
Abstract
Samsum ant venom (SAV) is a rich repertoire of natural compounds with tremendous pharmacological properties. The present work explores its antineoplastic activity in different cell lines followed by its confirmation in vivo. The cell lines, HepG2, MCF-7, and LoVo showed the differential dose-dependent antineoplastic effect with an increased level of significant cytokines, including Interleukin (IL)-1β, IL-6, and IL-8 and transcription factor, Nuclear factor-kappa B (NF-κB). However, the venom was more effective on HepG2 and MCF-7 cells than LoVo cells. Furthermore, the extract was administered to four groups (n = 8) of rats. Group I was taken as a control without any treatment, whereas group II received CCl4 (1 mL/kg) for induction of mild hepatoma. Group III was given 100 μg/kg of SAV twice a week for 1 month. Group IV was pretreated with the CCl4 (like group II) followed by dosing with SAV (100 μg/kg) for 2 months as per the authors' prestandardized dosing schedule. Intriguingly, the rats of group IV demonstrated significant decrease in key cytokines, IL-1β and IL-6, as well as the transcription factors, including Tumor Necrosis Factor-alpha (TNF-α), NF-κB, and Inhibitor-kappa B (I-κB) as compared with group II. Furthermore, increase in IL-10 and First apoptosis signal (FAS) in the same group confirmed that SAV induces apoptosis at the given dose through immunomodulation leading to enhanced tumor killing in vivo. Hence, SAV has an excellent antineoplastic activity that can be directly used to treat certain types of cancer. Moreover, study of its ingredients can pave ways to design novel anticancer drugs. However, further in-depth investigation is required before its clinical trials.
Introduction
Ants are considered as the chemical factories as their venom may contain over 75 different types of chemical compounds. 1 –3 Pachycondyla is a genus having a large group of ants in which most of them are geographically distributed in the tropical and subtropical regions. Among them, the Samsum ant (Pachycondyla sennaarensis) is one of the most prominent members of the genus. The ant has been held responsible for the induction of anaphylaxis around the world. 4 The venom from these social insects results in mild local reactions, including acute pain, inflammation, itching, and irritability in humans. However, some severe allergic reactions have also been linked to their venom depending on the dose of the venom and individual's hypersensitivity toward the same. 5
Nevertheless, lots of literature entails that some of the bioactive components having various structural repertoire present in the venom of many animals/insects have tremendous therapeutic potentials against the different forms of disease and health crisis. 6 –11 Moreover, many of their components have shown high specificity and potency to many established molecular targets. 12 The aqueous solutions of the venoms have been reported to have various enzymatic and nonenzymatic proteins, free amino acids, and small biologically active compounds, such as histamine, 5-hydroxytryptamine, acetylcholine, norepinephrine, and dopamine. 13 Among many others, the samsum ant venom (SAV) has many pharmacological properties, such as regulation of local inflammation, pain, triggering an immune response, and enhancing the healing process of the damaged tissues especially in the liver. 14,15 Interestingly, this ant venom has been documented to have strong antineoplastic effect against the breast cancer cells in a dose- and time-dependent manner without affecting the neighboring healthy cells. 16,17
Despite significant scientific advancements in cancer management, cancer has been one of the leading causes of clinical deaths after cardiovascular diseases globally. Today, the traditional cancer treatment is based on chemotherapy, radiotherapy, and both as combined modalities. All of these treatment strategies are either effective against the particular type of cancer until a particular time, or their serious side effects force discontinuation of the chemotherapy. 18,19 Earlier studies have established the antioxidant and immune-boosting effects of SAV in vitro and in vivo. 16,20
The present study is aimed to confirm if these properties of SAV attribute in antineoplastic activity through immunomodulation against the cell lines of three primary forms of breast, liver, and colon cancers, such as MCF-7, HepG2, and LoVo. Also, the authors were interested in investigating the potential anticancer activity of SAV in chemical-induced hepatoma in rats.
Materials and Methods
Chemicals
All the chemicals and general reagents, including the cell culture media were purchased either from Sigma-Aldrich Chemical Company, USA, or Thermo Scientific (Gibco), USA. The rest of all other chemicals used were of analytical grade bought from BDH (United Kingdom) and Merck (Germany) brands.
Collection and preparation of the SAV
Colonies of samsum ants (including around 2000–2500 workers, with a brood of all stages and multiple queens) were collected from Al Ehsaa Governorate, East Riyadh (Kingdom of Saudi Arabia). These collected nests were kept in the insectarium of the Department of Zoology, College of Sciences, King Saud University (Riyadh). The ants were housed in specially designed plastic nest bottles with the sufficiently large surface area (45 × 30 × 18 cm) until venom extraction. The sting apparatus was removed by grabbing with the help of forceps at the last segment of the abdomen followed by their detachment gently. The venom was collected by pinching their venom gland, and the venom was stored in a small glass tube. 20
The samples were homogenized mildly followed by centrifugation at 1500 rpm for at least 2 min. 21 Moreover, the supernatant thus received was kept at −20°C until their treatment with the cell lines.
Cell culture and treatment
In the present study, there were three most common forms of human cancer cell lines, breast cancer (MCF-7) cells, liver cancer (HepG2) cells, and colon cancer (LoVo) cells. All of them were purchased from ATCC, USA. The cells were treated with SAV at different concentrations (0.1, 1, 10, 100, and 1000 ng/mL). All the cells were cultured separately with standard protocols with medium containing (Dulbecco's modified Eagle's medium [DMEM]), 10% fetal bovine serum, 100 U/mL penicillin, as well as 100 μg/mL streptomycin at 37°C with 5% CO2, 95% air, and complete humidity. Once the cells reached 90% confluence, they were detached using 0.05% trypsin/ethylenediaminetetraacetic acid and counted using Trypan Blue staining by hemocytometer.
Animal treatment
For confirmation of cell line-based in vitro studies, in vivo study was also designed in parallel. The experimentation on animals under the study was approved by the Animal Ethics Committee of the Zoology Department in College of Science at King Saud University, Riyadh, KSA. CCl4 was chosen to induce liver cirrhosis and mild hepatoma in the rats. 22,23 For this, 32 healthy Swiss albino rats (adult, 150 ± 20 g) were purchased from Central Animal House (College of Pharmacy, KSU, Riyadh). They were housed in the large cages under ethically approved controlled conditions (temperature at 25°C ± 3°C, 12-h day–12-h night cycle and clean water with pellet diet ad libitum).
They were divided into four groups taking eight rats in each group (n = 8). The group I was negative control group without any treatment. Group II received a single dose of 1 mL/kg CCl4 in corn oil (1:1 volume) followed by two additional half of the same dose once a week through an intraperitoneal (IP) injection for induction of hepatoma. The occurrence of hepatoma was confirmed by assessment of liver and kidney function tests, and histopathology (data not shown). Group III was administered with 1 mL of 100 μg/kg of SAV twice a week for 1 month by IP route. Group IV was pretreated with the CCl4 (1 mL/kg) like group II. After a month, the rats of this group were administered IP with 1 mL of 100 μg/kg SAV twice a week for 2 months as per their prestandardized dosing schedule. 9,20,22,23
After completion of the treatment, the animals were sacrificed by cervical dislocation on the next day to the last dose. Their liver was stored in a suitable quantity of RNA stabilization reagent (RNAlater, Qiagen, Germany) at −80°C for real-time polymerase chain reaction (RT-PCR) and ELISA analysis. Besides, their serum was collected from freshly drawn blood of the animals also stored for analysis.
Ethical approval
The current study did not involve any endangered or protected species. Regarding the experimental animals, all procedures were conducted by the standard protocols outlined in the guidelines for the care and use of laboratory animals by the Committee for the Purpose of Control and Supervision of Experiments on Animals and the National Institutes of Health, USA. Also, the study protocol (care and handling of experimental animals) was approved by the Animal Ethics Committee of the Department of Zoology in the College of Science at King Saud University, Riyadh.
MTT assay for cell viability
MTT assay in the present study was performed after 24 h of treatment of SAV with the cell lines in a 96-well culture plate with the standard protocol. MTT solution was prepared at 1 mg/mL in phosphate buffered saline (PBS) that was filtered through a 0.2 μm filter. Then, 50 μL of MTT plus 200 μL of DMEM (without Phenol Red) were added to each well, except the cell-free blank wells. The cells were incubated for 3 h at 37°C in a CO2 incubator (Labmed, Korea) with the maintained condition of 5% CO2, 95% air, and complete humidity. After 3 h, the MTT solution was removed and replaced with 200 μL of DMSO and 25 μL Sorenson's glycine buffer. The plate was further incubated for 4 min at room temperature, and a plate reader determined the optical density of these wells at a wavelength of 570 nm.
RNA extraction and cDNA synthesis
Total RNA from all the three cell lines, MCF-7, HepG2, and LoVo, were collected in the RNA stabilization reagent. The liver sample of treated animals was also stored in the same reagent. From those, RNA was isolated using the RNeasy Mini Kit (QIAGEN, Germany). The isolation was conducted according to the manufacturer's instructions and quantified by measuring the absorbance at 260 nm, and RNA quality was estimated by measuring the 260/280 ratios. The cDNA synthesis was performed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems®, USA) according to the manufacturer's instructions.
Around 1.5 μg of the total RNA from each sample was added to a mixture of 2.0 μL of 10 × reverse transcriptase buffer, 0.8 μL of 25 × dNTP mix (l00 mM), 2.0 μL of 10 × reverse transcriptase random primers, 1.0 μL of MultiScribe reverse transcriptase, and 3.2 μL of nuclease-free water. 24 Thus, the final reaction mixture was kept at 25°C for 10 min that was heated to 37°C for 120 min, and then 85°C for 5 sec followed by their cooling to 4°C.
Quantification of mRNA expression of Tumor Necrosis Factor-alpha, interleukin-1, IL-6, and IL-8 in the cell lines by RT-PCR
The quantitative analysis of mRNA expression of the target genes was executed by RT-PCR. 20,24 For this, cDNA from the mentioned preparation was subjected to PCR amplification using 96-well optical reaction plates in the ABI Prism 7500 System (Applied Biosystems). The 25-μL reaction mixture had 0.1 μL of 10 μM forward primer and 0.1 μL of 10 μM reverse primer (40 μM final concentration of each primer) with 12.5 μL of SYBR Green Universal Mastermix, 11.05 μL of nuclease-free water, and 1.25 μL of cDNA sample. All the primers used in this procedure were chosen from pubmed.com online service. Data were analyzed using the relative gene expression method as per Applied Biosystems User Bulletin No. 2. Data were presented as the fold change in gene expression normalized to the endogenous reference gene and relative to a calibrator.
Measurement of key immunological factors in the animal samples by ELISA and RT-PCR
The serum samples from the treated animals were subjected to analysis of many key cytokines interleukin (IL)-1, IL-6, and IL-10 by the commercially available kits (Abcam, United Kingdom, and RayBiotech, USA). Furthermore, the level of expression of NF-κB, IκB, Tumor Necrosis Factor-alpha (TNF-α), and first apoptosis signal (FAS) was measured by RT-PCR gene expression in the liver samples.
Statistical analysis
All data of the experiments were analyzed by statistical analysis software GraphPad Prism. The post-hoc analysis was conducted by one-way ANOVA with Tukey's test for significance selecting p-value <0.05 under the software. In the figures, the asterisks (*) indicates significantly different values from the control (group I), whereas # indicates significantly different values from group II.
Results
Cell culture
The purpose of these in vitro experiments conducted on the cancer cells was to assess the effectiveness of SAV in the inhibition of tumor cell proliferation and its role in inflammation involved with them. For this, MTT assay of the treated cells, as well as the measurement of expression of critical immune factors, such as TNF-α, IL-1β, IL-6, and IL-8 gene expression were conducted. The efficacy of SAV was tested on three cell lines: breast cancer (MCF-7) cells, liver cancer (HepG2) cells, and colon cancer (LoVo) cells.
Effect of SAV on cellular proliferation
SAV was used in five different concentrations (0.1, 1, 10, 100, and 1000 ng/mL) for MTT assay in the established cancer cells for the assessment of inhibition of cellular proliferation. The extract demonstrated the dose-dependent inhibitory effect on their cellular proliferation; however, the statistical significance was observed in doses of 100 and 1000 ng/mL. In the present study, they chose 100 ng/mL as the standard dose for all further studies.
The results showed that SAV at the dose of 100 ng/mL was capable of inhibiting the growth of MCF-7 and HepG2 by 57.1% and 50.6%, respectively, but it was only 35.8% in the case of LoVo against their control. Although the ant extract demonstrated its efficacy in the range of 1–100 ng/mL in MCF-7 and HepG2, except the LoVo showed 1000 ng/mL as a statistically significant dose compared with the control. Hence, the extract was more effective against MCF-7 and HepG2 cell lines as compared with the LoVo (Fig. 1).
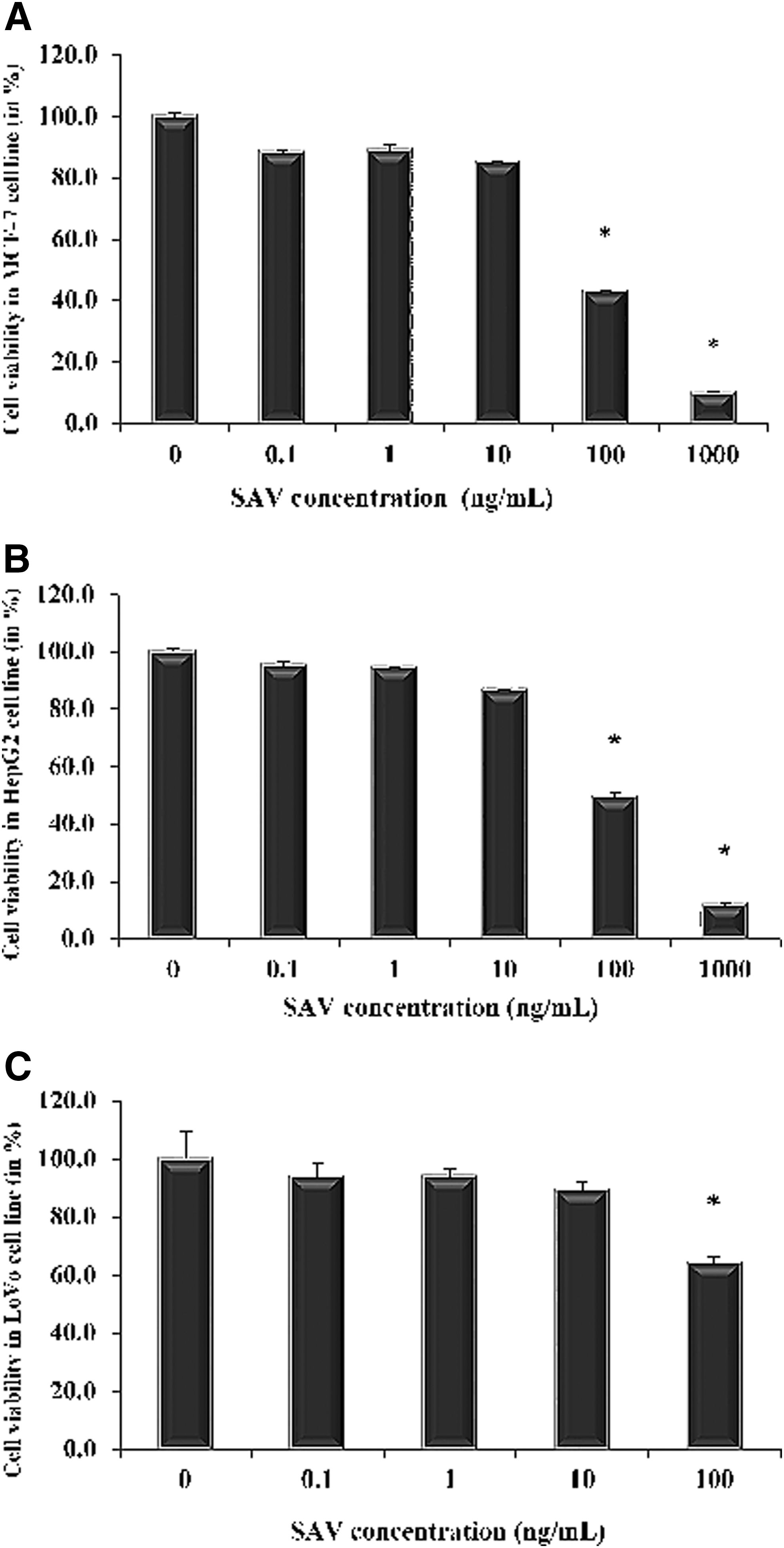
MTT assay for the anticancer effects of SAV against different cancer cell lines. MTT assay for the anticancer effects of SAV against different cancer cell lines
Effect of SAV treatment on the critical cytokines in the cell lines by RT-PCR
After treatment of the cells with SAV (dose of 100 ng/mL), their RNA was isolated and subjected to measurement of essential cytokines, IL-1, IL-6, IL-8, and TNF-α, taking GAPDH as the reference gene in all the experiments.
IL-1β
After the treatment of all three cell lines with SAV, the level of IL-1 was found to be increased by 16.1% and 1166.3% in MCF-7 and HepG2, whereas LoVo showed a decrease in its level by 25.3% as compared with the control (Fig. 2).
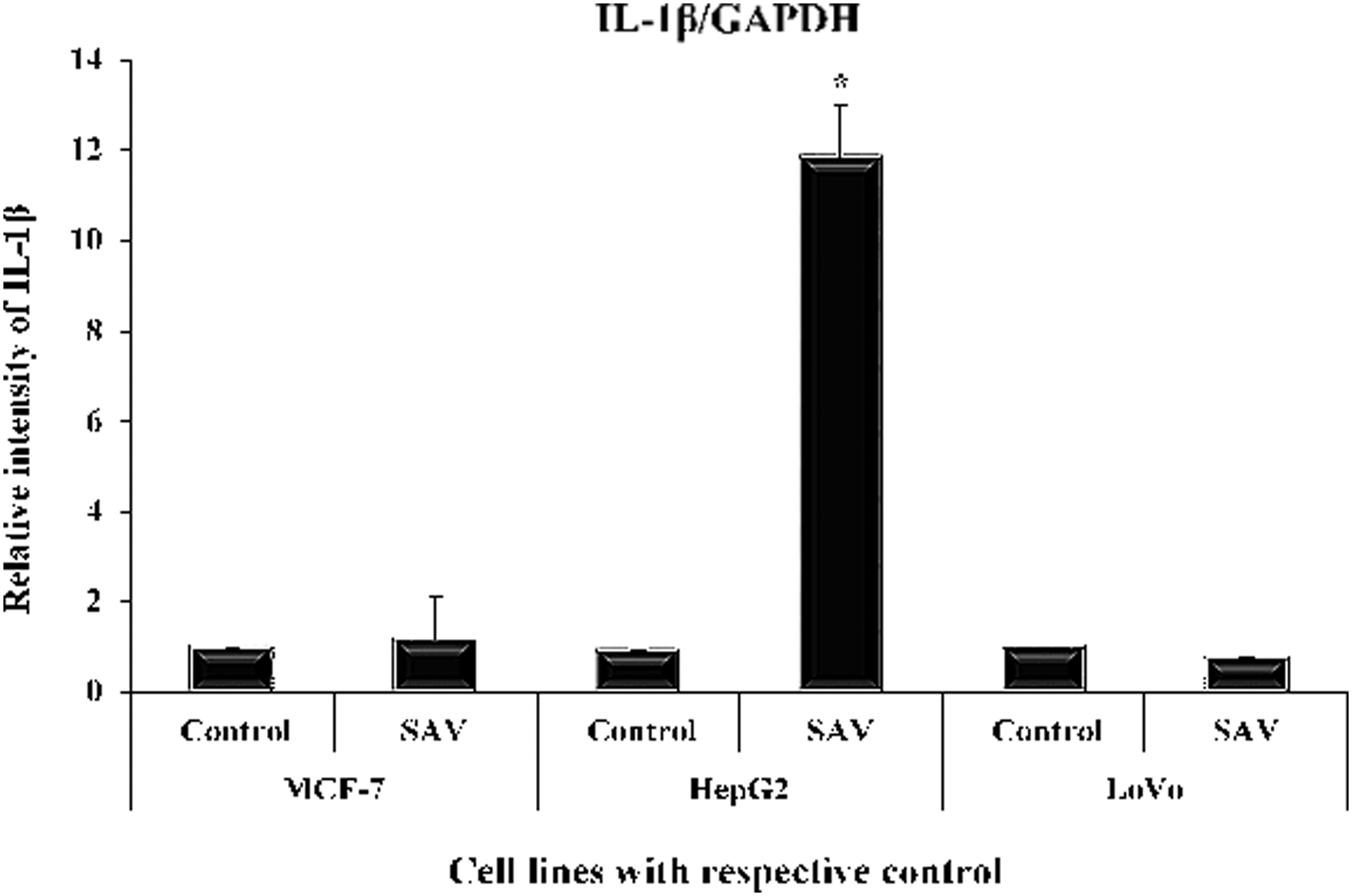
Showing level of Interleukin-1β in MCF-7, HepG2, and LoVo cells. Showing level of IL-1β in MCF-7, HepG2, and LoVo cells (in nanogram per milliliter of the supernatant of the cultured cells) by RT-PCR. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control. RT-PCR, real-time polymerase chain reaction; SEM, standard error of the mean.
IL-6
The treatment of SAV with MCF-7, HepG2, and LoVo cell lines caused an elevation in its level by 112.7%, 1546%, and 30% with respect to their controls (Fig. 3).
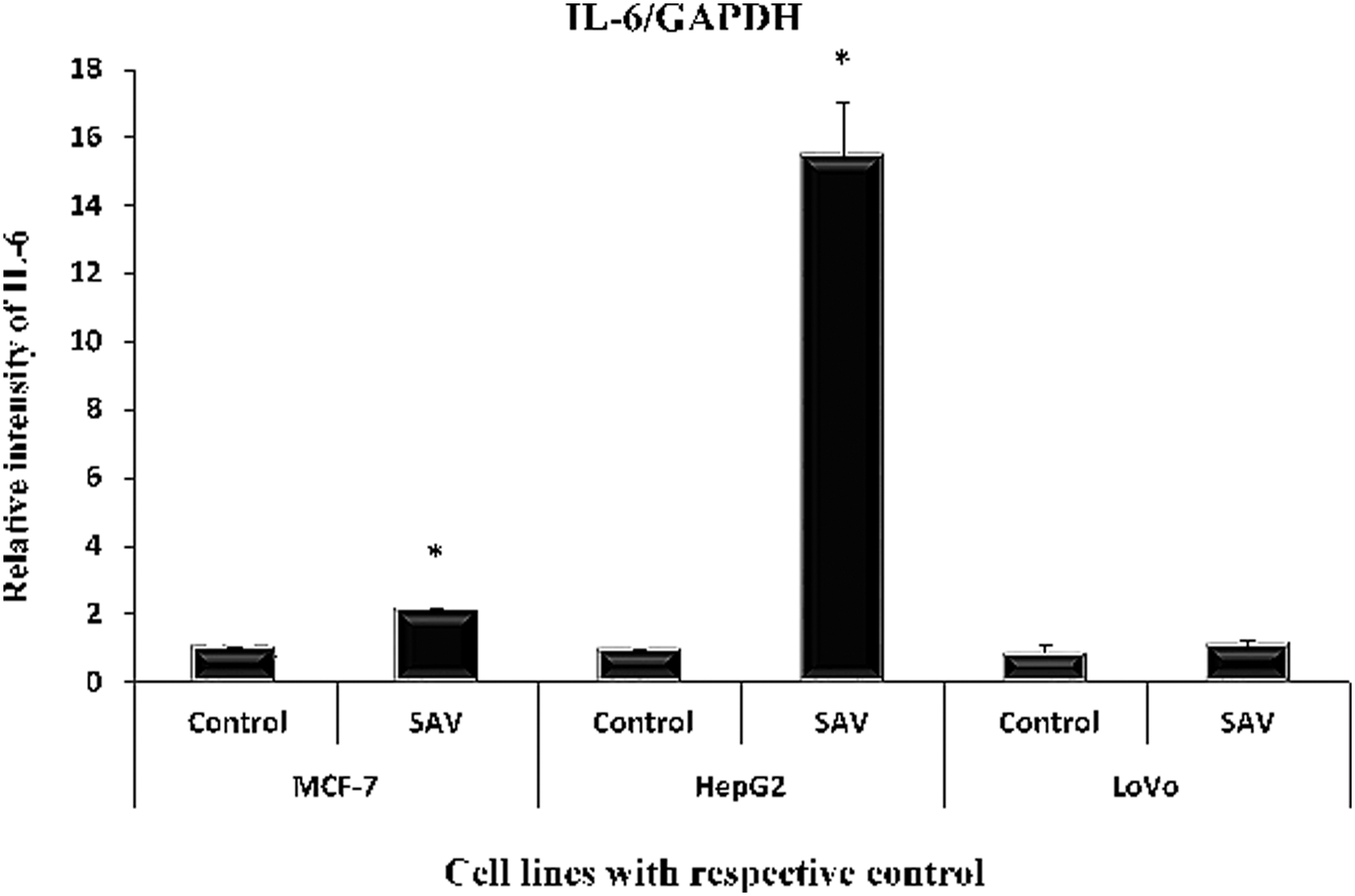
Showing level of IL-6 in the three cell lines. Showing level of IL-6 in the three cell lines (in nanogram per milliliter of the supernatant of the cultured cells) by RT-PCR. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control. SEM, standard error of the mean.
IL-8
The ant venom extract led to increasing its level by 755.4%, 1040.1%, and 12.7% in MCF-7, HepG2, and LoVo cell lines, respectively, in comparison to their control (Fig. 4).
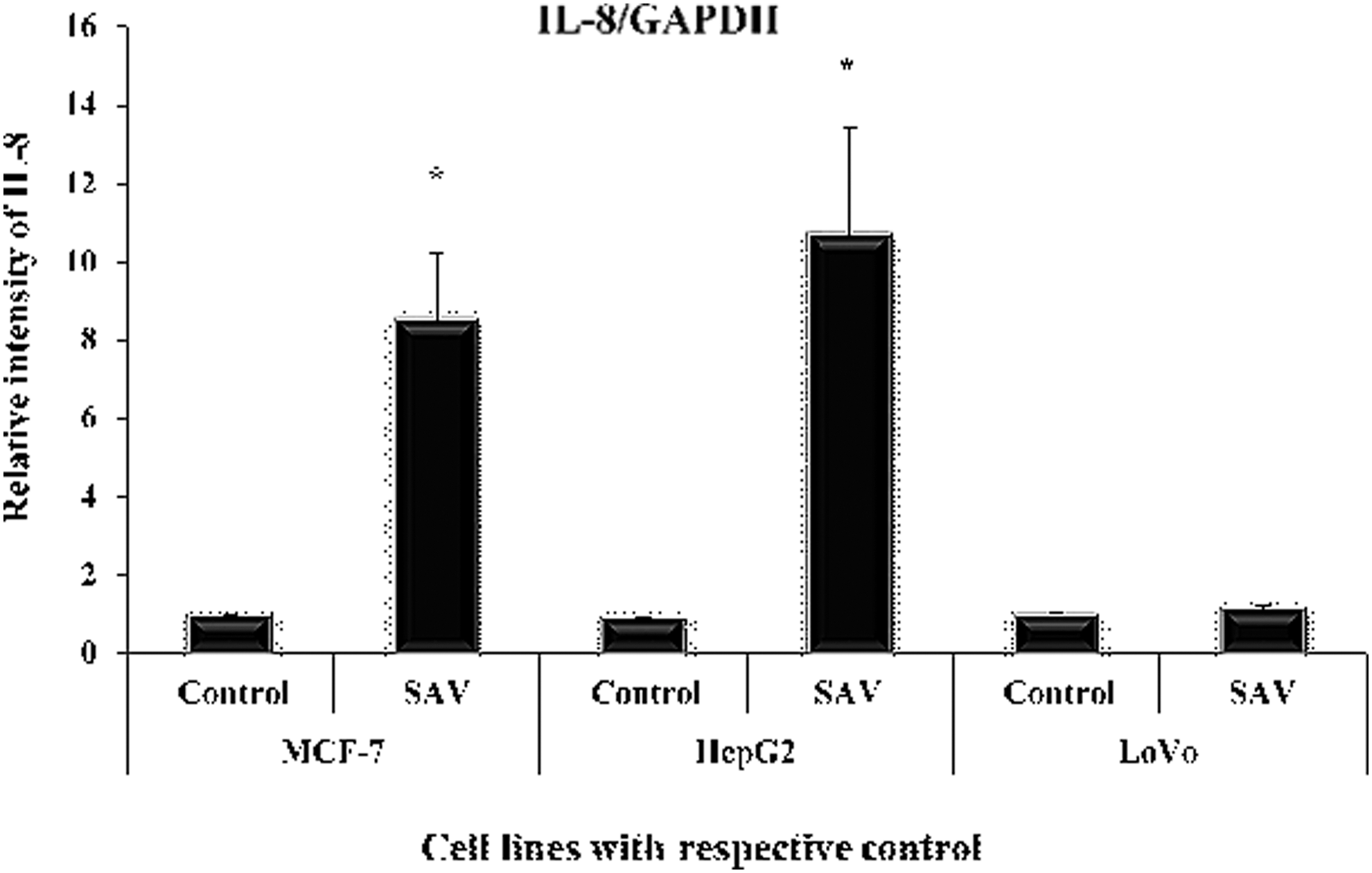
Showing level of IL-8 in the three cell lines. Showing level of IL-8 in the three cell lines (in nanogram per milliliter of the supernatant of the cultured cells). All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control. SEM, standard error of the mean.
TNF-α
The treatment with SAV increased its level by 201.4%, 813.6%, and 16.4% in the cell lines, MCF-7, HepG2, and LoVo, in the sequence as compared with their respective controls (Fig. 5).

Showing level of Tumor Necrosis Factor-alpha in the three cell lines. Showing level of TNF-α in the three cell lines (in nanogram per milliliter of the supernatant of the cultured cells) by RT-PCR. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control. SEM, standard error of the mean.
Animal studies
All the serum samples were subjected to measurement of key cytokines by the commercially available ELISA Kits to investigate if any correlation exists between their in vitro and in vivo findings.
IL-1β
In the present study, group II demonstrated an increase in its level by 116.73% with respect to the control, group I. However, group III and group IV showed a decrease in its level by 42.36% and 48.11%, respectively, as compared with group II (Fig. 6).
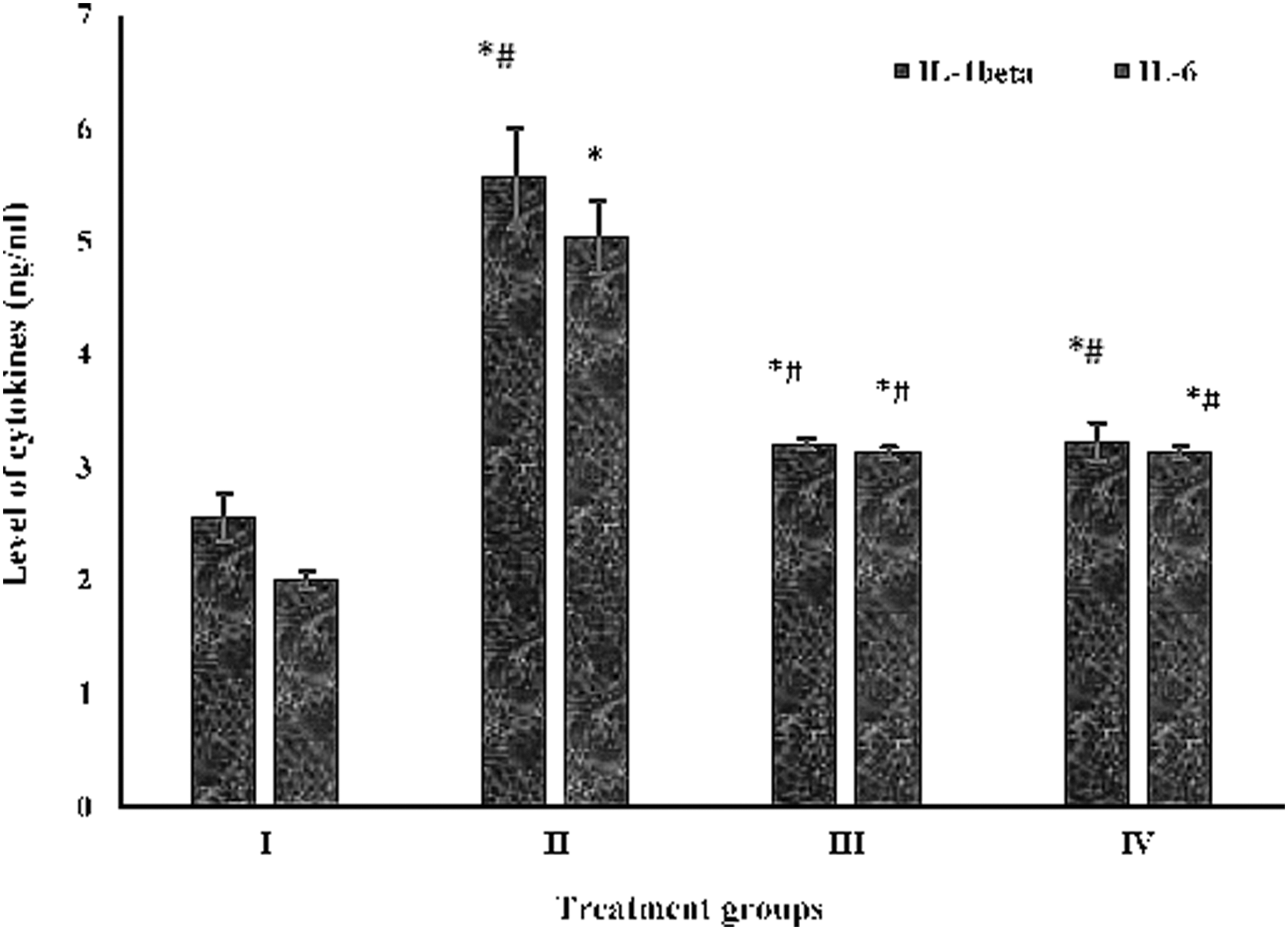
Showing level of IL-1β and 6 in the serum samples. Showing level of IL-1β and 6 (in nanogram per milliliter of the supernatant of the serum) by ELISA analysis. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control (group I). #Significantly different values from group II. SEM, standard error of the mean.
IL-6
In the present in vivo analysis, group II exhibited an enhancement in its level by 60.19% in comparison to group I, whereas groups III and IV showed a decline in its level by 38.01% and 43.56%, respectively, with respect to group II (Fig. 6).
IL-10
To confirm if SAV ceases the inflammation in the target organ, anti-inflammatory IL-10 cytokine was measured. Group II showed a decrease in its level by 149.47% with respect to the control, whereas groups III and IV demonstrated an increase in its level by 61.57% and 103.16% as compared to group II (Fig. 7).
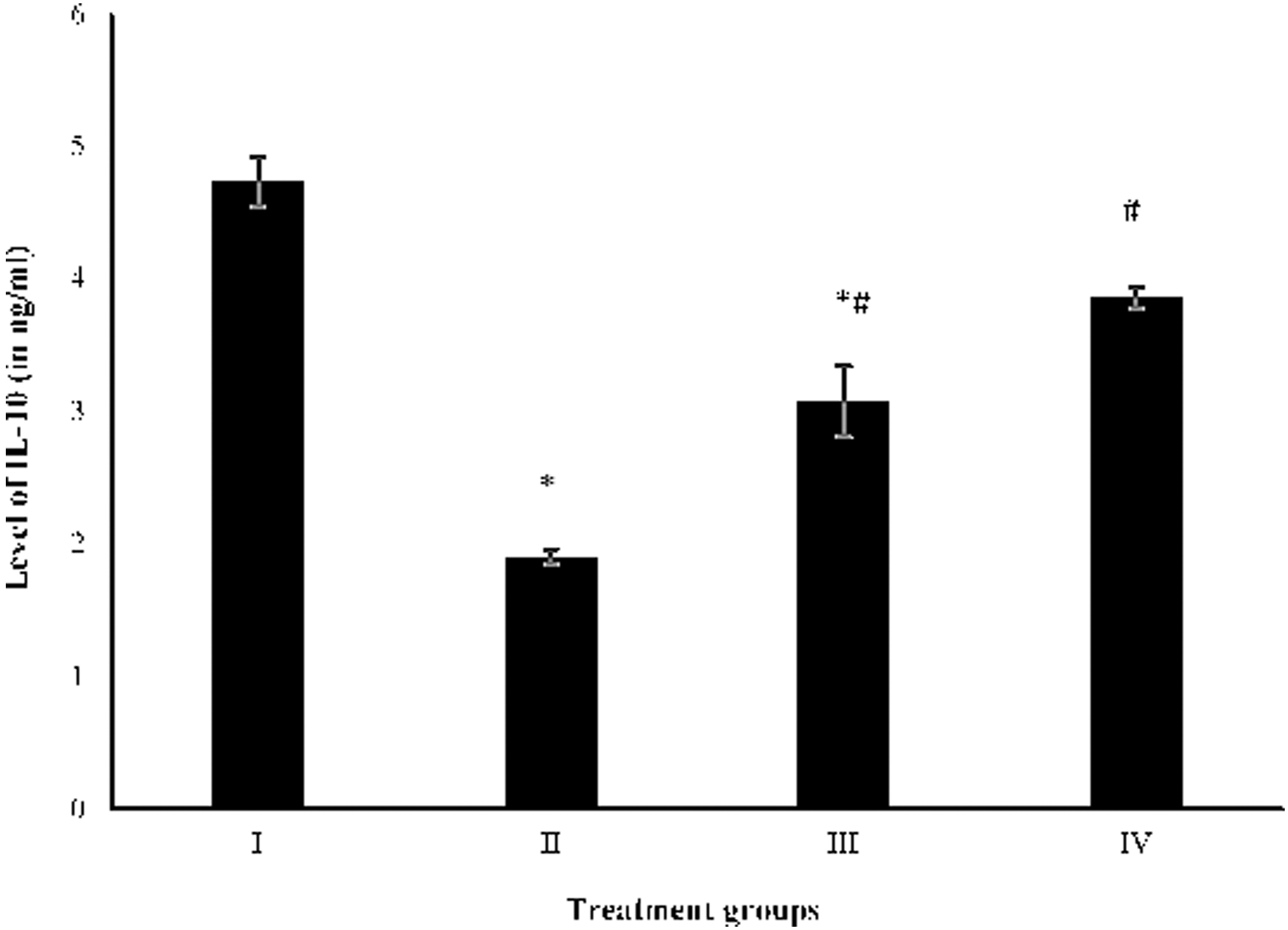
Showing level of IL-10 in the serum samples. Showing level of IL-10 (in nanogram per milliliter of the supernatant in the serum) by ELISA analysis. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control (group I). #Significantly different values from group II. SEM, standard error of the mean.
Measurement of expression of Nuclear Factor-kappa B by RT-PCR
It is an important marker gene to assess the extent of necrosis in the target tissue. Group II exhibited an increase in its expression by 319.49% as compared with the control, whereas group III and IV showed a decline in its expression by 40.61% and 52.75% with respect to group II (Fig. 8).

Showing level of NF-kB and IkB in the liver samples. Showing level of NF-kB and IkB (in nanogram per milliliter of the supernatant of the liver tissue sample) by RT-PCR. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control, group I. #Significantly different values from group II. SEM, standard error of the mean.
Measurement of expression of Inhibitor kappa B by RT-PCR
It is a prominent subunit of Nuclear Factor-kappa B (NF-κB), which is considered as an active form once dissociated from the parent transcription factor, while its translocation from cytosol to the nucleus. The expression of this factor was elevated by 208.99% in group II, whereas it was decreased by 54.30% and 52.60% in groups III and IV in comparison to group II (Fig. 8).
TNF-α and FAS
Both are important markers for apoptosis induction after a cell encounters any toxicant or injury/insult in which FAS assessment confirms the occurrence of apoptosis in any cell.
Group II showed an increase in the expression of TNF-α by 593.44% with respect to the group I, whereas groups III and IV demonstrated its decrease by 54.84% and 72.57% as compared with group II (Fig. 9).

Showing level of first apostosis signal and TNF-α in the liver samples. Showing level of FAS and TNF-α (in nanogram per milliliter of the supernatant of the liver tissue sample) by RT-PCR. All data have been expressed in mean ± SEM of five independent set of experiments. *Significantly different values from the control, group I. #Significantly different values from group II. SEM, standard error of the mean.
Moreover, groups II and III showed an increase in the expression of FAS by 31.63% and 66.01% with respect to their control, whereas group IV demonstrated an increase in its expression by 235.94% as compared with group II (Fig. 9).
Discussion
Natural compounds or a mixture of such bioactive compounds from any natural source with antineoplastic potential have been a hot spot for oncological research since long. The natural compounds are a preferred choice over synthesized chemical-based drugs because of their null or a very small degree of toxicity with better acceptability in biological systems under various in vivo studies. 13,25 –28 Moreover, it is reported that the venom from Brachyponera sennaarensis has a significant antitumor effect on breast cancer cells in a dose-dependent and a time-dependent manner without affecting the viability of nontumor cells. 16,20 Also, Heinen and da Veiga 29 have documented that many of the ants produce very potent venoms that have a possibly potent anticancer effect.
The principal components of the venom from Brachyponera sennaarensis, including trimethyl pyrazine and Phenol-2, 4-bis (1, 1 dimethyl ethyl), have demonstrated many pharmacologically essential features in various studies that are very promising for diverse medical and clinical applications. 8,21 Hence, SAV also falls in the same league of nutraceuticals with prominent pharmacological and medicinal properties. It has a rich repertoire of pharmacologically active compounds that has been found useful in the regulation of pain, inflammation, wound healing, and even cancer. 16,20 Recently, their laboratory has published the antioxidant and anti-inflammatory properties of SAV in the rats challenged with CCl4. 14,20,24
The present work is aimed to investigate if these features of SAV can attribute to its potential anticancer activity or not under in vitro and in vivo studies.
In the present study, the in vitro results indicate that SAV is differentially effective in all three cell lines, MCF-7, HepG2, and LoVo. However, the first two cell lines showed a better response to SAV as their cell proliferation was decreased up to 50% at a fixed dose, which was quite significant as compared with the control cells. The cytokines, IL-1β, 6, and 8, along with TNF-α, were also found elevated in HepG2 and MCF-7 significantly post SAV treatment, although they were not much altered in LoVo cells. These findings entail that the SAV triggers different mechanisms of cell death in all three cell lines mediated by differential immunomodulation.
In MCF-7 and HepG2, SAV might trigger a cellular immune response by elevating the cytokines (IL-1β, 6, and 8) and the transcription factor TNF-α. However, SAV might suppress same cellular events in the LoVo cells. This difference in the pattern of immunomodulation might be attributed to the different origin of the cancer cell lines. This distinction in SAV-mediated immunomodulation may be attributed to the differential antineoplastic activity of the venom in the present study. Additionally, it is well known that colon has its normal flora of specific microbes on its mucosal surface that usually assists in its protection from any antigens or pathogen. It is an important part of its innate immunity that plays a vital role in controlling infection and tumorigenesis. It is also reported that toll-like receptors (TLRs) are major influencing factors in innate immunity, especially in the mucosal and gastrointestinal portion of the body, including the colon. 30 –32 In rodents, 11–13 TLRs have been reported so far. Their impaired signaling of innate immunity or any alteration in TLRs has been documented to trigger an abnormal inflammatory response that might facilitate even tumorigenesis. 30,31
Recently, Semlali et al. 30 have reported that TLR4 (one of the types of TLRs) is overexpressed in the colon cancer tumor epithelial cells as compared with the other types of cancers. In the present study, it is highly speculative that the anticancer potential of SAV might act on colon cancer cell line, LoVo, through TLRs. The complex interaction between SAV and TLRs with innate immunity consequently might compromise the overall anticancer potential of SAV in the colon cancer cells among all the three cell lines. Earlier, scientists have shown 33,34 that insulin-like growth factor (IGF) plays a major role in cellular proliferation, survival, and chemoresistance in various forms of cancer, including MCF-7 and HepG2 cells.
Expression of its receptor, IGF-1R is one of the consistent hallmarks of malignant transformation in preclinical breast cancer models in vitro as well in vivo. 35,36 Furthermore, Badr et al. 16 have reported that SAV significantly abrogated the level of IGF-1-mediated breast cancer cell proliferation and actin polymerization. However, in the case of hepatocellular carcinoma and HepG2 cells, it has been published that IGF-II is more involved than IGF-I in its spread and malignancy. 37,38 Hence, the antineoplastic activity of SAV might involve in the downregulation of IGF-I and IGF-II in all the cell lines. As the antineoplastic activity of SAV was more pronounced in MCF-7 and HepG2 as compared with LoVo, it seems the downregulation of IGF by the venom extract might also be one of the attributing factors for the differential anticancer activity.
In addition, each cell line has unique characteristics, including mechanism action of any drug, metabolism, immune response, sensitivity toward any chemical/drug, activation, and expression of cell death receptors, etc. When a cell line is treated with an antineoplastic drug, all the mentioned factors impart together making an intricate network of upregulation and downregulation of various genes and proteins involved in cell division and cell death. The outcome of all these interactions finally decides the fate of the cell, if it should enter into cell cycle or undergo programmed cell death. 39 From results, it seems that SAV might induce an immune response to a different extent in the three cell lines leading to the differential degree of cell death in the cells in the present investigation.
Under the in vivo study, the ELISA of the serum sample from CCl4-treated animals demonstrated a high level of inflammatory cytokines, IL-1β and 6 that were found significantly lowered in the animals with hepatoma after treatment with SAV. Furthermore, the level of anti-inflammatory cytokine, IL-10 was also elevated to a significant extent in the same group. All these results confirm the anti-inflammatory properties of SAV that is a bit contradictory to the in vitro findings. Moreover, the RT-PCR of TNF-α, NF-kB, and its active form, Inhibitor kappa B (Iκ-B) in the liver samples of the fourth group, demonstrated a vivid decline in their expression as compared with the group II. Furthermore, Fujii et al. 40 have shown that IL-10 can help CD8(+) T cells in the sustenance of anticancer activity.
Earlier, IL-10 has been reported to decrease the tumor burden in the rodents. 41 The elevated level of NF-kB and its subunit Iκ-B are essential requisites in cellular proliferation in malignant condition as they are supposed to abrogate the caspase-mediated apoptosis. 42 However, decreased level of these transcription factors indicates ceasing of inflammation and tissue damage. Furthermore, to confirm the mode of cell death in the damaged tissues, the expression of FAS and TNF-α were assessed. The pattern of both the parameters clearly entails that SAV triggers apoptosis in the target organ significantly by controlling the inflammation and tissue injury. These findings are, by their previous studies, showing SAV at the given dose as a strong antioxidant agent that has helped the CCl4-challenged animals in the restoration of tissue damage and retaining of normal function. 20,24 It is well known that FAS, type I membrane receptor, belongs to a member of the TNF-receptor superfamily. The activation of FAS induces apoptosis by its ligand through activation of the caspase cascade, which is a hallmark of apoptosis. 43 It is noteworthy that a moderately higher level of NF-kB and IL-10 is assumed to increase the sensitivity of many chemotherapeutic agents and even directs cellular machinery to undergo apoptosis. 42,44
Also, Badr et al. 16 have documented that SAV abrogates IGF-1-triggered phosphorylation of ERK and AKT signaling modulation that consequently flags strong apoptotic signals through p38MAPK phosphorylation. Hence, the present study along with the previously published works confirm that SAV has biologically active molecules of immunomodulatory effects that orchestrate the immune system favoring apoptosis in the target tissues over the necrotic mode of cell death. Alternatively, SAV has demonstrated that its therapeutic dose organizes the antioxidant machinery to nullify the free radicals that further decrease the aggression of the inflammatory response promoting tissue healing in the damaged tissues and apoptosis in the unrepairable sections. 20,45 –47
Additionally, it is also established that cellular redox status determines the fate of a cell if it will undergo cell cycle or die through apoptosis or necrosis. 18,48 It is evitable that SAV in the treated cell lines and the rodents might dictate the redox status that favors cell death by apoptosis at the given dose as found in several studies (Fig. 10).

Concept arrow-diagram on efficacy of anticancer activity of SAV. Showing concept arrow-diagram on efficacy of SAV after cell line and animal-based studies highlighting the major events. >Indicates more incidences of the mode of cellular death. SAV, samsum ant venom.
Moreover, the differential antiproliferative effect on different cell lines also exhibits its preferential specificity for specific target tissues, such as breast and liver cancer cells over the colon cancer cell line. Their current findings also correlate well with their earlier work 20,24 in which SAV has shown potent antioxidant nature of its biocomponents in the rodent model study. The malignancy and propagation of any cancer in vivo is attributed to free radical-mediated oxidative stress and related extent of inflammation. 48,49 In the present study, SAV seems to moderate the extent of inflammation up to the level in which only the healthy cells can thrive, whereas the cancerous cells are rerouted toward apoptosis at the same time.
Hence, the antioxidant potential of SAV might dictate the CCl4-induced tissue injury and mild tumorigenesis in the treated animals in such a way that apoptosis became the preferred mode of cell death over the necrosis. 20,47 However, the study needs to be done with other cell lines and cancer models of different organs in vivo to confirm the efficacy of SAV as an anticancer agent.
It is noteworthy that SAV exerts its anticancer activity by enhancing inflammatory response in vitro leading increased cell death, whereas the venom extract demonstrates its antineoplastic activity by ceasing inflammation in vivo orchestrating the condition that favors apoptosis. Nevertheless, SAV is a complex mixture of diverse types of biologically active compounds that have not been fully elucidated so far. Hence, further study is warranted to identify these essential bioactive components for their exact mechanism and target-specific application under clinical trials for effective cancer treatment.
Conclusion
The present study establishes anticancer activity of SAV in vitro as well as in vivo. The venom extract at the standard dose is antioxidant in nature that triggers immunomodulation (IL-1β, IL-6, IL-10) and regulates the key transcription factors (NF-κB, Iκ-B, TNF-α) leading to the ceasing of the inflammation concomitant with an increased rate of tissue healing. All these events promote apoptosis in the tumorous and damaged cells that consolidates SAV as an important anticancer agent with promising clinical implication. However, the venom is a mixture of complex chemical ingredients of the diverse type that can trigger various apoptosis and cell death pathways that needs further in-depth research to pave the exact mode of pathways in vitro as well in vivo.
Authors' Contribution
J.A. had conducted animal handling and treatment and preparation of samples for in vivo experiments. A.S. organized the RT-PCR experiments that were executed with J.A., I.H., and H.E. had analyzed the results and discussion. I.H. and J.A. had prepared the graphical representation of the results. The article was written by I.H., J.A., and S.H.M. that was crosschecked by H.E., I.A., and M.A., and M.A. supervised and provided the laboratory facilities for conducting experiments, and animal and cell culture treatment.
Footnotes
Acknowledgment
The authors would like to extend their sincere appreciation to Deanship of Scientific Research at King Saud University for funding the research group no. RGP-1438-004.
Disclosure Statement
No competing financial interests exist.