
Abstract
Background:
Nonspecific siRNA expression limits its application in cancer gene therapy. Therefore, a tightly regulated and reversibly inducible RNAi system is required to conditionally control the gene expression. This investigation aims at constructing a hypoxia/colorectal tumor dual-specific bidirectional short hairpin RNA (shRNA) expression vector.
Materials and Methods:
First, carcinoma embryonic antigen (CEA) promoter designed in two directions. Then, pRNA-bipHRE-CEA vector was constructed by insertion of the vascular endothelial growth factor enhancer between two promoters for hypoxic cancer-specific gene expression. To confirm the therapeutic effect of the dual-specific vector, two shRNA oligonucleotides were inserted in the downstream of each promoter. QRT-polymerase chain reaction and western blot assays were performed to estimate the mRNA and protein expression levels.
Results:
Both mRNA and protein levels were significantly reduced (50%–60%) in the hypoxic colorectal cancer-treated cells when compared with the controls.
Conclusion:
The novel bidirectional hypoxia-inducible shRNA expression vector may be efficient in colorectal cancer-specific gene therapy.
Introduction
Short hairpin RNA (shRNA) is an efficient and widely used technique for silencing gene expression in mammalian cells. Although this approach has several advantages, such as specific and long-term gene silencing, there are certain limitations that prevent this vector-based approach from being widely used in gene therapy. The core obstacle is the nonspatial and nontemporal control of siRNA expression, causing adverse effects on nontarget tissues. Other concerns include triggering of type I immune responses and off-target effects. 1 –4
For a successful and safe gene therapy, precise regulation of gene expression is required to minimize the side-effects in normal tissues. To this end, certain inducible RNAi systems have been developed and used in gene therapy for in vitro and in vivo studies of different cancers. These include Cre/loxP-, ecdysone-, Lac repressor-, and tetracycline-inducible systems. 5 –10 However, siRNA expression in the absence of an inducer (leakiness of these systems) is still a major concern that limits application in disease treatment. 11 –13
Hypoxia as a solid tumor feature that could be targeted by application of the hypoxia tumor- specific promoters, which is known as an effective strategy with high specificity in cancer gene therapy. 14 –17 Therefore, combining RNAi technology with such a dual-targeting transcriptional regulation system may provide more sophisticated gene expression control.
Carcinoma embryonic antigen (CEA) is an oncofetal tumor marker overexpressed in epithelial cancer cells, such as colorectal cancer cells (over 90%), but not in normal cells. 18 –20 The promoter of CEA has been extensively used in colorectal cancer-specific gene therapy. 21 –24 Moreover, when the function of this promoter was compared with that of a viral promoter, higher and more persistent activity was observed in CEA-positive human cancer cells. 25 Considering all the evidence, the authors hypothesize that combination of this tumor-specific promoter with a hypoxia inducible factor 1/hypoxia response element (HRE) system may provide maximum gene expression with a high level of specificity for colorectal cancer. Hence, this inducible promoter may prove to be an effective candidate for the construction of a vector-based inducible system to express shRNA in colorectal cancer cells.
In addition, most commercial vectors are designed exclusively for the expression of a single gene, whereas bidirectional expression vectors can express two genes simultaneously, which can be used to induce gene expression or in gene silencing techniques. In addition, the simultaneous silencing of multiple genes in cancer therapy has the advantage of minimizing multiple drug resistance caused by small chemical molecules. 26,27
In this study, a novel bidirectional hypoxia-inducible shRNA expression vector was constructed for colorectal cancer-specific gene therapy. To the best of knowledge, this is the first study to explore such a dual-targeting transcriptional regulation system for use in cancer gene therapy by knockdown with siRNAs. In addition, the therapeutic effect of the dual-specificity shRNA vector was verified by placing β-catenin- and Bcl-2-specific shRNAs downstream of the promoters.
Materials and Methods
Construction of a bidirectional hypoxia-inducible shRNA expression vector
Using bioinformatic tools, Web Promoter Scan Service and Neural Network Promoter Prediction, the core promoter sequence of CEA (428 bp) and its enhancer (1.4 kbp) were selected and designed in opposite directions. The HREs of the vascular endothelial growth factor (VEGF) gene (eight tandem copies) 5′-CCACAGTGCATACGTGGGCTCCAACAGGTCCTCTT-3′ (35 bp) were placed upstream of each promoter to generate a bidirectional hypoxia-responsive promoter. Finally, this bidirectional expression cassette (2749 bp) was synthesized by the Shine Gene Molecular Biotech Company and subsequently cloned into the pRNAT-U6.1/Neo expression vector (6380 bp) (Fig. 1). Double digestion analysis was used to verify substitution of the U6 promoter into the bidirectional CEA promoter.
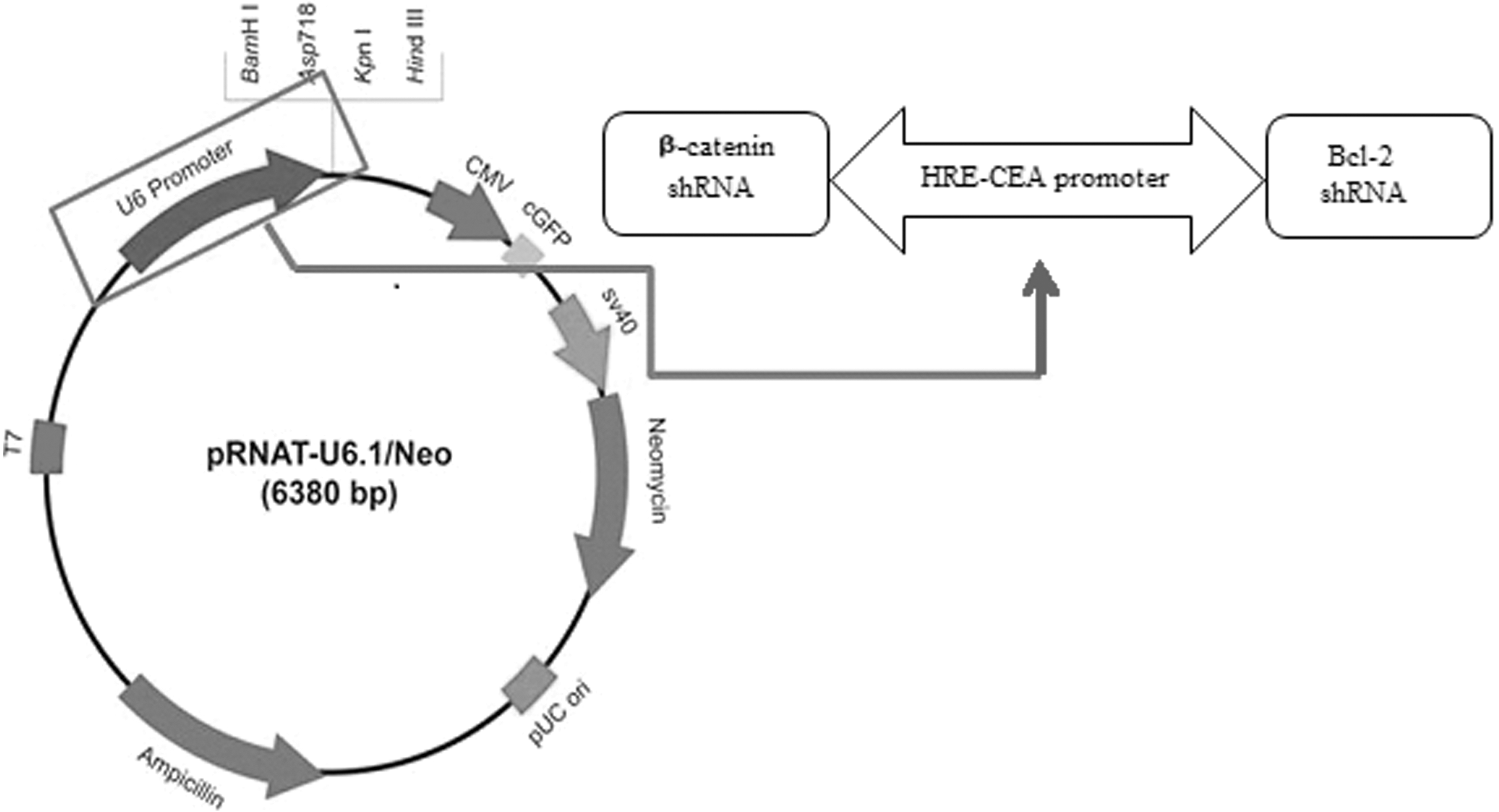
pRNAT-bipHRE-CEAsh (Bcl-2-β-catenin) vector map. The bipHRE-CEA promoter was replaced by the U6 promoter in the pRNAT-U6.1/Neo expression vector. CEA, carcinoma embryonic antigen; HRE, hypoxia response element.
To assess the activity of the promoter construct, specific shRNAs for the β-catenin and Bcl-2 genes were designed via InvivoGen-siRNA Wizard software, synthesized, and inserted downstream of each promoter (Fig. 2). Subsequently, a bidirectional hypoxia-inducible shRNA expression vector was constructed.
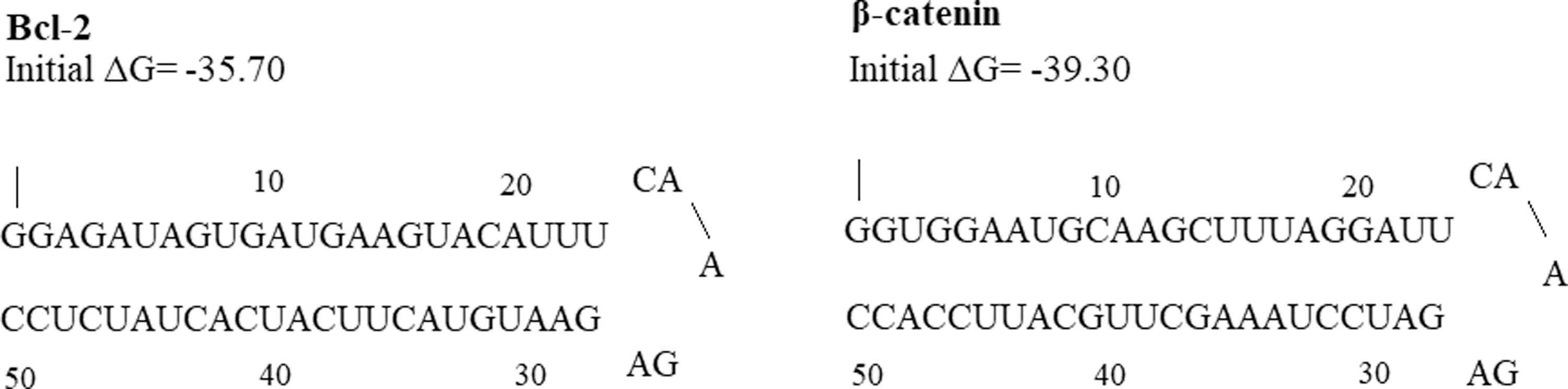
The second structure of Bcl-2 and β-catenin shRNAs. shRNAs, short hairpin RNAs.
Cell culture
The human colon cancer cell line HT-29, human breast carcinoma cell line MCF-7, and human cervical carcinoma cell line Hela were obtained from the Pasteur Institute (Tehran, IRAN). Each cell line was grown in complete growth medium RPMI 1640 + 10% (v/v) fetal bovine serum +1% (v/v) penicillin/streptomycin antibiotic solution (Gibco-BRL, Rockville, MD) at 37°C in a humidified incubator with 5% CO2. The cells were subcultured three times before experiment.
Transfection and GFP assay
Twenty-four h before transfection, cells were seeded into two 6-well culture plates at 6 × 105 cells per well. The cells were transfected with the recombinant shRNA expression vector using Lipofectamine 3000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. After a 24-h incubation, the cells were incubated for an additional 48 h under hypoxic (1% oxygen) or normoxic (20% oxygen) conditions. Transfection efficiency was evaluated by observing GFP signals under fluorescent microscopy. GFP was separately controlled by the CMV promoter.
RNA extraction and qRT-polymerase chain reaction analysis
The treated and untreated cells were washed twice with PBS and harvested following a 72-h incubation. Total RNA was extracted from cells using the TRIzol reagent according to the manufacturer's instructions. RNAs were reverse-transcribed using the PrimeScript RT reagent kit (Thermo). Quantitative polymerase chain reaction (PCR) experiments were used to analyze Bcl-2 and CTNNB1 gene expression using a standard SYBR Green PCR kit (Thermo) protocol. PCR was carried out in triplicate to generate three independent experiments. PCR products were quantified and normalized using β-actin as an internal standard. The sequences of the primers (Metabion) used were as follows: Bcl-2 Forward: 5′-GGTGGGGTCA TGTGTGTGG-3′, Reverse: 5′ CGGTTCAGGTACTCAGTCATCC-3′; β-catenin Forward: 5′-TGGCTTGGAATGAGACTGCTG-3′, Reverse: 5′-CCACCCATCTCATGTTCCATC-3′; and β-actin Forward: 5′-CATGTACGTTGCTATCCAGGC-3′, Reverse: 5′-CTCCTTAATGTC ACGCACGAT-3′.
Western blot analysis
At 72 h posttransfection, HT-29 cells were lysed in 150 μL of lysis buffer (150 mM/L NaCl, 100 mM/L Tris-Hcl, 5% sodium deoxycholate, 5% sodium dodecyl sulfate, 1% Triton X-100, 5 mM ethylenediaminetetraacetate, 10% glycerol, 0.2 mM/L PMSF, and antiprotease) on ice. The cells were then centrifuged at 12,000 g for 20 min. The supernatant was collected for the analysis of protein concentration by Bradford protein assay. The extracted proteins were separated by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto nitrocellulose membranes (Whatman GmbH, Germany). Proteins were incubated with β-catenin and Bcl-2 antibodies (BioLegend) at 1:500 and 1:200 dilutions, respectively, and then with horseradish peroxidase-conjugated secondary antibody (Santa Cruz). SIGMAFAST™ 3,3′-diaminobenzidine tablets (Sigma) were used to visualize protein bands. The β-actin protein was chosen as an internal control.
Statistical analysis
Each experiment was performed in at least three biological replicates, and all data are reported as mean ± standard deviation. Comparisons between two groups were made using Student's t-test. A significant difference was defined as p < 0.05.
Results
Construction of hypoxia-inducible dual-specific shRNA expression vector
The final sequence (2749 bp) was cloned into the EcoRI and SacI RE sites and was verified by double digestion and DNA sequencing.
Downregulation of Bcl-2 and β-catenin genes at the mRNA level by the pRNAT-bipHRE-CEAsh (Bcl-2-β-catenin) vector
To evaluate the tumor specificity and efficacy of gene silencing with this dual targeting system, the pRNAT-bipHRE-CEAsh (Bcl-2-β-catenin) vector, constructed by insertion of specific shRNA against Bcl-2 and β-catenin genes under the control of the bipHRE-CEA promoter, was transfected into HT-29, MCF-7, and Hela cells. In the HT-29 cell line, the expression inhibition rates under hypoxia were 56% (55.75% ± 2.21%; n = 3) and 51% (50.62% ± 3.18%; n = 3) for β-catenin and Bcl-2, respectively (p < 0.05). The HRE-CEA promoter showed a much higher activity (2–3 times) under hypoxia compared with normoxia (Fig. 3). Because the gene silencing efficacy of the promoter in the two directions was nearly equal, to examine the tumor specificity of the modified CEA promoter, Bcl-2 shRNA expression driven by the bipHRE-CEA promoter was investigated in HeLa as a CEA-nonproducing cell control and in MCF-7 cells as a moderately CEA-expressing cell line under hypoxia and normoxia. As shown in Figure 4, the expression level of Bcl-2 mRNA under hypoxia and normoxia was not significantly different between the two cell lines, and it only showed ∼13% (12.66% ± 1.45%; n = 3) and 5% (5.2% ± 0.94%; n = 3) reduction under hypoxia in MCF-7 and Hela cells, respectively, compared with controls. These data verified the high tumor specificity of the bipHRE-CEA promoter.
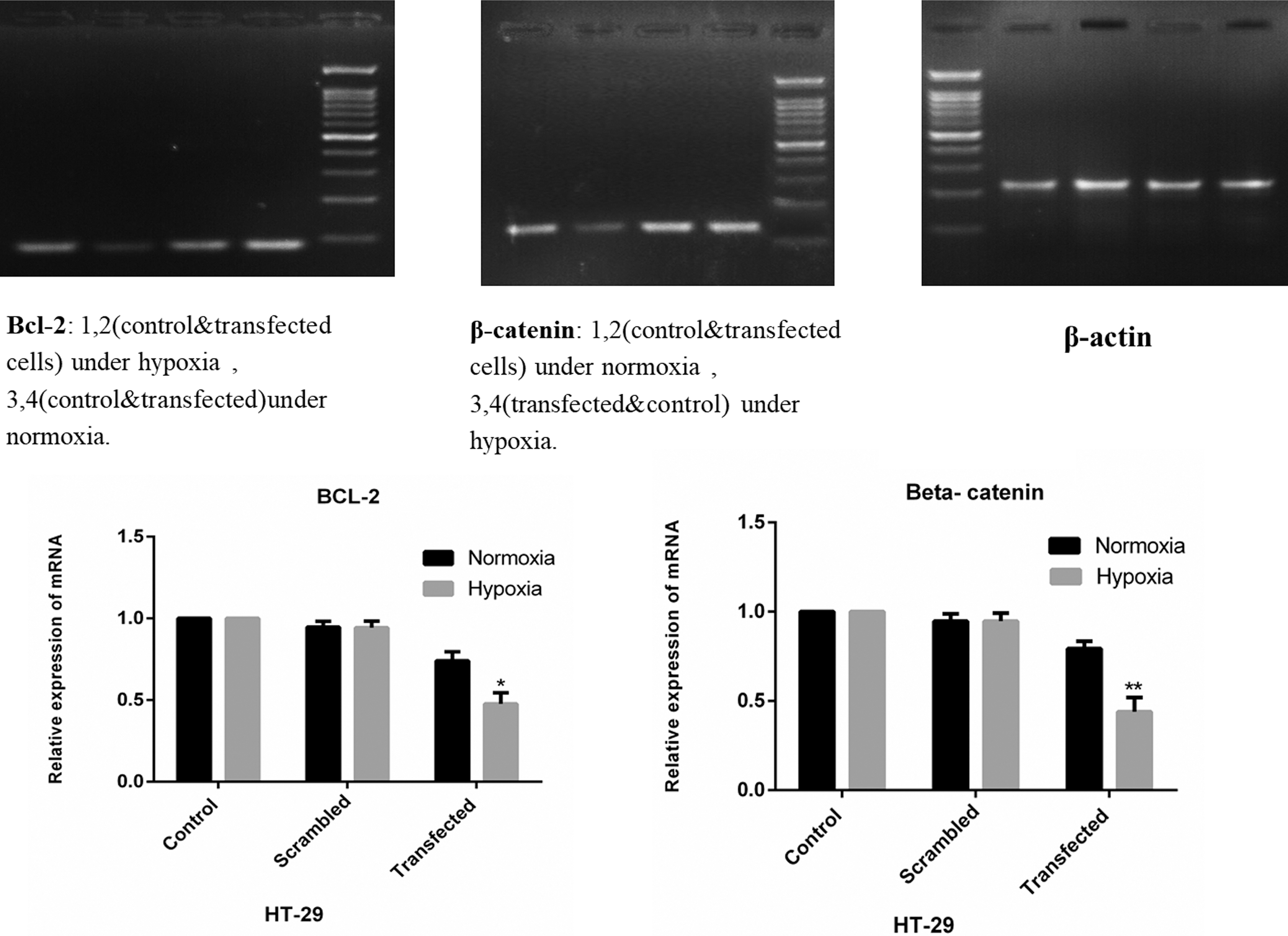
The mRNA expression levels of β-catenin and Bcl-2 in HT-29 cells after shRNA expression vector transfection. Cells were transfected with hypoxia-inducible shRNA expression vector targeting β-catenin and Bcl-2 under hypoxia and normoxia. At 72 h posttransfection, both gene expression levels were measured by real-time PCR assay (*p = 0.0013; **p = 0.008). PCR, polymerase chain reaction.
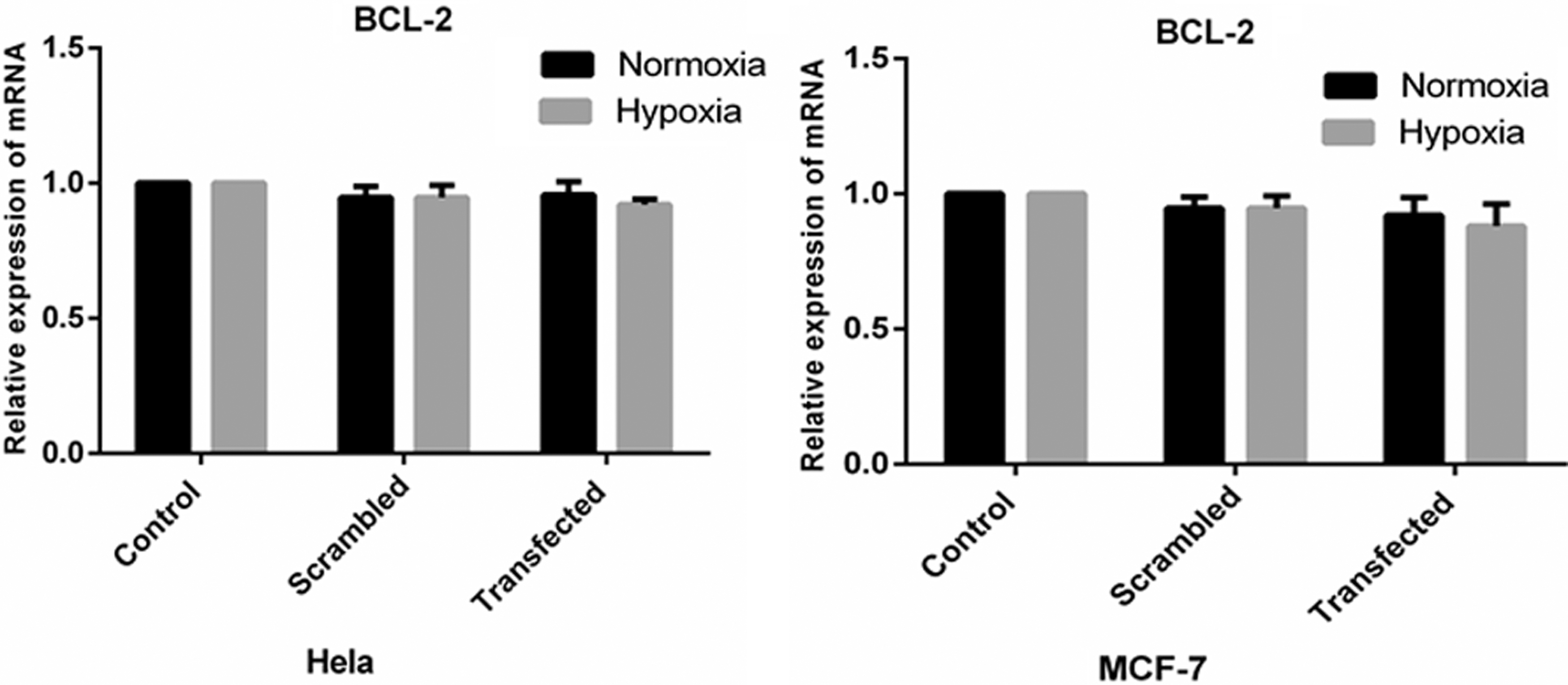
The mRNA expression level of Bcl-2 in Hela and MCF-7 cells after shRNA expression vector transfection. Cells were transfected with hypoxia-inducible shRNA expression vector under hypoxia and normoxia. At 72 h posttransfection, Bcl-2 gene expression level was measured by real-time PCR assay.
Downregulation of Bcl-2 and β-catenin genes at the protein level by the pRNAT-bipHRE-CEAsh (Bcl-2-β-catenin) vector
Western blot analysis was used to verify the real-time PCR results and showed a significant reduction in the protein levels of both genes (p < 0.05) (Fig. 5). The inhibition rates were 52% (51.82% ± 7.25%; n = 3) and 46 (46.12% ± 7.35%; n = 3) for β-catenin and Bcl-2, respectively (p < 0.05). Collectively, these results indicated that the bidirectional hypoxia/colorectal tumor dual-specificity shRNA expression vector simultaneously knocked down the β-catenin and Bcl-2 mRNA and protein expression levels, and the VEGF enhancer affected the promoter's performance and led to a significant increase in its activity under hypoxia compared with normoxia.
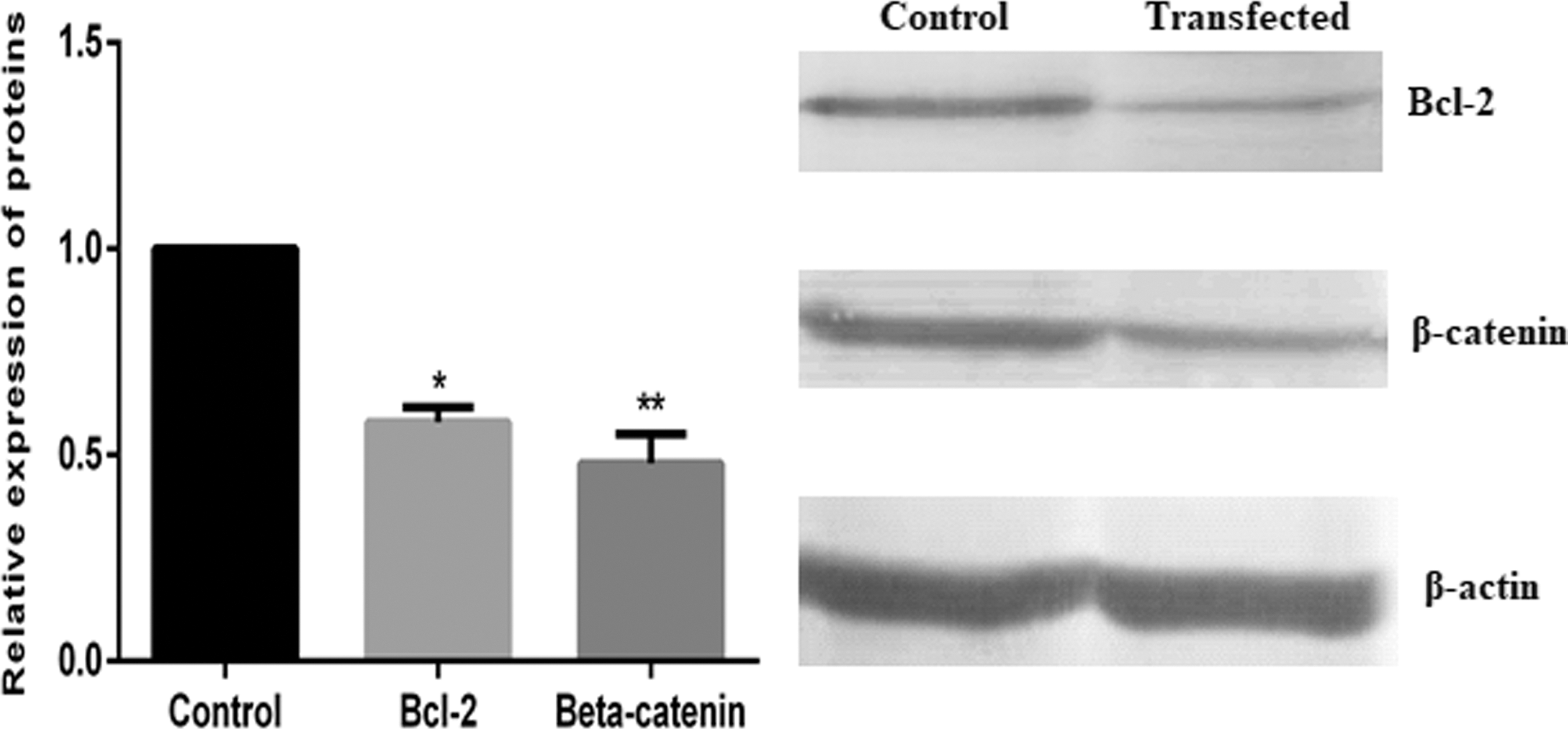
Protein expression of β-catenin and Bcl-2 in HT-29 cells following shRNA expression vector transfection. Relative expression of β-catenin and Bcl-2 protein levels was measured by western blot analysis at 72 h posttransfection under hypoxia (*p = 0.008; **p = 0.009).
Discussion
A dual hypoxia/colorectal tumor-specific vector involving a bidirectional CEA mini promoter and HREs of the VEGF gene was constructed for stronger regulation of gene expression under hypoxic conditions.
Solid tumors contain hypoxic regions consisting of highly treatment-resistant cells. Many studies have demonstrated that silencing genes such as those involved in chemoresistance, those belonging to the antiapoptotic Bcl-2 gene family, and those encoding double-strand break repair enhance the effects of chemotherapy and radiotherapy. 28 –30 In addition, hypoxia provides an opportunity to specifically target treatment-resistant malignant tumor cells and not normal tissues. 31 –33 Therefore, hypoxia-inducible gene expression systems have been developed to target gene expression to hypoxic cells. However, hypoxia-inducible systems have been investigated more in suicide gene therapy than in RNAi-based cancer gene therapy. 34 Therefore, the authors have developed a new bidirectional hypoxia-inducible shRNA system, which is composed of the tissue-specific CEA promoter and HRE.
Because previous investigations have shown that HREs of the VEGF gene have the highest inducibility in hypoxia and lower baseline activity in normoxia, these elements were selected and applied as eight tandem repeats upstream of each promoter. They also confirmed that gene activation was influenced by the number of these elements present, although there is still no consensus on HRE copy number. 16,35,36 However, a saturation effect was reported for constructs containing >6 copies of HRE. 34,37 The bidirectionally proportional activity of the HRE was confirmed in previous investigations. 36,37
In the present research, the bidirectional activity of multiple HREs in combination with the CEA promoter was investigated in an RNAi-mediated gene silencing strategy. For this purpose, the authors inserted sequences of specific shRNAs against β-catenin and Bcl-2 downstream of the promoters to construct pRNAT-bipHRE-CEA shRNA (Bcl-2-β-catenin) vector. The silencing effect of the hypoxia response shRNA expression vector was relatively low (18%–25%) under normoxia, whereas it significantly increased to ∼50%–60% after hypoxia induction in the HT-29 cell line.
The HRE-CMV promoter has been used to regulate siRNA expression in response to hypoxia. 38 Although the CMV promoter directs higher levels of shRNA expression than tissue/tumor-specific promoters, the lack of cell specificity and transient expression activity are serious disadvantages that prevent its possible application in the therapeutic field. 39 Investigations have shown that the CEA promoter has the advantage of providing long-term expression in vivo. 25 According to previous research, the CEA promoter has very low expression in CEA-negative cells, such as HeLa cells, revealing the sophisticated tumor specificity of this promoter, which could significantly reduce the basal expression level and side-effects. 22,25 As these data show (Fig. 2B), the bipHRE-CEA promoter also shows high tumor specificity. However, leaky expression still exists under normoxia that could minimize via transductional targeting.
In summary, these results suggest that the combination of the VEGF gene's HRE and the CEA promoter specifically increases gene expression under hypoxia in colorectal cancer cell lines. Therefore, the novel hypoxia-inducible siRNA expression system is a potential tool for RNAi-based cancer gene therapy.
Footnotes
Authors' Contributions
Majid Shahbazi designed the study and analyzed the results. Bita Javan conducted the experiments and wrote the article.
Acknowledgment
The authors thank the Golestan University of Medical Sciences for funding this work.
Disclosure Statement
No competing financial interests exist.