
Abstract
Cancer vaccines mostly aim to induce cytotoxic T lymphocytes (CTLs) against tumors. An appropriate adjuvant is of fundamental importance for inducing cellular immune response. Since the antigen in particulate form is substantially more immunogenic than soluble form antigen, it is beneficial to interact with antigen-presenting cells membrane to induce robust CD8+ T cell activation following vaccination. Based on previous research, we designed an adjuvant formulation by combining Astragalus saponins, cholesterol, and liposome to incorporate antigen into a particulate delivery system, so as to enhance cellular immune response. Meanwhile, angiogenesis contributes to tumor growth and metastasis, and basic fibroblast growth factor (bFGF) is involved in tumor angiogenesis. Therefore, using lipo-saponins adjuvant formulation and a human recombinant bFGF antigen protein, we tried to induce bFGF-specific CTL response to inhibit tumor angiogenesis to achieve antitumor activity. After five immunizations, the lipo-saponins/bFGF complex elicited robust antibody response and markedly higher amount of interferon-γ in BALB/c mice, resulting in superior antitumor activities. Decreased microvessel density in CD31 immunohistochemistry and the lysis of vascular endothelial cells by the T lymphocytes from the immunized mice indicated that the immunity inhibited the angiogenesis of tumors and further led to the inhibition of tumors. Our data suggest that the approach to construct adjuvant formulation between liposome and Astragalus saponins appeared highly desirable, and that Astragalus saponins may be utilized as a valuable additive for enhancing the effectiveness of vaccines and stimulating an appropriate immune response that can benefit tumor therapy.
Introduction
Multiple evidences suggest that immune cells play a prominent role in the control of malignancy. 1 –3 A protein-based vaccine without a potent adjuvant usually results in a weak immune response. Therefore, the use of appropriate adjuvants for the induction of an effective immune response in tumor immunotherapy is critical. An ideal adjuvant enhances immunogenicity of antigens and improves antigen delivery in antigen-presenting cells. 4,5
The classification of adjuvants includes cytokines, microbial-derived adjuvants, emulsions, polysaccharides, minerals, surface active compounds, nucleic acid adjuvants, and particulate antigen delivery systems. 4 New generations of nucleic acid-based adjuvant such as DNA adjuvant, although it has high immunogenicity than the traditional adjuvants, cannot be used clinically because of their unpleasant side effects. Hence, a vaccine adjuvant should ensure that the vaccine can initiate a potent immune response and minimize its side effects. 6
Saponins are steroid or triterpenoid glycosides, derived from some wild plants, bacteria, or lower marine animals. 7 Previous studies have shown that saponins can enhance macrophage phagocytosis, secretion of antibodies, and cytotoxic T lymphocyte (CTL) production against exogenous antigens. 8,9 A lot of research focuses on saponins of Quillaja saponaria, which can been wrapped by neutral phospholipids and cholesterol as an immunostimulating complex (ISCOM). 10 Antigens used in such ISCOM can effectively induce specific humoral and cellular immune responses. 11 But Quillaja saponaria saponins are only used in veterinary vaccines because of their high hemolytic activity. 12 Astragalus, a Chinese traditional herb, the largest genus in the family Leguminosae, its root represents a well-known diuretic, antiperspirant, and tonic drug in traditional Chinese medicine. Astragalus saponins can significantly enhance humoral and cellular immune responses in mice, with very low hemolytic activity in mice. 13 Because of this, we sought to examine whether the saponins from Astragalus is an appropriate vaccine adjuvant for tumor immunotherapy in the study.
Liposome, another extensively used adjuvant, is a vesicle formed by hydrophobic interaction of phospholipid bilayer. It can encapsulate antigen protein into its interior and deliver antigen to antigen-presenting cell. 14 But the problems with liposomes include inherent physical instability and lack of immunogenicity, one-way of increasing the immunogenicity of vaccines is to incorporate the antigen into a particulate delivery system. 15 Since the antigen in particulate form is substantially more immunogenic than soluble form antigen, it is beneficial to interact with antigen-presenting cells membrane to induce robust CD8+ T cell activation following vaccination. 16,17 Demana et al. 18 found that incorporating the adjuvant Quil A (saponins from Quillaja saponaria) into liposomes is highly desirable in terms of immunogenicity. The size of liposomes (with no Quil A) is usually above 100 nm in diameter (Fig. 1A). A spherical cage-like structure of ∼40 nm in diameter will be formed by the addition of 20% Quil A (Fig. 1B), the structure represents the classical ISCOM. With the addition of 70% Quil A, the structures are broken up into 10–12 nm small ring-like micelles (Fig. 1C). 18 The addition of Quil A disrupts the enclosed bilayer of liposomes to form regular particles. The more Quil A is added, the smaller the particles will become. Incubation with these particles can increase activation and proliferation of T cells. 18
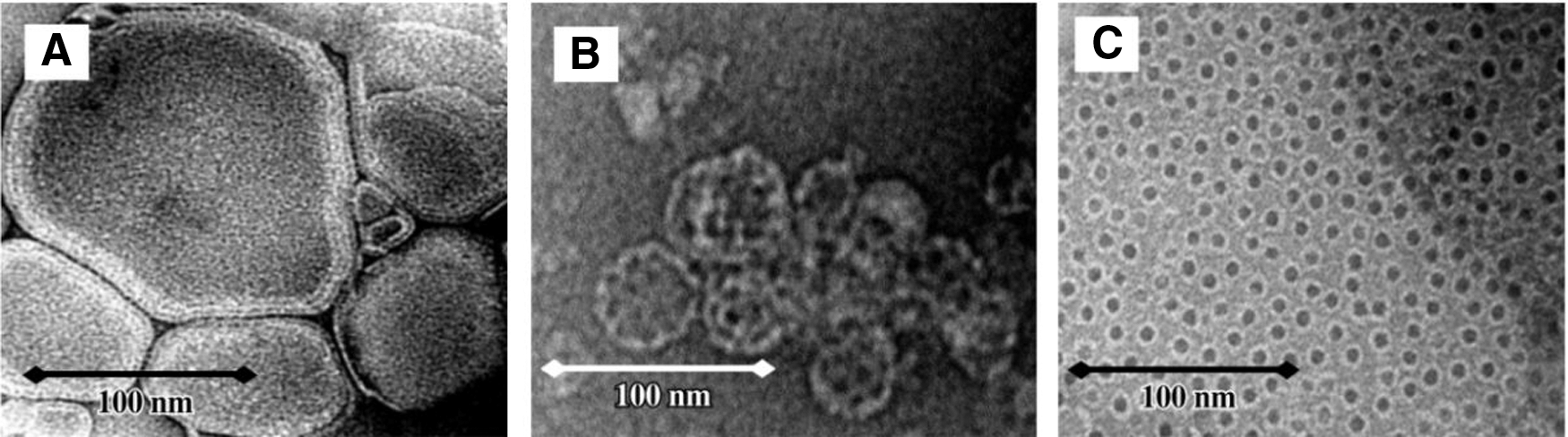
Transmission electron micrographs of formulations containing
The primary purpose of cancer vaccine is to induce persistent antitumor CTL response. 19,20 On this account, here we used a similar adjuvant formulation to improve antigen-specific CTLs response. The adjuvant formulation was prepared by the hydration method, 21,22 forming by Astragalus saponins, cholesterol, and liposome.
Angiogenesis plays a vital role in tumor growth, progression, and metastasis, tumor microvessel helps tumor cells to get metabolic requirements. 23 And antiangiogenesis has been proved to be a very effective strategy for tumor treatment by active immunity. 24,25 Basic fibroblast growth factor (bFGF), also known as bFGF, FGF-2, or FGF-β, is involved in tumor neovascularization and metastasis of cancer cells as an angiogenic factor. 26,27 The immunity targeting bFGF might be an effective strategy for cancer immunotherapy. However, autologous bFGF protein would be recognized as a self-molecule and induce no response due to immune self-tolerance. Therefore, here we used a human-derived recombinant bFGF protein as a xenogeneic antigen to immune mice, attempting to achieve antitumor activity through antiangiogenesis way.
Materials and Methods
Preparation of bFGF antigen protein
The amino acid sequence of human-derived recombinant bFGF antigen protein is MAAGSITTLP ALPEDGGSGA FPPGHFKDPK RLYCKNGGFF LRIHPDGRVD GVREKSDPHI KLQLQAEERG VVSIKGVCAN RYLAMKEDGR LLASKCVTDE CFFFERLESN NYNTYRSRKY TSWYVALKRT GQYKLGSKTG PGQKAILFLP MSAKS. The bFGF gene was inserted into prokaryotic expression vector plasmids (pQE30; Qiagen, USA) and recombinant plasmids were analyzed by DNA sequence analysis. Inclusion bodies of bFGF protein expressed in Escherichia coli. The protein with six tandem histidine residues was collected by Ni-chelating sepharose column (GE Healthcare Science), after the inclusion body was lysed under high pressure (APV 2000, Denmark).
Animals and cell line
Six-week-old female BALB/c mice were purchased from Slack experimental animal Co., Ltd. (Shanghai, China). CT26 cell (murine colon carcinoma cell) and MS1 cell (murine vascular endothelial cell) were purchased from American Type Culture Collection and respectively cultured in DMEM (Dulbecco's Modified Eagle's medium) Medium (with 10% fetal bovine serum or FBS) and RPMI-1640 Medium (with 20% FBS).
Preparation of adjuvant formulation
The adjuvant formulation was prepared by the hydration method. 21,22 In brief, liposomes were added to chloroform dissolved in cholesterol and evaporated to dryness at 37°C for 1 hour, then antigen protein was incorporated at 4°C, and finally mixed with saponin (dissolved in dimethyl sulfoxide) by an injector at room temperature (the saponin, cholesterol, and liposome was mixed at the ratio of 1:2:20 in weight).
Immunization
Mice were immunized by s.c. injection near axillas once a week for five consecutive weeks per mouse per injection with the following: (1) normal saline (NS Group); (2) saponins/bFGF: 10 μg bFGF protein, 5 μg saponins (YuanYe Biotechnology Co., Ltd., Shanghai, China) and 10 μg cholesterol; (3) lipo/bFGF: 10 μg bFGF protein, 10 μg cholesterol and 100 μg liposome (Liposome 2000; Invitrogen, Carlsbad, CA); (4) lipo-saponins/bFGF: 10 μg bFGF protein, 5 μg saponins, 10 μg cholesterol and 100 μg liposome. Blood samples were collected from the immunized mice on day 7 after each immunization. Serum was separated and stored at −70°C.
Tumor models
After five immunizations, the mice were challenged with 5 × 106 tumor cells s.c. in the right anterior axillary. Tumor dimensions were measured every 3 days by calipers and tumor volumes were calculated according to the following formula: width2 × length × 0.52.
In addition, to investigate the effect of the immunity on cancerometastasis, CT26 lung cancerometastasis models were established. Briefly, BALB/c mice were injected with 3 × 104 CT26/100 μL cells i.v. on day 7 after the last immunization. After 2 weeks, mice were sacrificed to count the cancer nodules on the surface of lung.
Enzyme-linked immunosorbent assay determination of anti-bFGF antibody
The level of bFGF-specific antibodies in the sera of the immunized mice was evaluated on day 7 after each immunization. Ninety-six-well micro titer plates (Falcon) were coated overnight at 4°C with 10 μg/mL bFGF in 50 mM carbonate buffer. After three washes with 0.1% Tween in phosphate-buffered saline (PBST), the plates were blocked with 5% skim milk in PBST for 1 hour at 37°C, followed by new washes. Serial dilutions of mice sera were added to the wells and incubated for 2 hours at 37°C. The wells were then washed and incubated with goat antimouse IgG antibodies conjugated with horseradish peroxidase (HRP; Sigma) for 1 hour at 37°C, washed again, and then incubated with substrate-chromogen solution for 20 minutes. The reactions were stopped by 0.5 M sulfuric acid and the absorbances were read at 405 nm with a BioRad microtiter plate reader.
Cytokine assay
The immunized mice were sacrificed and splenocytes were aseptically separated with lymphocyte separation medium. The splenocytes (3 × 106 cells in 200 μL, n = 5) were washed twice, resuspended in cell-culture medium, and then stimulated in vitro with 10 μg/mL of bFGF protein at 37°C for 48 hours. Supernatants were collected and the amounts of interferon-γ (IFN-γ) and interleukin (IL)-4 in the medium were measured using ELISA (enzyme-linked immunosorbent assay) Kits (Rockford, IL) according to the manufacturers' instructions.
CD31 immunohistochemistry
CD31 immunohistochemistry within tumor tissues was conducted using a monoclonal rabbit antimouse immunoglobulin to determine the intratumoral microvessel density. Frozen sections were fixed in cold acetone for 20 minutes. After two washes with PBS, the slides were incubated with 3% hydrogen peroxide to inactivate endogeneous peroxidase. After a new wash, they were blocked with rabbit sera at 37°C for 15 minutes, incubated with primary antibodies (1:400) against CD31 at 37°C for 2 hours, with secondary antibody for 40 minutes, and then with tertiary antibodies (HRP-labeled streptavidin) for 40 minutes. Diaminobenzidine was added as a chromogen. Tissue sections were stained under a microscope, rinsed in water, and then counterstained with hematoxylin. Microvessels were counted under a microscope.
CTL assay in vitro
CTL activity was measured using CytoTox 96 NonRa-dioactive Cytotoxicity Assay Kit (Promega, Madison, WI). The splenocytes (effector cells) from the immunized mice were cocultured with target cells (mouse microvascular endothelial cells MS1 and CT26 cancer cells, respectively) at different effector:target ratios. Lysis reaction was carried out for 10 hours at 37°C, after the reaction, 100 μL supernatant was removed to detect lactate dehydrogenase activity. Specific lysis was calculated with the absorbance at 490 nm as suggested by the manufacturer.
Statistical analysis
All data were analyzed using SPSS 22.0. The statistical significance of data was determined by Student's t-test and analysis of variance. Differences were considered significant when p < 0.05.
Results
Characterization of antibodies anti-bFGF
Sera from the immunized mice were collected and antibodies levels were assessed by ELISA. The optical density value in mice immunized with lipo/bFGF and lipo-saponins/bFGF were increased significantly, compared with saponins/bFGF and normal saline (N.S) mice (p < 0.01) (Fig. 2), but no great difference was observed between the two groups (lipo/bFGF and lipo-saponins/bFGF) (p = 0.22). Relatively, only very weak responses were induced by bFGF protein alone (results not shown).

Expression of antibodies anti-bFGF. Blood samples were collected on day 7 after the last immunization and sera were separated. Plates coated with bFGF (0.5 μg/well) antigen protein were incubated with serial dilutions of the sera. Antibodies expressions were determined by ELISA. (n = 10). bFGF, basic fibroblast growth factor; ELISA, enzyme-linked immunosorbent assay; N.S, normal saline; O.D, optical density value.
Characterization of cytokines
To evaluate the effect of the adjuvants on Th1 and Th2 cytokine response, the release of IFN-γ and IL-4 was assessed. Splenocytes from the immunized mice were stimulated in vitro with bFGF protein. The amounts of IL-4 and IFN-γ in the cell-culture medium were measured. As expected, lipo-saponins/bFGF showed its advantage in enhancing cellular immune responses, with a markedly higher amount of IFN-γ than the other groups (p < 0.01) (Fig. 3). As for the amount of IL-4, it increased in lipo-saponins/bFGF and lipo/bFGF mice, but the data were not statistically significant different (p = 0.31) (Fig. 3).
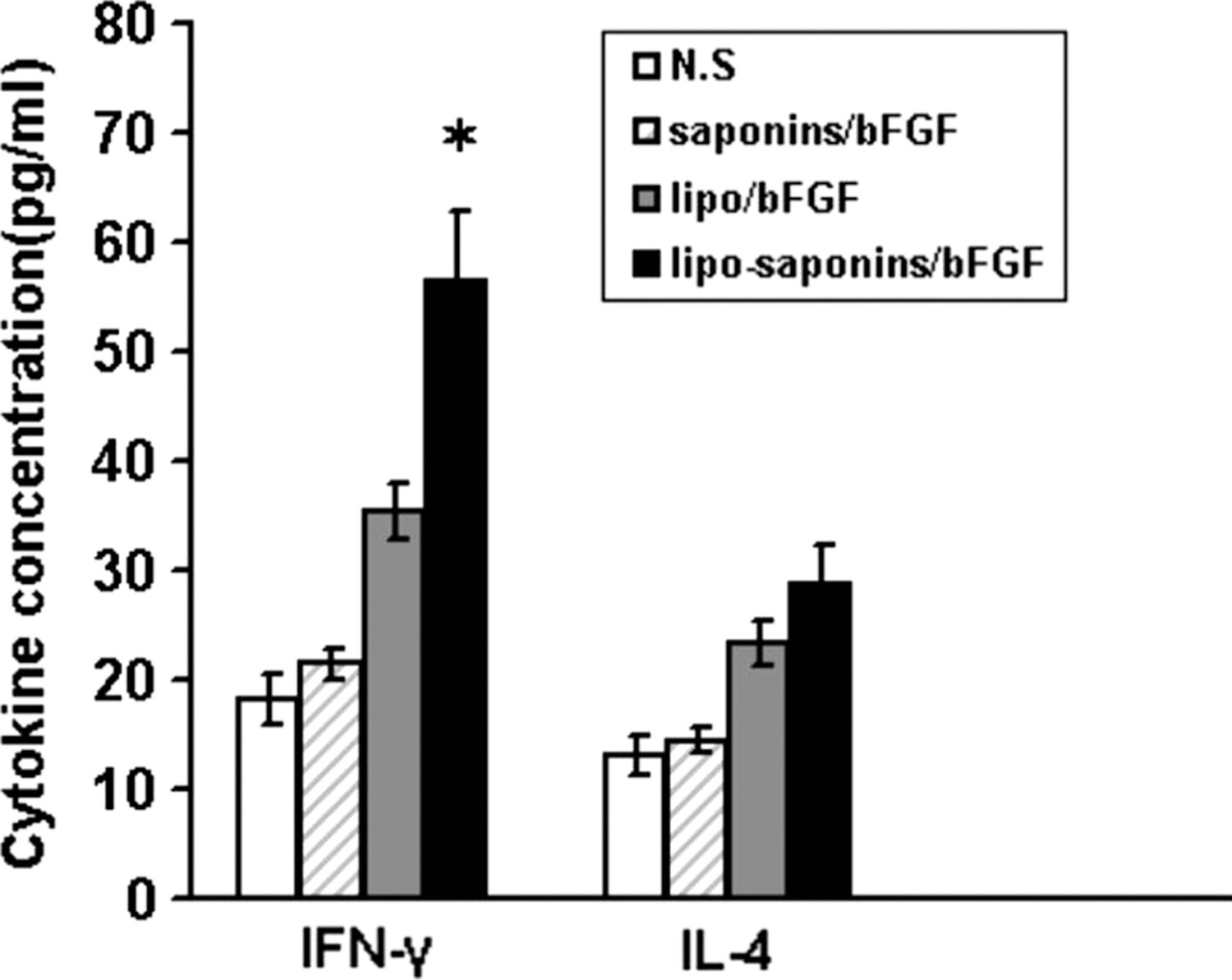
Cytokine release. After five immunizations, mice splenocytes (3 × 106 cells in 200 μL, n = 5) were separated and restimulated in vitro with 10 μg/mL of bFGF protein for 48 hours. The amounts of IL-4 and IFN-γ in cell-culture medium were measured using ELISA kits (n = 5). IFN-γ, interferon-γ; IL, interleukin-4. Asterisk (*) indicates a significant difference in amount of interferon-γ (p < 0.01) between lipo-saponins/bFGF group and other groups.
Induction of antitumor immunity
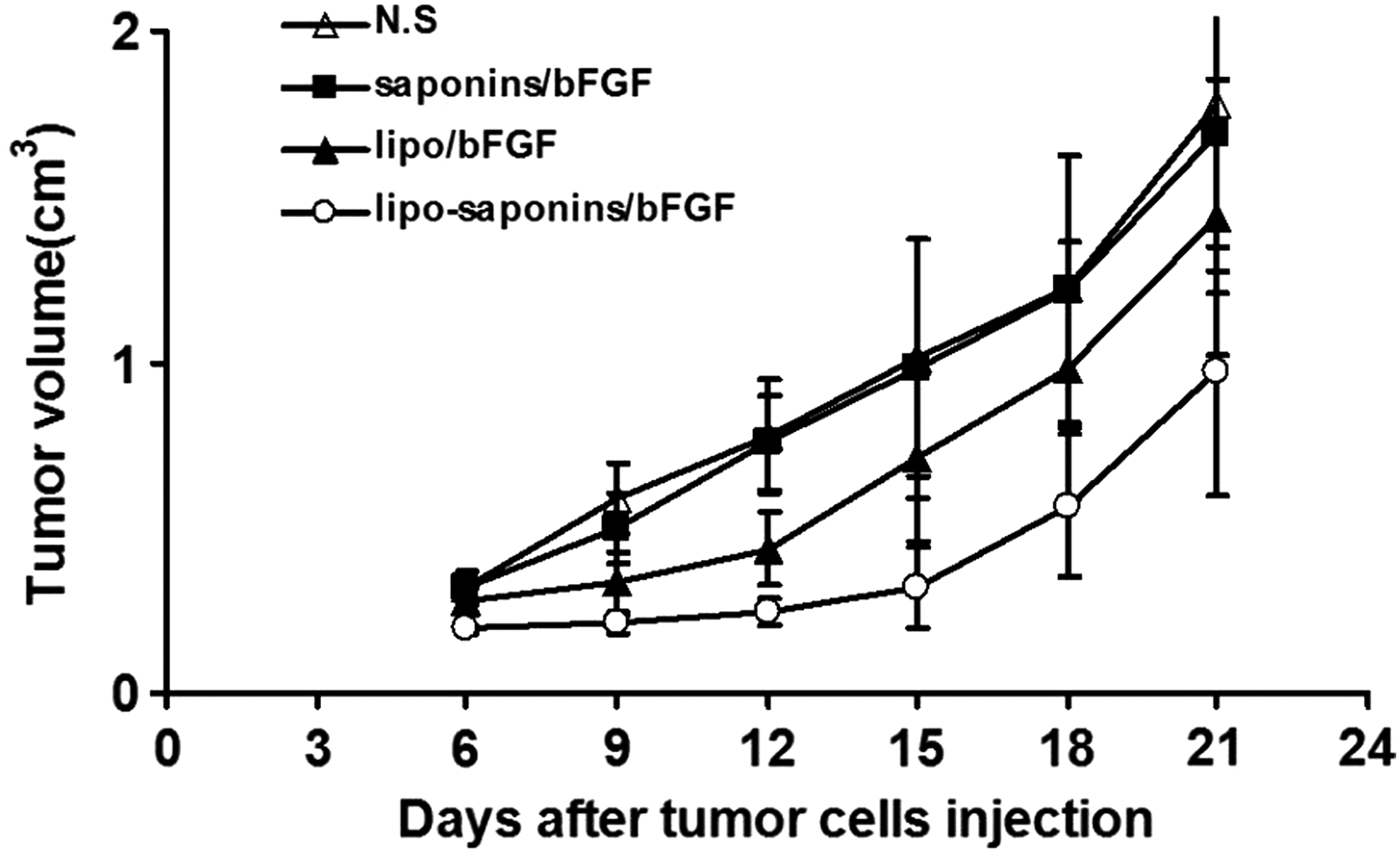
As shown in Figure 4, saponins/bFGF immunity hardly inhibited tumor growth, and lipo/bFGF had only a mild suppressive effect. More than other counterparts, lipo-saponins/bFGF induced a quite observable protective antitumor activity (p < 0.01). In the model of lung cancerometastasis, lipo-saponins/bFGF had a slight block on cancerometastasis, the number of lung surface nodules was reduced than that of N.S and saponins/bFGF mice (p = 0.046). Compared with lipo/bFGF group, the block was not significant (p = 0.37) (Fig. 5A, B), and hematoxylin and eosin staining confirmed the results by the metastatic colonies in the sections (Fig. 5C).
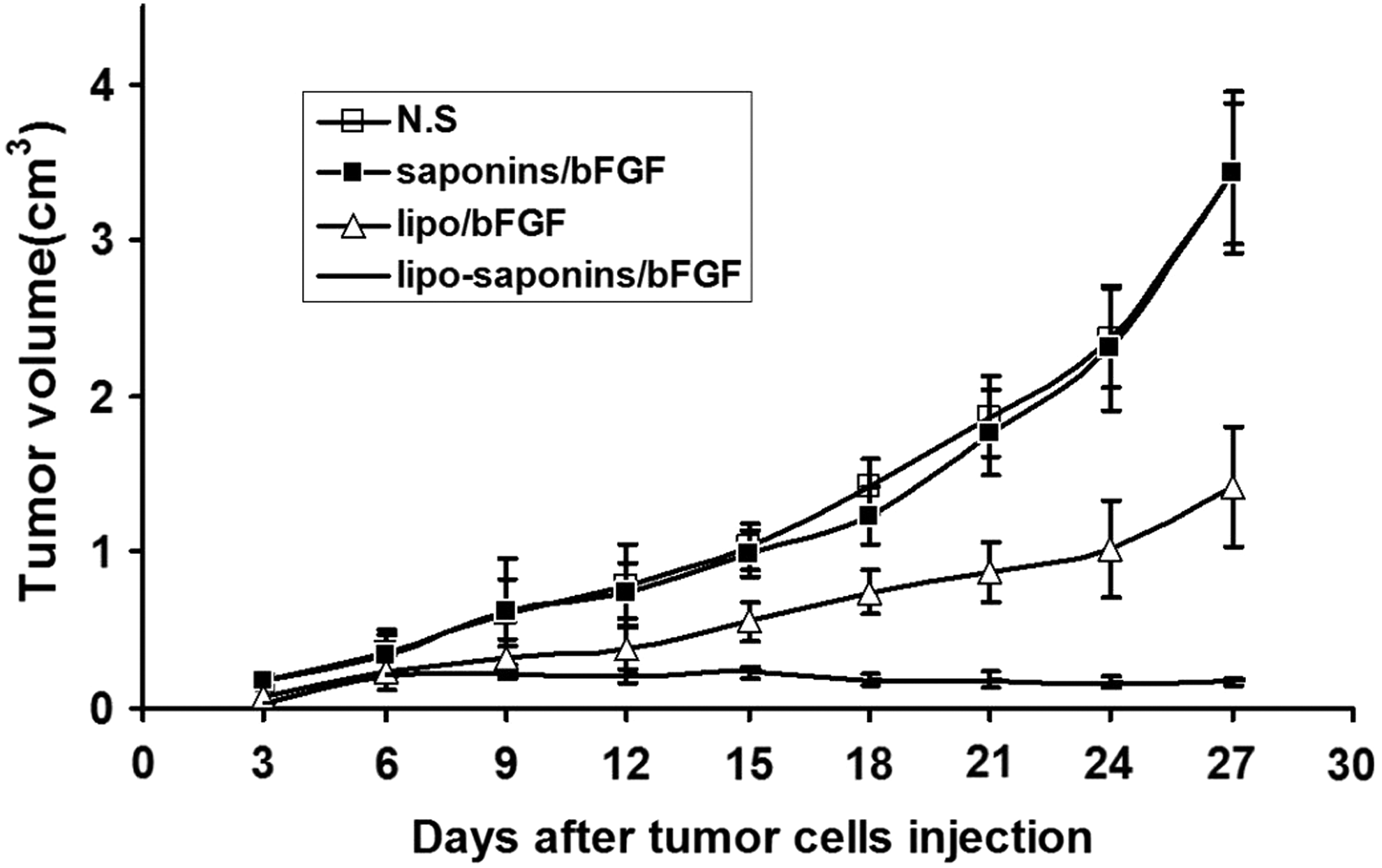
Induction of protective antitumor immunity. After five immunizations, BALB/c mice were challenged subcutaneously with CT26 cells (5 × 106). Tumor dimensions were measured every 3 days by calipers (n = 10).
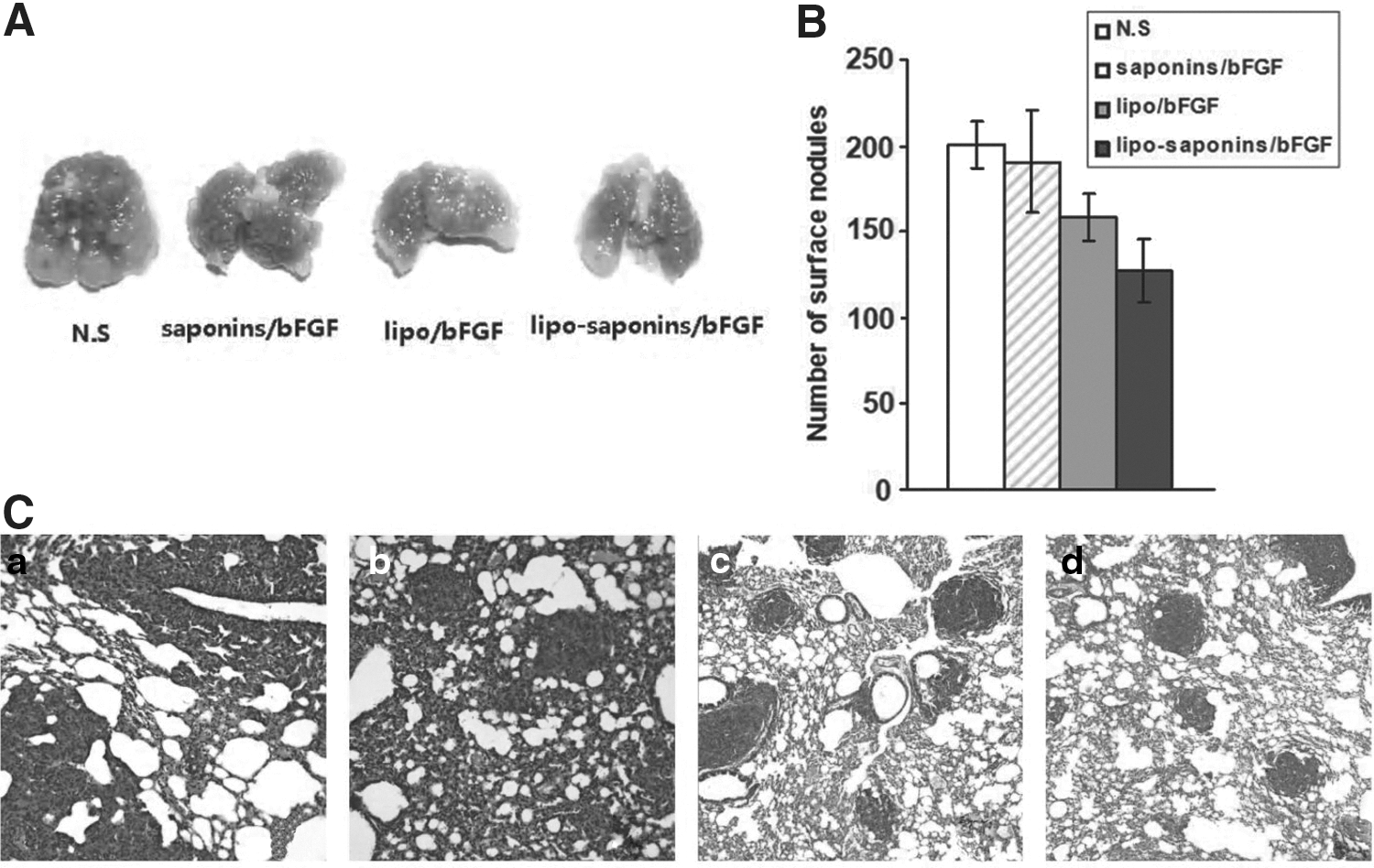
Induction of lung metastasis model. Mouse lung metastasis model was established by i.v injection with 3 × 104 CT26 cells. Mice were sacrificed on day 14 after cancer cells injection.
Inhibition of angiogenesis
The effects of these immunizations on tumor angiogenesis were detected by immunohistochemistry staining of frozen sections with antibodies reactive to CD31. The results showed that the immunity targeting bFGF restrained tumor angiogenesis in varying degrees in both lipo/bFGF mice and lipo-saponins/bFGF mice, microvessel density in lipo-saponins/bFGF mice was significantly decreased than that in lipo/bFGF mice (p < 0.05), and saponins/bFGF immunity did not seem to effect angiogenesis (Fig. 6).
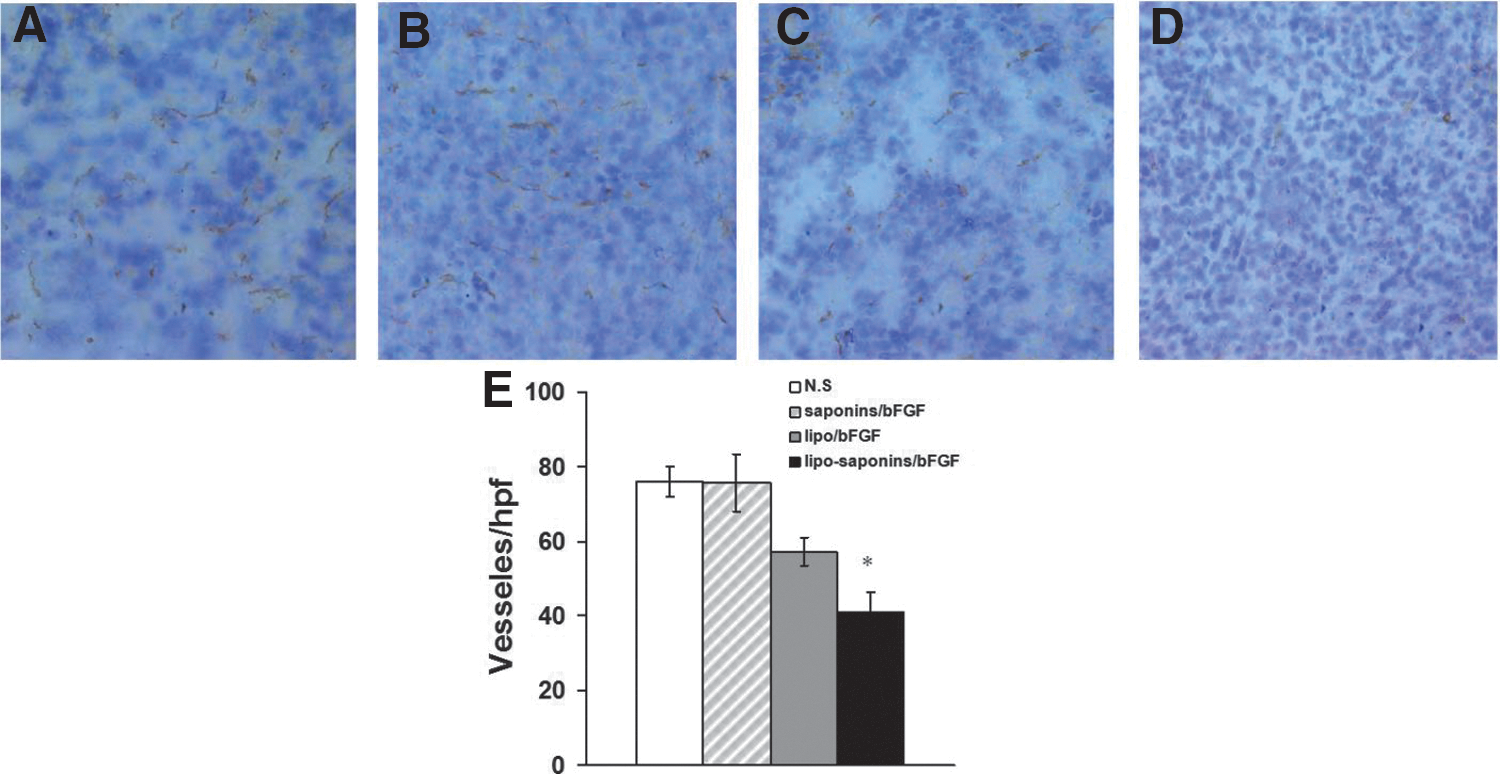
Inhibition of angiogenesis within tumors. Frozen sections of tumors from the animals were specifically stained for endothelial cells using a monoclonal rabbit antimouse immunoglobulin reactive to CD31. Microvessel density was determined by counting the number of microvessels per high power field in the sections.
Induction of adoptive immunotherapy
To further confirm the antitumor effect, adoptive immunotherapy was administered with activated spleen cells (2 × 107, i.v. injection) from the immunized mice. Next, the recipient mice were subcutaneously challenged with CT26 cancer cells (2 × 106) 24 hours after the adoptive immunity. The adoptive immunotherapy with the spleen cells of lipo-saponins/bFGF mice demonstrated the most inhibiting action on tumor growth, especially during the first few days (Fig. 7) (p < 0.01).
Induction of adoptive immunotherapy. Activated T lymphocytes from the immunized mice were adoptively and intravenously transferred into recipient mice. The recipient mice were subcutaneously challenged with CT26 cells 24 hours after the intravenous injection (n = 5).
CTL-mediated cytotoxicity in vitro
MS1 (mouse vascular endothelial cells) target cells and CT26 target cells were respectively cocultured with T lymphocytes from the immunized mice to test bFGF-specific cytotoxicity of T lymphocytes in vitro. The lymphocytes from both lipo/bFGF and lipo-saponins/bFGF mice showed CTL activity against MS1 cells (Fig. 8), and the activity by lipo-saponins/bFGF was superior (p < 0.01).
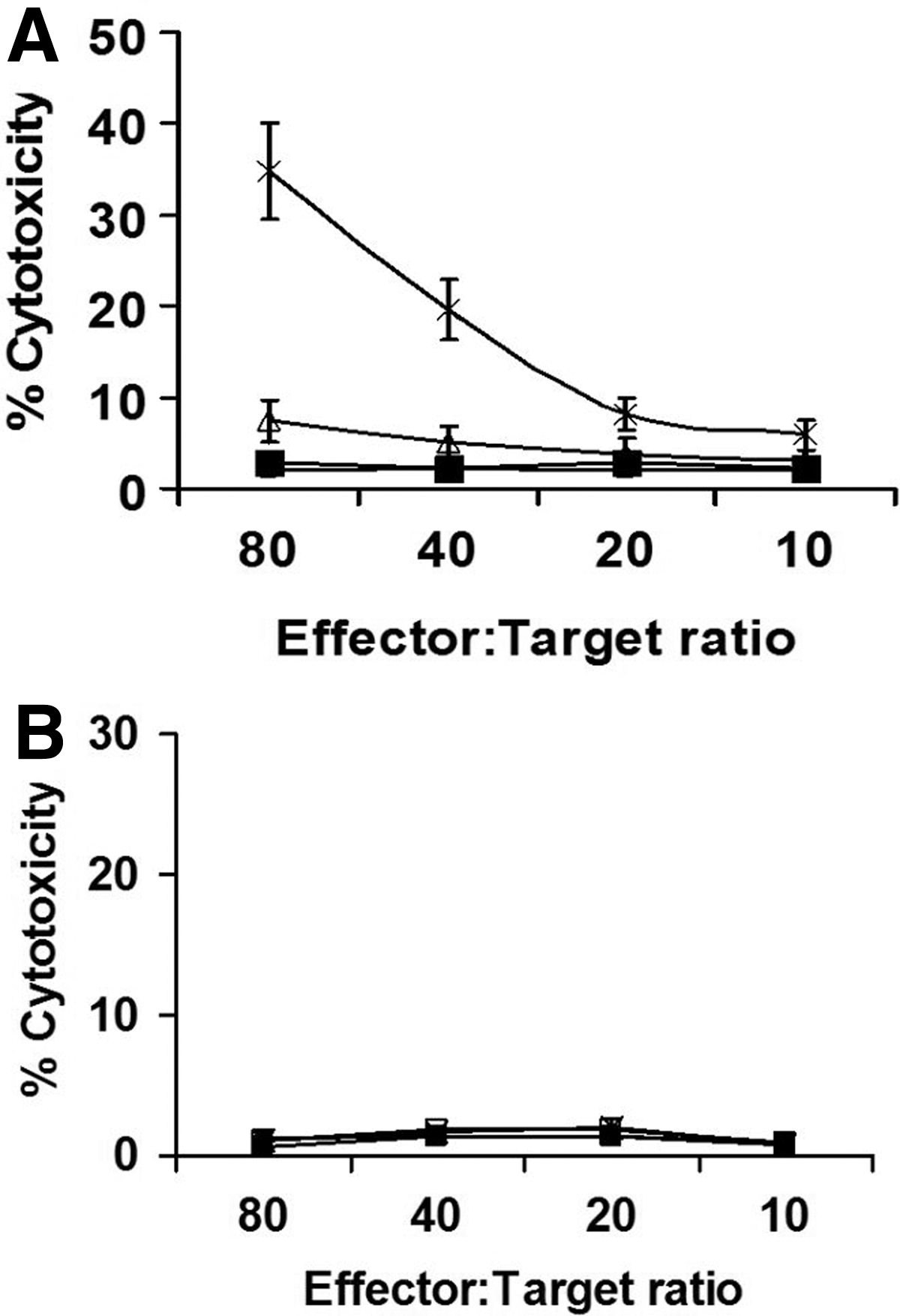
CTL assay in vitro. MS1 target cells and CT26 target cells were respectively cocultured with T lymphocytes from the mice immunized with N.S (□), saponins/bFGF (▪), lipo/bFGF (▵), and lipo-saponins/bFGF (*) at different effector:target ratios. After 10 hours of incubation at 37°C, the lactate dehydrogenase activity in supernatant was detected. T lymphocytes from lipo-saponin/bFGF mice showed the most apparent CTL cytotoxicity against MS1 cells
Side effect of the vaccination
No apparent side effects were observed (including animal behavior, feeding, water intake, weight loss, ruffling of fur, and mortality rate) after five immunizations. Furthermore, no pathologic change in internal organs was found by microscope observation. No obvious hemolytic activities were detected by red blood cell count and blood smear (data not shown).
Discussion
Adjuvant can significantly improve the immunogenicity of vaccine, and can make the immune system biased toward Th1 or Th2 type response. 28 Over the last few decades, a variety of adjuvants including mineral salts, aluminium adjuvant, microorganism adjuvants, emulsions, cytokines, polysaccharides, and nucleic acid-based adjuvants have been tested for vaccine design, most of them are not suitable for human use, with limitations or systemic toxicity. 4 Saponin is a natural glycoside of steroid or triterpene with the ability to modulate the cell-mediated immune system and has the advantage of low dose for adjuvant activity. 29,30 Their lipophilic acyl side chains were showed to have remarkable stimuli for CTL production against exogenous proteins. 31 The purpose of the cancer vaccine is to induce CTL response, 32 the unique capacity of saponins to stimulate the production of CTLs makes them ideal for therapeutic cancer vaccines. 33,34 In this study, we designed the liposome-saponins adjuvant formulation, taking into account the capacity of saponins to stimulate the production of CTLs and the ability of liposome to deliver the antigen to antigen-presenting cells. More importantly, the adjuvant formulation can incorporate antigen to form a particulate structure similar to ISCOM. 18 As expected, the adjuvant formulation elicited not only antibody response (Fig. 2), especially the bFGF-specific CTL response (Figs. 3 and 8).
Angiogenesis plays a crucial role in tumor progression. 23,35 Antiangiogenic therapy attempts to stop the formation of tumor microvessels and break up existing capillaries that feeds the cancerous mass. 23,36 bFGF is an important growth factor in angiogenesis. 26 It not only stimulates the proliferation of vascular endothelial cells, but also participates in tumor neovascularization. 37 Active antiangiogenic immunotherapy by targeting bFGF is effective in some previous studies. Hori et al. demonstrated that tumor growth was suppressed by bFGF-specific immunoneutralizing antibodies as a bFGF inhibitor to suppress tumor angiogenesis. 38 Using bFGF-activated fibroblasts as vaccine, Li et al. 39 induced an autoimmune response with potent antitumor activity. In this work, our liposome-saponins adjuvant formulation induced a more potent protective antitumor immunity (Fig. 4), tumor growth was significantly suppressed, and CD31 immunohistochemical staining displayed that angiogenesis within tumors was apparently inhibited by lipo-saponins/bFGF immunity (Fig. 6). These proved that the lipo-saponins/bFGF immunity truly had a marked impact on angiogenesis. As for cancerometastasis, our observations showed that lipo-saponins/bFGF vaccine brought only a slight block on the lung metastasis of cancer cells, with no great difference compared with lipo-saponins/bFGF vaccine (Fig. 5). It may be explained that inhibition of a single angiogenic factor pathway is unlikely to completely suppress tumor metastasis, because multiple angiogenic factors are involved. 40
Antitumor immunity mainly depends on the toxicity of T lymphocytes in some mouse models. 41,42 An immune response can be classified as Th1 or Th2 types characterized respectively by the production of cytokines like IFN-γ and IL-2, or IL-4, IL-5, IL-10, and IL-13. 43 Evaluation of IFN-γ and IL-4 responses in immunized mice revealed that lipo-saponins/bFGF mice showed a sharp increase in IFN-γ production than lipo/bFGF and saponins/bFGF mice (Fig. 3). The research of Demana et al. 18 suggest that the addition of saponins could profoundly affect the structure of the delivery system of liposomes and increase the immunogenicity to stimulate antigen specific lymphocyte proliferation and activation. Further, marked antitumor activity was observed in this group. More, the activity was confirmed by adoptive immunotherapy with the activated spleen cells from the corresponding mice (Fig. 7). In vitro, CTL assay showed that the CTLs leaded to the lysis of mouse microvascular endothelial cells (Fig. 8A). These findings suggested that the antitumor activity may be mostly attributed to bFGF-specific CTL response, which could suppress angiogenesis of tumor and hence resulted in the inhibition of tumor growth.
In conclusion, the novel approach to construct adjuvant formulations between different adjuvants of known stimulatory actions appeared to highly desirable, it could result in synergistic or complementary enhancement of immune responses to vaccines. The liposome-saponins adjuvant formulation was really efficient in stimulating both humoral and cellular immune responses, especially cellular immune. Therefore, we suggest that Astragalus saponins may be utilized as a valuable additive for enhancing the effectiveness of vaccines and stimulating an appropriate immune response that can benefit tumor therapy.
Footnotes
Acknowledgments
The work mainly performed in Gansu University of Chinese Medicine, Lanzhou, Gansu, China. This work was supported by the Natural Science Foundation of Gansu Science and Technology Department (Grant No. 1208RJZA221), National Natural Science Foundation of China (Grant No. 81360345), and Opening Foundation of Dunhuang Medicine and Transformation Key Laboratory of Ministry of Education (Grant No. DHYX17-07).
Disclosure Statement
The authors report no conflicts of interest. The authors are responsible for the content and writing of the article.