTra Ace qPCR RT Kit and SYBR Green were derived from Toyobo (Japan). Anti-miR-NC, anti-miR-155, miR-155 mimic, and mimic NC were obtained from Ambion. Polymerase chain reaction (PCR) primers were synthetized by Sangon (Shanghai, China). PTEN, AKT, and p-AKT antibodies were purchased from Abnova. mTOR, p-mTOR, and Beclin-1 antibodies were bought from Santa Cruz. β-actin antibody was obtained from GeneTex. HRP labeled secondary was derived from Bio-Rad. Radioimmunoprecipitation assay (RIPA) was obtained from Beyotime (Jiangsu, China). FuGENE HD Transfection Reagent was purchased from Promega. Cell Apoptosis Detection and CCK-8 Kits were bought from Yeasen (Shanghai, China). Plasmid pSicoR was obtained from Addgene (United Kingdom). ADM was obtained from Hengrui Medicine (Jiangsu, China). Annexin V-FITC and Propidium (PI) Kit were from Thermo Fisher Scientific. A total of 25 male nude BALB/c nu/nu mice aged 6–8 weeks and weighing 20–22 g were purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China).
MG-63 resistant cell induction and resistance index calculation
MG-63 cells were cultured in DMEM containing 10% FBS and 1% streptomycin and maintained at 37°C and 5% carbon dioxide. The cells were treated by 0.1 mg/L ADM for 24 hours when the fusion reached 60%–70%. After cell recover by passage, ADM concentration was gradually increased to 0.2, 0.4, 0.8, and 1.6 mg/L. Finally, the cells kept stable in 1.6 mg/L ADM were identified as MG-63/ADM. The model of stable drug resistance was established by ADM. MG-63 cells were cultured for 24 h with ADM at a dose of 1.6 mg/L and then incubated in ADM-free medium. The MG-63/ADM cells were established on the basis of the normal human osteosacoma cells by long-term culturing with increasing amounts of ADM. After 4 months, ADM-treated cells acquired the ability that cells were all stable living in the medium of ADM (1.6 mg/L). MG-63 cells acquired obvious resistance to the light concentration of ADM, named MG-63/ADM.
MG-63 and MG-63/ADM cells were treated by ADM at 0, 0.025, 0.25, 0.5, 1.0, 2.5, 5, 10, and 20 mg/L for 48 hours. Cell viability was determined by CCK-8 Kit at 450 nm. IC50 was calculated by Excel software. Resistance index (RI) = IC50 at MG-63/ADM/IC50 at MG-63.
Dual luciferase assay
The full length fragment of PTEN 3′-UTR was connected to pGL3 luciferase reporter vector to form pGL3-PTEN-wt. The mutation of PTEN 3′-UTR was used to construct pGL3-PTEN-mut. FuGENE HD Transfection Reagent was applied to co-transfect 1 μg pGL3-PTEN-wt or pGL3-PTEN-mut with 50 nm/L miR-155 mimic to HEK293T cells. Dual luciferase activity was tested after 48 hours.
MG-63/ADM cell transfection
MG-63/ADM cells were divided into five groups, including anti-miR-NC, anti-miR-155, pSicoR-blank, pSicoR-PTEN, and anti-miR-155+pSicoR-PTEN group. The cells were treated by 1.6 mg/L ADM for 24 hours.
Establishment of an orthotopic lung cancer model in nude mice model and grouping
A total of 25 nude BALB/c nu/nu mice were intraperitoneally injected with pentobarbital sodium (10 mg/kg) for anesthesia and then fixed in the right lateral decubitus position after anesthesia. Then 0.2 mL A549 cell suspension (4 × 106 cells/mL) from groups of anti-miR-NC, anti-miR-155, pSicoR-blank, pSicoR-PTEN, and anti-miR-155+pSicoR-PTEN was percutaneously inoculated into the upper margin of the sixth intercostal rib on the right anterior axillary line to a depth of about 5 mm rapidly and after that, the needle was promptly pulled out (five mice/group). Nude mice were maintained in the right lateral decubitus position after injection and observed until complete recovery. Every 3 days, the long diameter (L) and short diameter (W) of tumor nodules in nude mice were measured by vernier caliper, and the volume of tumor (mm3) = 0.5236 × L × W 2 (mm3) and the tumor growth curve of each group were established.
Quantitive reverse transcription polymerase chain reaction
The tissue was grinded in liquid nitrogen and cells were collected by centrifugation at 10000 g for 1 min, which were then treated with Trizol (100 mg tissue: 1 mL Trizol, cells: 600 μl Trizol) (Invitrogen, USA). Then the solution was moved to an Ep tube and added with 200 μl chloroform. After vibrated for 15 s, the upper aqueous phase was added with 500 μl isopropanol for 10 min. After centrifuged at 12000 g for 10 min, the precipitation was added with 1 mL ethanol (75%). After centrifuged at 4°C and 7500 g for 5 min, the supernatant was removed and the tube was dried for 10 min. Next, the RNA was solved in DEPC water. RNA content and purity were determined by ultraviolet spectrophotometer. Reaction solution was prepared according to the instruction, including 2 μg total RNA, 1 μl oligo primer (50 μM), 1 μl dNTP mix (10 μM), and ddH2O. The solution was predegenerated at 65°C for 5 minutes. Next, cDNA first chain synthesis reaction system was prepared, including 2 μl 10 × RT buffer, 4 μl MgCl2 (25 μM), 2 μl DTT (0.1 M), l μl RNAase OUT (40 U/μl), 1 μl SuperScrip III RT (200 U/μl), and ddH2O (Promega, USA). The reaction condition was composed by 50°C for 50 min and 85°C for 5 min. Real-time quantitive PCR was then performed by using SYBR Premix Ex Taq GC kit (Takara, Japan) (7.5 μl 2×premix, 10 mM forword and reverse primers, dH2O to a final volume of 15 μl) in the following condition: 94°C denature for 30 sec, followed by 40 cycles each containing 94°C denature for 5 sec, and 60°C annealing for 30 sec with LightCycler 480 (Roche Diagnostics, USA). The primer sequences used were as follows: miR-155PF: 5′-ACGCTCAGTTAATGCTAATCGTGATA-3′, miR-155PR: 5′-ATTCCATGTTGTCCACTGTCTCTG-3′; U6PF: 5′-ATT GGAACGATACAGAGAAGATT-3′, U6PR: 5′-GGAACG CTTCACGAATTTG-3′; PTENPF: 5′-AGGGACGAACTG GTGTAATGA-3′, PTENPR: 5′-CTGGTCCTTACTTCCCC ATAGAA-3′; Beclin-1PF: 5′-GGTGTCTCTCGCAGATTC ATC-3′, Beclin-1PR: 5′-TCAGTCTTCGGCTGAGGTTCT- 3′; β-actinPF: 5′-GAACCCTAAGGCCAAC-3′, β-actinPR: 5′-TGTCACGCACGATTTCC-3′. β-actin was selected as internal reference. Relative gene expression was semiquantitative analyzed by 2-ΔΔCt method. 2-ΔΔCt = gene copy number in test group/gene copy number in control. Experiments were carried out in triplicates.
Western blot
Total protein was extracted by 600 μL RIPA on ice for 30 minutes and centrifuged at 4°C and 10,000 g for 10 minutes. The supernatant was moved to a new EP tube for quantification. A total of 50 μg protein was separated by 10% SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) and transferred to membrane. Next, the membrane was incubated in primary antibody at 4°C for 12 hours (PTEN, AKT, p-AKT, mTOR, p-mTOR, Beclin-1, and β-actin at 1:200, 1:300, 1:100, 1:300, 1:100, 1:200, and 1:800, respectively). Then the membrane was incubated in secondary antibody (1:5000) for 60 minutes after being washed by PBST thrice. At last, the protein expression was detected by enhanced chemiluminescence.
Flow cytometry
Cells were collected and incubated in 5 μL Annexin V-FITC and 5 μL PI avoiding light. Then the cell apoptosis was tested on flow cytometry.
Statistical analysis
All data analyses were performed on SPSS 18.0 software. The measurement data are depicted as mean ± standard deviation and compared by t-test. p < 0.05 was considered as statistical significance.
Results
miR-100 targeted inhibited mTOR expression
Bioinformatics analysis demonstrated the complementary binding site between miR-155 and the 3′-UTR of PTEN mRNA (Fig. 1A). Dual luciferase assay showed that miR-155 mimic and anti-miR-155 significantly declined and enhanced the relative luciferase activity in HEK293T cells, respectively (Fig. 1B), indicating the regulatory relationship between miR-155 and PTEN mRNA.
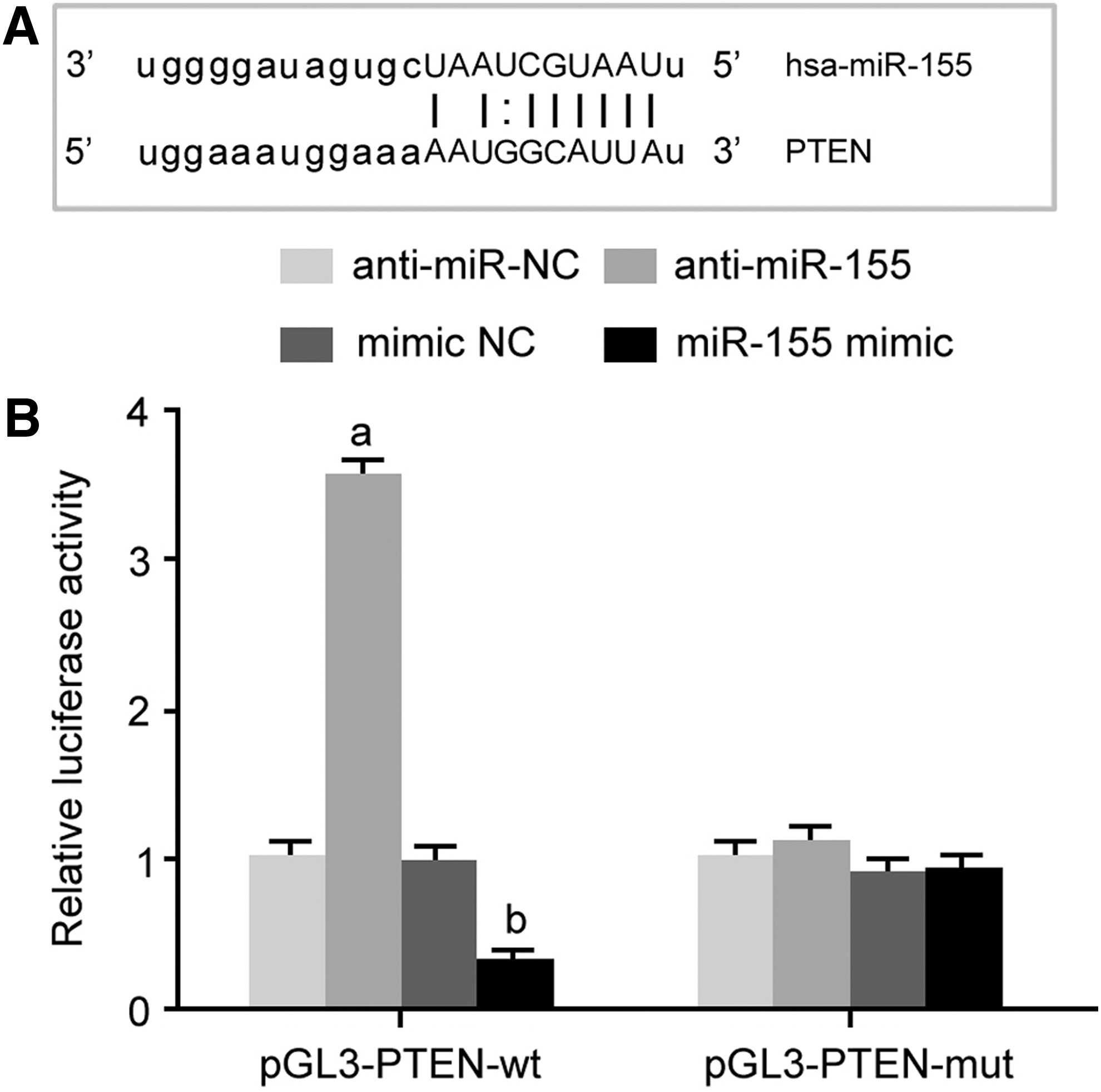
miR-155 targeted inhibited PTEN expression.
ADM significantly upregulated miR-155 expression and reduced PTEN level in MG-63/ADM cells
CCK-8 assay demonstrated that the sensitivity of MG-63/ADM cells to ADM was decreased compared with that of MG-63 cells (Fig. 2A). IC50 was 0.59 mg/L in MG-63 cells, while it was 9.37 mg/L in MG-63/ADM cells. RI was 15.88, suggesting the successful establishment of ADM resistant MG-63 cells. qRT-PCR demonstrated that miR-155 significantly upregulated, while PTEN and Beclin-1 mRNA obviously decreased in MG-63/ADM cells compared with MG-63 cells (Fig. 2B–D). Western blot revealed that p-AKT and p-mTOR markedly increased, whereas PTEN and Beclin-1 protein apparently reduced in MG-63/ADM cells compared with MG-63 cells (Fig. 2E). ADM treatment markedly elevated miR-155, p-AKT, and p-mTOR expressions, whereas reduced PTEN level. Beclin-1 was slightly upregulated, and autophagy and apoptosis levels were low (Fig. 2F, G). The changing ranges of miR-155, p-AKT, p-mTOR, and PTEN in MG-63 cells were smaller, while Beclin-1 expression, cell autophagy, and apoptosis were significantly enhanced. Their data demonstrated that ADM induced alternation of miR-155 and the interaction of PTEN and PI3K/AKT/mTOR signaling pathway (Fig. 3).

ADM significantly upregulated miR-155 expression and reduced PTEN level in MG-63/ADM cells.
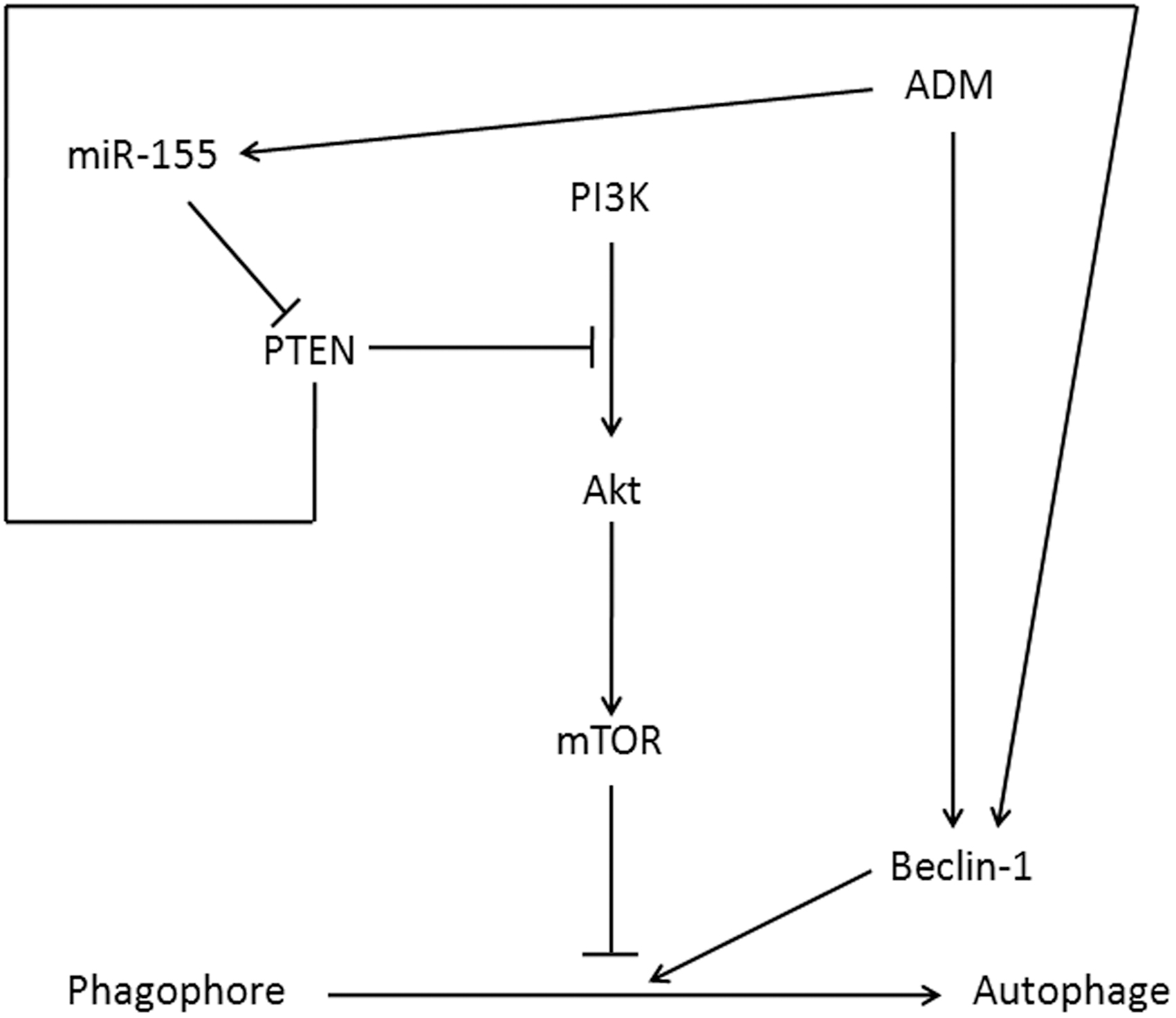
The sketch map of ADM induced alternation of miR-155 and the interaction of PTEN and PI3K/AKT/mTOR signaling pathway. mTOR, mammalian target of rapamycin.
Inhibition of miR-155 elevated PTEN expression and promoted osteosarcoma cell autophagy and apoptosis induced by ADM
Anti-miR-155 and/or pSicoR-PTEN significantly enhanced PTEN and Beclin-1 expressions (Fig. 4A, B), cell apoptosis, and autophagy induced by ADM (Fig. 4C) and declined p-AKT and p-mTOR levels.

Inhibition of miR-155 elevated PTEN expression and promoted osteosarcoma cell autophagy and apoptosis induced by ADM.
The tumorigenicity of osteosarcoma cells was reduced through the inhibition of miR-155 or elevation of PTEN
The result of tumorigenicity in nude mice showed that compared with the anti-miR-NC group, the tumorigenic potential of anti-miR-155 group was remarkably reduced. Moreover, compared with pSicoR-blank group, tumorigenicity in nude mice of pSicoR-PTEN group was obviously alleviated. The combined treatment of decrease of miR-155 and elevation of PTEN apparently impeded the tumorigenicity of osteosarcoma cells (Fig. 5).
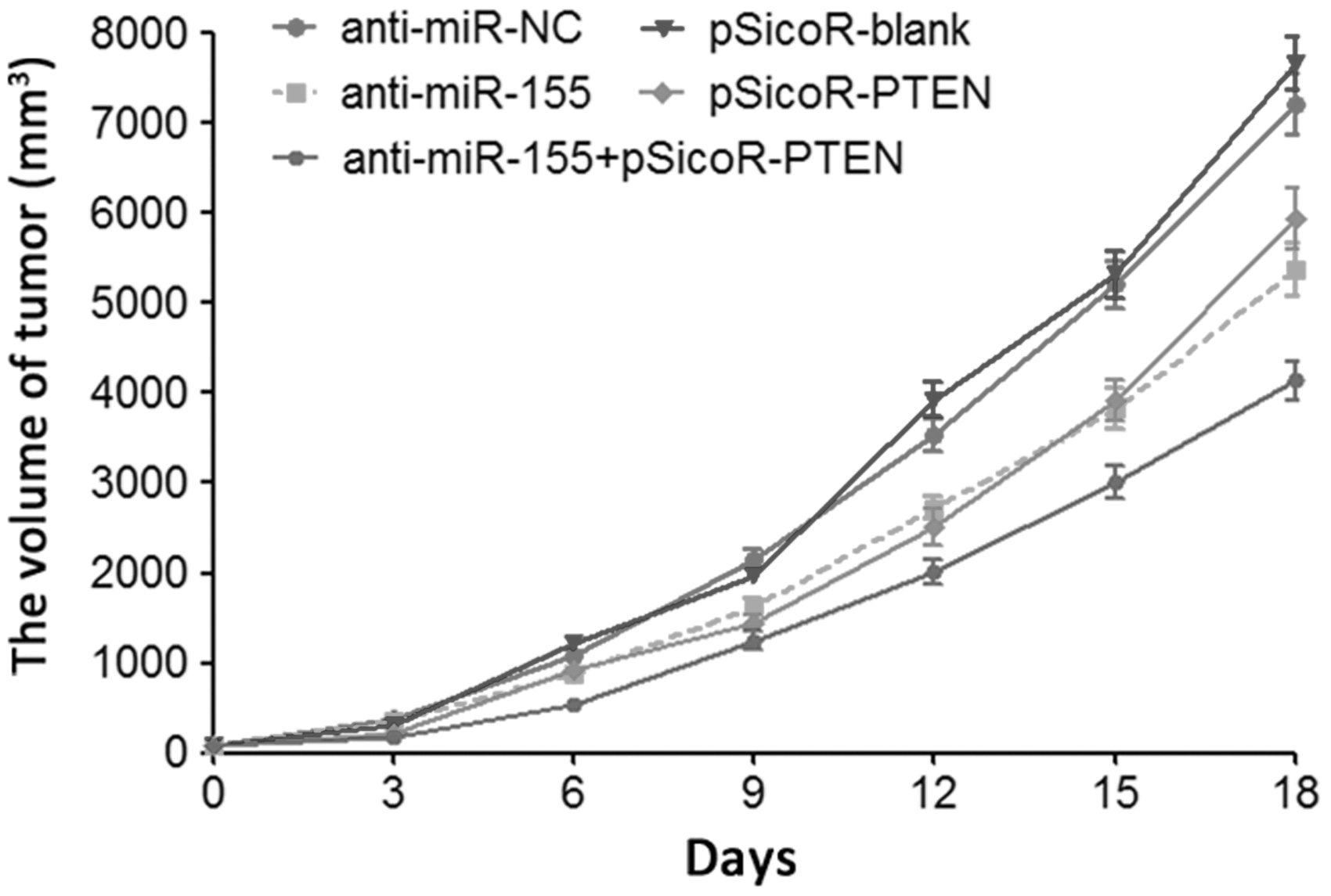
The tumorigenicity of osteosarcoma cells by evaluation of tumor volume.
Discussion
Their study demonstrated that miR-155 significantly increased in osteosarcoma tissue, indicating its oncogene role in the pathogenesis of osteosarcoma. 11 The authors conducted a bioinformatics analysis and revealed the targeted binding site between miR-155 and the 3′-UTR of PTEN mRNA. Their results showed that miR-155, p-AKT, and p-mTOR significantly increased, while PTEN obviously reduced in MG-63/ADM cells compared with MG-63 cells. It suggested that miR-155 elevation plays a role in suppressing PTEN expression, enhancing PI3K/AKT/mTOR signaling pathway, and promoting drug resistance. PI3K/AKT/mTOR signaling pathway widely existed in various tissues and cells. It plays an important role in cell autophagy, proliferation, cell cycle, apoptosis, and differentiation. mTOR is a direct regulatory protein to autophagy in PI3K/AKT/mTOR signaling pathway. mTOR activated by AKT can block the function of ULK-Atg13-FIP200 trimer complex through phosphorylating Atg13 to inhibit autophagosome formation and initialization. 12 In their study, the authors found that the downregulation of miR-155 inhibited the level of mTOR and induced the autophage by the elevation of PTEN.
In addition, Beclin-1 is the homologous gene of Atg6 in mammals that promotes autophagosome membrane formation and guides other autophagy protein localization. Therefore, it is the marker of autophagy. 13 The authors also revealed that either the treatment of ADM or the rise of PTEN level through the decrease of miR-155 activated the expression of Beclin-1 and in turn elicited the autophage process. Several studies revealed that PTEN plays a role in facilitating cell autophagy, enhancing cell sensitivity to radiotherapy 14,15 and chemotherapy. 4 Multiple studies have confirmed the role of PTEN downregulation in osteosarcoma drug resistance. 16 –18 Their data presented here indicated the relationship between PTEN expression and osteosarcoma cell autophagy.
Previous studies on the relationship between miR-155 and tumors illustrated that miR-155 level obviously upregulated in chemoresistant osteosarcoma cell line compared with drug sensitive cells, and miR-155 expression in ADM induced A549/dox was markedly higher than that in drug sensitive cell A549. 19,20 Moreover, miR-155 plays a role in enhancing drug resistance in breast cancer cells through inhibiting FOXO3a expression, and compared with tamoxifen sensitive breast cancer patients, miR-155 level was apparently increased in the breast tumor tissue of tamoxifen-resistant patients. 21,22 Similarly, in this study, the authors found that miR-155 expression in drug-resistant osteosarcoma cell line was significantly higher than that in drug sensitive cells, revealing the relationship between miR-155 and tumor cell resistance, which was similar with Chen et al., 19 Lv et al., 20 and Yu et al. 21
Of note, their data unraveled that Beclin-1 level was low in both of MG-63 and MG-63/ADM cells without the intervention of ADM, indicating low level of autophagy. However, the level of Beclin-1 was obviously elevated in MG-63 cells treated by ADM compared with MG-63/ADM, suggesting that ADM markedly induced MG-63 cell autophagy. ADM facilitated the apoptosis of MG-63 cells, while MG-63/ADM cells exhibited low sensitivity to cell apoptosis induced by ADM. It was also found that ADM obviously upregulated miR-155, p-AKT, and p-mTOR expressions, whereas reduced PTEN level in MG-63/ADM cells. Therefore, the authors proposed that miR-155 and PTEN-PI3K/AKT/mTOR were the important factors in affecting osteosarcoma cell autophagy induced by ADM.
In the therapy of tumor, He et al. reported that sorafenib apparently downregulated PTEN expression, while enhanced AKT phosphorylation and mTOR expression in hepatic cancer cells. 4 Indeed, PTEN expression and autophagy in drug-resistant HepG2-SR cells treated by sorafenib were significantly lower, while p-AKT and p-mTOR levels were higher than that in parental HepG2 cells, indicating that PTEN downregulation and PI3K/AKT/mTOR signaling pathway enhancement induced by chemotherapy were related to the reduction of autophagy sensitivity. In addition, Ning et al. demonstrated that PI3K/AKT/mTOR enhancement and autophagy inhibition caused by PTEN depletion obviously increased the trastuzumab resistance in breast cancer cells. 23 Their study showed that PTEN downregulation was associated with the reduction of autophagy, which was similar to He et al. 4 and Ning et al. 23 Their further detection found that anti-miR-155 and/or pSicoR-PTEN significantly enhanced PTEN and Beclin-1 expressions, cell apoptosis, and autophagy induced by ADM and declined p-AKT and p-mTOR levels.
Inhibition of miR-155 significantly accelerated breast cancer drug resistant cell MCF-7/TAM and SKBR3/TAM apoptosis, restrained cell proliferation, and declined drug resistance to tamoxifen. Their results demonstrated that suppression of miR-155 obviously enhanced tumor cell apoptosis and autophagy induced by chemotherapy and elevated chemosensitivity, which was in accordance with Lv et al. 20 and Shen et al. 22 This study revealed that miR-155 targeted suppressed PTEN expression, enhanced PI3K/AKT/mTOR signaling pathway, inhibited cell apoptosis and autophagy induced by ADM, and reduced sensitivity to ADM.
Conclusions
MiR-155 targeted inhibited PTEN level, upregulated PI3K/AKT/mTOR signaling pathway, restrained cell apoptosis and autophagy induced by ADM, and lowered sensitivity to ADM.
Footnotes
Acknowledgment
This work was supported by Shiyan Science and Technology Bureau of Hubei Province in 2014 (No. 14Y58).
Disclosure Statement
There are no existing financial conflicts.