
Abstract
Sodium phenylbutyrate (SPB) as a salt of 4-phenylbutyric acid (4-PBA) has been reported to be an ammonia scavenger, histone deacetylase inhibitor, and an endoplasmic reticulum stress inhibitor in various diseases, including neurological diseases, inflammatory disorders, and carcinogenesis. Although phenylbutyrate showed effective antitumor properties in many cancers, its role in oral squamous cell carcinoma (OSCC) remains further characterized. Thus, the OSCC cell lines CAL27, HSC3, and SCC4 were treated with a series of doses of SPB for different times. The IC50 of three cell lines for SPB was determined to be 4.0, 3.7, and 3.0 mM. The CCK-8 assay indicated that the treatment of SPB induced continuous inhibition of cell vitality of three cell lines. Apoptosis was assessed by Hoechst assay that showed that SPB could significantly promote cell apoptosis. Moreover, the apoptosis-related pathway was analyzed, and the results showed that the expression of antiapoptosis factor BCL-2 was downregulated by SPB but the cleavage of caspase-3 was increased. Meanwhile, it was found that SPB also impaired the migration and invasion of OSCC cells in vitro. Mechanistically, the transforming growth factor-β (TGFB) related epithelial–mesenchymal transition (EMT) was inhibited by SPB with decreased mesenchymal marker N-cadherin and increased epithelial marker E-cadherin. Furthermore, the antitumor effect of SPB in vivo was also demonstrated. The administration of SPB induced remarkably tumor regression with decreased tumor volume, and the TGFB level and EMT phenotype in vivo were also inhibited. These data demonstrated that the treatment of SPB could function as antitumor therapeutics for OSCC.
Introduction
Oral squamous cell carcinoma (OSCC) is the main type of oral cancer that is a major public health problem worldwide. As the sixth most common malignancy, the high and increasing incidence rates of OSCC are shown in developing countries. 1 The malignant phenotype of OSCC is derived from the transformation of normal oral mucosa to premalignant changes, such as leukoplakia and submucous fibrosis, which are reversible, and 3%–12% of the premalignant lesions eventually could progress to OSCC. 2 Although with advances in recognition and treatment in recent years, no significant improvement of the 5-year overall survival rate has been obtained, which was attributed to the advanced stages of disease at late-stage diagnosis and the frequent development of relapse and second primary tumors. Chemoradiotherapy is often combined to surgery for the treatment of OSCC patients, but it is not effective on late-stage metastatic tumors. Thus, identification of new treatment for OSCC is indispensible for improving the efficiency of cancer therapy. 3 –5
Sodium phenylbutyrate (SPB) is a salt of an aromatic fatty acid, 4-phenylbutyrate (4-PBA) or 4-phenylbutyric acid, which was first recognized as a therapeutic for inhibiting platelet aggregation 4-PBA in the mid-70s. 6 Currently, phenylbutyrate is an FDA-approved drug for ammonia removal and used for the treatment of urea cycle disorders. 7 Mounting pieces of evidence have identified other functions of phenylbutyrate, including histone deacetylase inhibitor (HDACi), an endoplasmic reticulum (ER) stress inhibitor, and regulator of mitochondrial biogenesis. 8 Therefore, the potential applications of phenylbutyrate were investigated in many diseases. Ozcan et al. reported that PBA functioned as leptin-sensitizing agents to decrease ER stress and inactivate the unfolded protein response in the hypothalamus of obese mice for reactivation of leptin receptor signaling. 9 Phenylbutyrate could also reverse hepatic fibrosis through inhibiting hepatic inflammatory response, repressed the expression of transforming growth factor beta (TGF-β), and α-smooth muscle actin (α-SMA), which were the critical factors for the metastasis of cancer cell. 10,11 Actually, the antitumor effects of phenylbutyrate were discovered in many cancers. Xu et al. found that the treatment of SPB to prostate cancer induced impaired cell vitality, clone formation, and the ability of migration. 12 However, the role of SPB in OSCC and the potential mechanism were unclear.
Epithelial–mesenchymal transition (EMT) is the process whereby cells shed their epithelial properties through downregulation of epithelial marker E-cadherin, etc. and adopt a more mesenchymal and invasive phenotype through upregulation of mesenchymal markers N-cadherin, α-SMA, etc. 13 EMT was recognized as a critical manipulator of metastasis during the carcinogenesis. TGF-β family members are essential cytokines derived from the tumor microenvironment that can initiate and maintain EMT, and TGF-β-activated EMT pathway was found to be involved in the development and metastasis of OSCC. 14,15 Thus, considering the inhibition of phenylbutyrate on TGF-β, it was speculated that SPB is able to regulate the TGF-β level of OSCC cells to inhibit the EMT phenotype.
In this study, the OSCC cell lines were treated with SPB, and the IC50 of SPB and the effects on cell vitality, apoptosis, and EMT phenotype in vitro and in vivo were estimated.
Materials and Methods
Cell lines and reagents
The OSCC cell lines CAL27, HSC3, and SCC4 were purchased from the cell bank of the Chinese Academy of Sciences (Shanghai, China) and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (Life Technologies), ampicillin, and streptomycin at 37°C, 5% CO2 conditions. Anti-GAPDH, caspase-3, and Bcl-2 antibodies were obtained from Cell Signaling Tech (Denver, MA).
CCK-8 assay
The OSCC cell lines were treated with SPB and then were harvested to wash with phosphate-buffered saline (PBS), and then cell counting kit-8 (Kumamoto, Japan) mixed with DMEM was used for cell viability assay, and the absorbance was measured at 450 nm by a microplate reader.
Hoechst staining assay
The conditional OSCC cell lines were harvested and incubated with Hoechst 33342 (5 μg/mL; Sigma) for 10 min at room temperature. After washing with 0.5% Triton X-100 in PBS, the changes in nuclear morphology were observed under a fluorescence microscope (OLYMPUS, Tokyo, Japan). Each experiment was repeated three times.
Flow cytometer assay
To determine the apoptosis, the cells were fixed in cold 70% ethanol at −20°C for 2 h. Then 10 mg/mL RNase was added, the cells were stained with 2 μL of annexin V mixed with 2 μL of propidium iodide (PI, eBioscience). The assay was used according to the manufacturer's instructions and analyzed using a flow cytometry.
Western blot assay
Total proteins of OSCC cells were extracted by lysing cells in RIPA buffer with Protease Inhibitor Cocktail (Pierce). BCA Protein Assay Kit (Beyotime) was used to measure the protein concentration. Equivalent amounts of protein were separated by 10% SDS-PAGE polyacrylamide gels and then transferred to PVDF membranes (Millipore). The membranes were blocked with 5% nonfat milk in Tris-buffered saline and then incubated with primary antibodies followed by horseradish peroxidase-conjugated secondary antibody (Abcam). The protein signals were detected by ECL chemiluminescence kit and quantified using ImageJ software. GAPDH was used as a loading control.
Transwell assay
A total of 2 × 104 OSCC cells transfected with or without SPB were in the upper chamber of a noncoated transwell insert for the migration assay. Cells that did not migrate were removed using a cotton swab and were stained by crystal violet and counted under an inverted microscope. Five random views were selected to count the cells.
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from tissues and cells using TRIZOL reagent (Invitrogen, Life Technologies) according to the manufacturer's protocols. For quantitative real-time polymerase chain reaction (qRT-PCR), 1 μg of total RNA was reverse transcribed to cDNA using Reverse Transcription Kit (Takara, Dalian, China). The PCR was set for 50°C for 2 min, followed by 40 cycles of 95°C for 15 s, then at 60°C for 1 min. The expression level (2−ΔΔCt) of TGF-β was determined. GAPDH was used to normalize the relative abundance of TGF-β.
Immunofluorescence
The EMT phenotype of OSCC cells was determined by immunofluorescence assays, which are performed according to a previous study. 16
Tumor model
A total of 2 × 106 CAL27 cells were subcutaneously injected in rear flank of nude mice (4 mice per group) and 2 mmol/kg was administrated to the mice bared with tumor. The tumor sizes were measured 1 week apart and the tumor volumes were calculated as V (cm3) = width 2 (cm2) × length (cm)/2.
Statistical analyses
The results are analyzed by the Statistical Package for Social Sciences version 16.0 (SPSS 16.0; SPSS, Inc., Chicago, IL) and the Prism statistical software package (Version 5.0; Graphpad Software, Inc.). Unpaired t-tests or Mann–Whitney U tests were used to compare the two groups. p < 0.05 was considered statistically significant. All experiments were performed at least three times.
Results
The IC50 of OSCC cell lines for SPB was determined
To investigate the antitumor effects of SPB on OSCC, the OSCC cell lines CAL27, HSC3, and SCC4 were used in this study. A series of doses of SPB including 1, 2, 4, 8, and 16 mM were treated with CAL27, HSC3, and SCC4. The cell vitality of CAL27, HSC3, and SCC4 was impaired in a dose-dependent manner and the IC50 of SPB for three cells was 4.091, 3.712, and 3.015 mM, respectively (Fig. 1A). Thus the concentration of 4 or 3 mM of SPB was used in subsequent assays. HSC3 and SCC4 cells were treated with SPB for 12, 48, and 72 h, and the results of CCK-8 indicated that SPB significantly inhibited the cell vitality, even the cells were treated for 12 h, and this inhibition was time dependent (Fig. 1B).
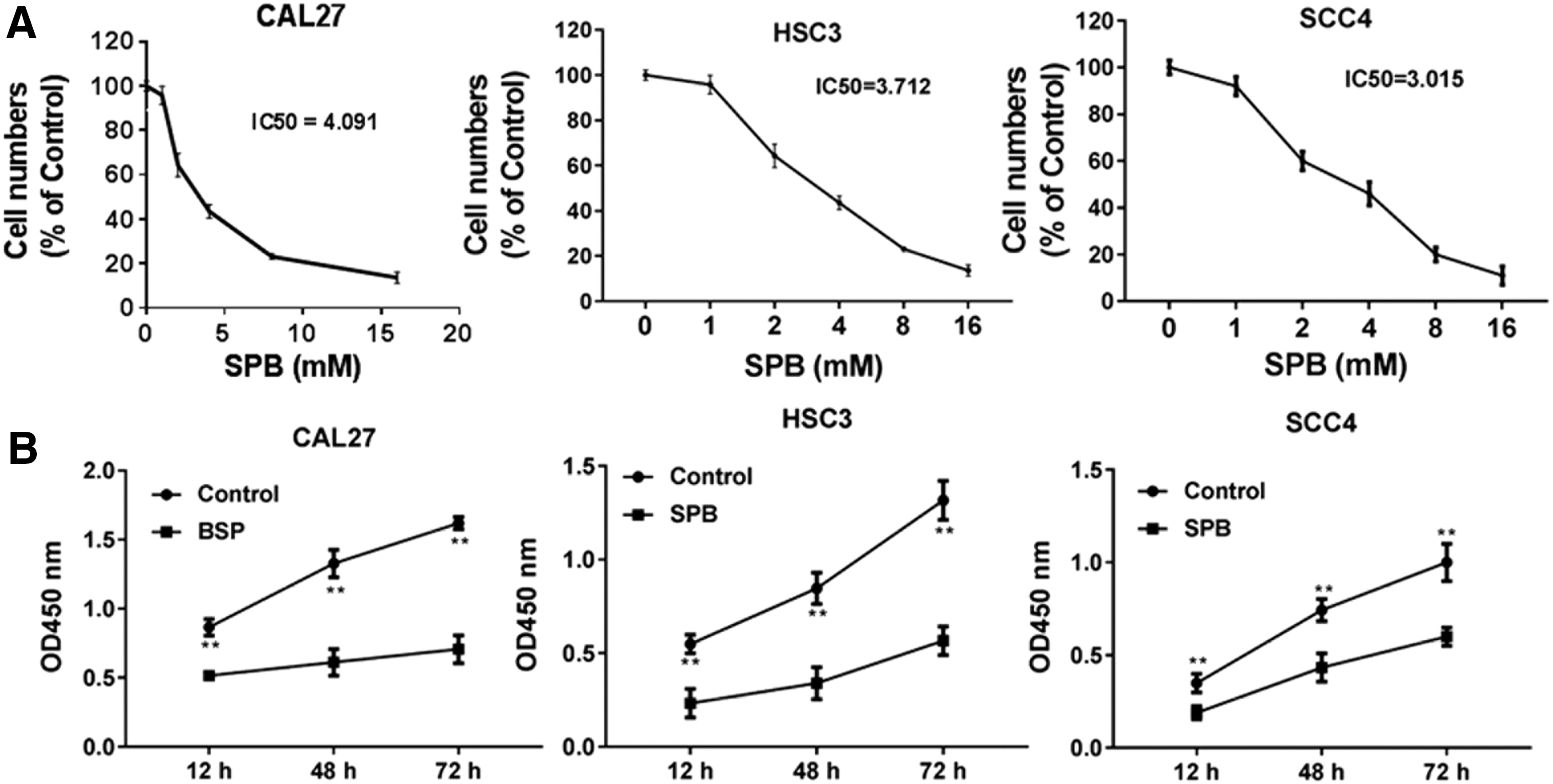
SPB inhibits the cell vitality of OSCC cells.
The effects of SPB on the apoptosis of CAL27, HSC3, and SCC4 in vitro
Considering the suppression of SPB on cell vitality of CAL27, HSC3, and SCC4 in vitro, the impacts of SPB on cell apoptosis were assessed. The results indicated that the treatment of SPB induced remarkable apoptosis of OSCC cell lines, which was consistent with the finding that SPB inhibited the cell vitality (Fig. 2A, B). Next, the mechanism of SPB-induced apoptosis was investigated. The apoptosis was involved in the antiapoptosis BCL-2 signal and proapoptosis caspase-3 signal. After treatment of SPB, the expression of BCL-2 was downregulated but the cleavage of caspase-3 was induced in CAL27, HSC3, and SCC4 cells (Fig. 2C). The data indicated that SPB exerts the proapoptosis role through activating the caspase-3 pathway.

SPB promotes the apoptosis of OSCC cells.
SPB inhibits the migration and invasion through regulation of TGF-β-related EMT phenotype
The role of SPB in the ability of migration and invasion of CAL27, HSC3, and SCC4 cells was further investigated. Transwell assay was performed and the results showed that the treatment of SPB significantly inhibited the ability of migration (Fig. 3A, B). Meanwhile, the expression of TGF-β in CAL27, HSC3, and SCC4 cells was decreased by SPB (Fig. 3C). The EMT phenotype including the epithelial marker E-cadherin and the mesenchymal marker N-cadherin was analyzed. The authors found that the treatment of SPB increased the expression of E-cadherin but reduced the expression of N-cadherin (Fig. 3D). These results suggested that SPB could inhibit the TGF-β-related EMT phenotype to impair the ability of migration and invasion of OSCC cells.

SPB impairs the migration of OSCC cells through EMT. The ability of migration was determined by
The treatment of SPB effectively suppresses the tumor growth in vivo
Since the antitumor function of SPB in vitro was identified, the efficiency of SPB for tumor inhibition in vivo was next evaluated. It was established that the xenograft model of human CAL27 and the mice bared with tumor were treated with SPB. The results showed that the administration of SPB could effectively inhibit tumor growth with decreased tumor volume (Fig. 4A). The EMT phenotype of tumor cell was also analyzed. The treatment of SPB, on one hand, repressed the expression of TGF-β in tumor tissues (Fig. 4B) and, on the other hand, increased the expression of E-cadherin but reduced the expression of N-cadherin in tumor tissues (Fig. 4C). Therefore, the data indicated that SPB has efficient antitumor effects on OSCC.
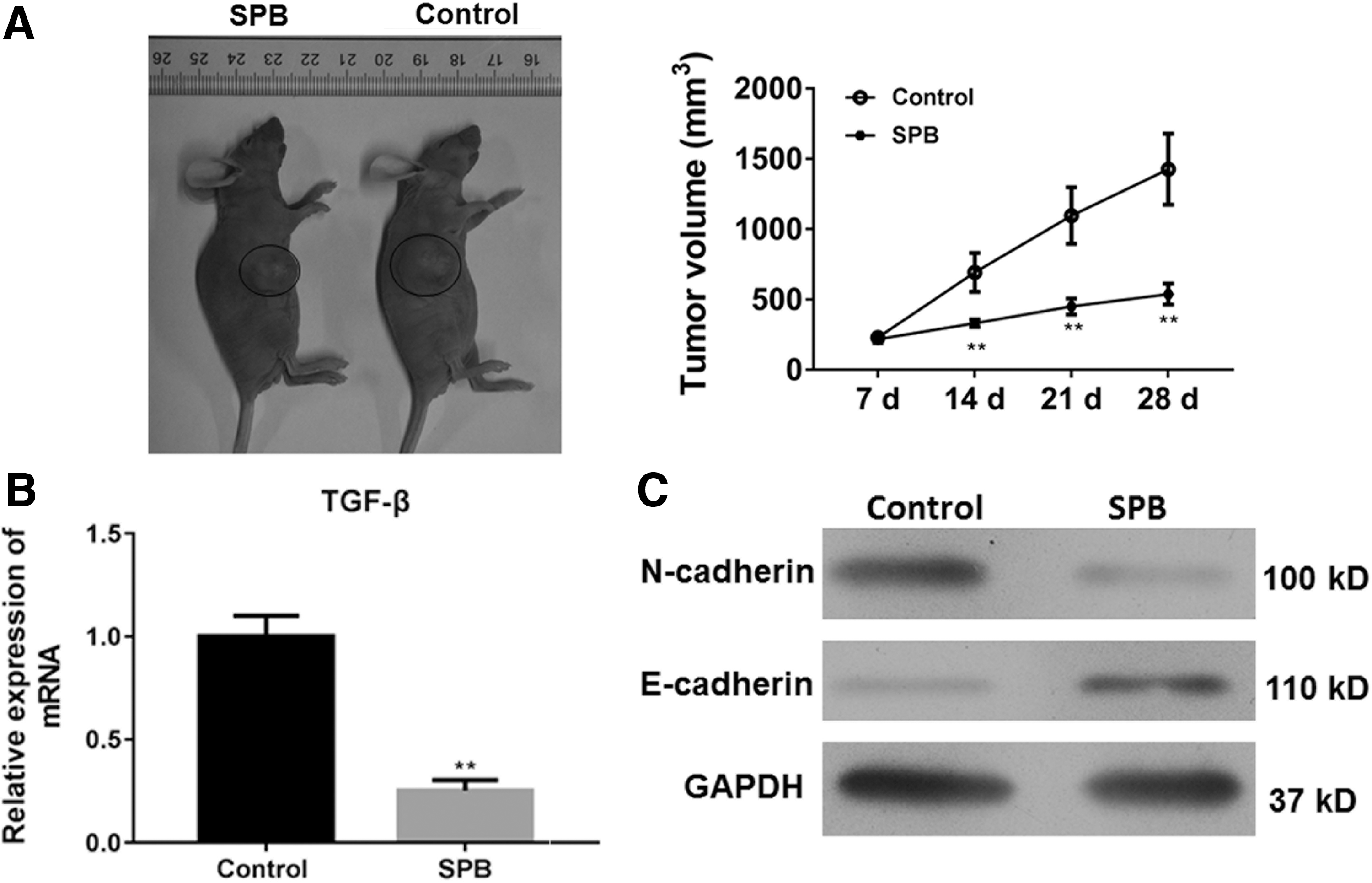
The antitumor role of SPB for OSCC cells in vivo. A total of 2 × 106 CAL27 cells were subcutaneously injected in rear flank of nude mice (4 per group) and 2 mmol/kg was administrated to mice.
Discussion
OSCC as a major subtype of head and neck squamous cell carcinoma (HNSCC) is characterized by many different pathological features to cancer at other sites in the head and neck region, including tobacco smoking and chewing, alcohol consumption, human papillomavirus (HPV) infection, and sun exposure. 3,17 The incidence and mortality rates for OSCC are higher in developing countries than in the developed world. The 5-year overall survival rate of OSCC in early stage is 80%, but that in late stage is only 20%. The combination of surgery and chemoradiotherapy is unsatisfied for the treatment of OSCC patients in late stage. 5,18 Thus, here the antitumor role of SPB was investigated, which is a salt of 4-PBA in the treatment of OSCC. It was found that SPB could promote tumor apoptosis and inhibit the TGF-β-related EMT phenotype of OSCC cells in vitro and in vivo.
Currently, different functions of phenylbutyrate have been discovered, such as ammonia scavenger, HDACi, an ER stress inhibitor, and regulator of mitochondrial biogenesis. 19 Considering these effects of SPB, mounting studies have shed light on its role in different diseases, including obesity, diabetes, acute renal failure, fibrosis, inflammation, and cancer. 9 Basseri et al. reported that, in adipogenesis of obesity, 4-PBA induced attenuation of adipogenesis with lipid accumulation and adiponectin secretion through suppression of ER stress. The treatment of SPB to mice fed a high-fat diet induced a significant reduction in weight gain and had reduced fat pad mass. 20 In aristolochic acid nephropathy, pretreatment with 4-PBA remarkably abrogated aristolochic acid I (AAI) induced ER stress and reduced activation of caspase-3-induced apoptosis. 21 Another study also showed the anti-inflammation role of phenylbutyrate in lipopolysaccharide (LPS)-induced lung inflammation; phenylbutyrate could impair the NF-κB and HIF-1α activities by inhibition of ER stress. 22 The roles of inflammation during carcinogenesis have been recognized for many years. Thus, the potential applications of SPB have been proposed in the treatment of cancer.
In this study, it was found that the treatment of SPB promoted cell apoptosis, which was dependent on BCL-2 and caspase-3, and inhibited the TGF-β-related EMT phenotype of OSCC cells. The results indicated that SPB could be used for tumor therapy of OSCC. The antitumor roles of SPB in other types of cancers have also been reported. Maslak et al. combined the SPB and 5-azacytidine for the treatment of patients with acute myeloid leukemia or myelodysplastic syndrome, and this combined therapy was clinically feasible with acceptable toxicity and measurable biologic and clinical outcomes. 23 For the patients with refractory solid tumors, a clinical study also combined 5-azacytidine with SPB that was generally well tolerated and safe, although on evidence for clinical benefit was found in this study. 24 Besides, Chen et al. reported that combinatory treatment with SPB and trastuzumab significantly abrogated the cell proliferation and apoptosis of SKBR3 breast cancer cells through upregulation of the cyclin-dependent kinase inhibitor p27kip1. 25 In HNSCC, phenylbutyrate improved the sensitivity of cisplatin for cell cycle arrest by inhibiting the FA/BRCA pathway in cancer cells. 26 In addition, the treatment of SPB alone also showed the tumor elimination in prostate cancer; SPB treatment impaired the cell vitality and attenuated the colony formation and migration abilities in prostate cancer cells, but the mechanism of SPB-regulated migration was unclear. 12 Here it was found that SPB could downregulate the expression of EMT inducer TGF-β, leading to the inhibition of TGF-β-related EMT phenotype (increased expression of E-cadherin but reduced expression of N-cadherin). The tumor suppressive role of SPB in vivo was demonstrated in prostate cancer, and it was also shown that SPB delayed the tumor growth in vivo and inhibited the EMT phenotype of tumor cell.
Collectively, in this study, a possibility of SPB for the treatment of OSCC was proposed. It was found that SPB impaired the cell vitality in a dose- and time-dependent manner, and induced the cleavage of caspase-3 for apoptosis. Importantly, SPB decreased the expression of TGF-β to inhibit the EMT phenotype in vitro and in vivo.
Footnotes
Acknowledgment
This study was supported by Huzhou University.
Disclosure Statement
No competing financial interests exist.