
Abstract
Primary hepatocellular carcinoma (HCC) is one of the most common malignant tumors. At present, the molecular mechanism of HCC remains unclear. A recent circular RNA (circRNA) profiling study showed that circRBM23 expression was upregulated in HCC tissues. Therefore, in this study, the impact of circRBM23 during the progression of HCC was evaluated. The expression levels of circRBM23 and miR-138 in HCC tissues and HCC cell lines were determined by RT-PCR and the results indicated that circRBM23 expression was increased in the HCC tissues and HCC cell lines, whereas miR-138 expression was decreased. An upregulation of circRBM23 expression in HCC cells was shown to increase cell viability, and also increased the ability of cells to migrate. Downregulation of circRBM23 was found to decrease cell viability, proliferation, and migration, and promote the expression of miR-138 and its related target genes, vimentin, and CCND3. Moreover, miR-138 was found to regulate HCC cell viability and migration, and the levels of vimentin and CCND3 protein expression were found to be inversely correlated with those of miR-138 expression. The downregulation of circRBM23 in HCC tissues can regulate the miR-138-mediated signal pathway by promoting miR-138 expression. The results in vivo demonstrated that circRBM23 is required for the tumorigenesis with downregulation of tumor suppressor miR-138. These data indicated that upregulated circRBM23 functioned as oncogene in HCC through regulating the tumor suppressor miR-138.
Introduction
The Encyclopedia of DNA Elements (ENCODE) project revealed that 70%–90% of sequences in the human genome have the potential for transcription into mRNA, but <2% of the sequences are capable of encoding proteins. 1 Those noncoding sequences can form noncoding RNA molecules through their transcription, and thereby regulate biogenic activities. 2,3 Thus, there exists a complex RNA–RNA network relationship between the different types of RNAs that allows them to regulate each other's expression level. 3,4 Based on this information, biologists have proposed the hypothesis of competing endogenous RNA (ceRNA). 5 The transcript of many RNAs can integrate with the response element microRNA (miRNA) response elements (MRE) of microRNA (miRNA), and thereby reduce the inhibitory effect of miRNAs on their target genes and improve their expression level. The RNA transcripts and their target genes are referred to as ceRNA. 5,6 This ceRNA-based method of regulation has been shown to affect several different biogenic activities; however, few studies have investigated the genes that exert ceRNA effects on tumors. 4,6 Primary hepatocellular carcinoma (HCC) is one of the most common malignant tumors, and has a high rate of mortality. Furthermore, its mortality rate among men ranks second among all tumor-related mortality rates. 7 At present, the molecular mechanism of HCC remains unclear. However, current studies of genes related to HCC have primarily focused on those that encode for miRNAs, long noncoding RNAs, circular RNAs (circRNAs), and mRNAs. Recently, studies on how ceRNA regulation of circRNA–mRNA affects the occurrence and development of HCC have received increased attention.
miRNA is a type of small single-stranded noncoding RNA with a length of 22 nt, and plays a key role in regulating ceRNA. 4 Mature miRNA can be rapidly integrated into an miRNA-induced silencing complex and inhibit RNA translation through its complementary integration with the mRNA's 3′ noncoding area, and thus negatively regulate mRNA expression. 4,8 It is now known that aberrant miRNA expression is a key factor leading to the development of several tumor types, including HCC. 4,8 Because certain miRNAs participate in regulating cell cycle progression, their aberrant expression may lead to cell cycle dysfunction and induce tumor formation. 4,8,9 Numerous studies concerning how cell cycle-related miRNAs may regulate HCC cells have proven that aberrant miRNA expression plays a vital role in tumor formation. 9 –11 circRNA is a type of RNA molecule with important biological functions, but has long been neglected. 12 The first circRNA was found in viruses in 1976. 13 circRNA is typically a circRNA consisting of exons. 12 Recent studies have shown that circRNA contains numerous MREs of miRNA and can exert a ceRNA function, and thus is a new member of the ceRNA family. 14 Owing to continuing studies of circRNA, its role in the occurrence of tumors has gradually become acknowledged by researchers. The most recent study showed that hsa-circ-0005075 can interact with the miRNAs of several target genes, including miR-23b-5p, miR-93-3p, miR-581, and miR-23a-5p, to remove or mitigate their inhibitory effect, and thus promote the progression of HCC. 15 By testing miRNA expression in HCC cells, Wang et al. 16 discovered 10 upregulated miRNAs and 10 downregulated miRNAs, among which, miR-138 expression was downregulated in 77.8% of HCC tissues when compared with miR-138 expression in adjacent nontumor tissues. In contrast, a recent circRNA profiling study showed that circRBM23 expression was upregulated in HCC tissues. 17 Therefore, in this study, it was hypothesized that downregulation of circRBM23 in HCC tissues can regulate the miR-138-mediated signaling pathway by promoting miR-138 expression, and further promote tumorigenesis.
Materials and Methods
Clinical sample collection
Eleven specimens of histopathologically confirmed HCC tissue and 11 specimens of adjacent normal liver tissue were obtained from the First Affiliated Hospital of Zhengzhou University, and stored at −80°C until use. None of the tissue donors had received any additional treatment before surgery. The study protocol was approved by The Medical Ethics Commission of the First Affiliated Hospital of Zhengzhou University, and all tissue donors provided their written informed consent.
Cell culture
HCC cell lines HepG2, HHCC, HUH7, and BEL-7402, and the normal liver cell line HL-7702, were obtained from ATCC (Manassas, VA). The cells were cultured in Dulbecco's modified Eagle's medium (Gibco, Waltham, MA) containing 10% fetal bovine serum in a humidified atmosphere consisting of 5% CO2 at 37°C.
Real-time quantitative PCR
Total RNA was extracted from each cell line and each sample of HCC tissue and paired paracancerous liver tissue using SYBR Premix Ex TaqTM II (TaKaRa, China), and then amplified using the following protocol: 95°C for 5 min, followed by 40 cycles of 95°C for 5 s, and 60°C for 60 s. The following primers were used for circRBM23: 5′-TATCCTAGCAATACCACCAGCA, 3′-CATGGCCTCAATCACTATGTC-5′. The primers used for miR-138 were 5′-ACACTCCAGCTGGGAGCTGGTGTTGTGAATCA-3′ and 3′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCGGCCTG-5′ (Sangon Biotech, Shanghai, China). The associated gene expression levels in the HCC cell lines or HCC tissues were compared with those in the normal liver cell line or paired samples of paracancerous liver tissue, respectively. All results represent the mean ± standard deviation (SD) of data obtained from three independent experiments.
Cell transfection
circRBM23 siRNA, Vector-circRBM23, and the miR-138 mimics were designed and synthesized by RiboBio (Guangzhou, China). The miR-138 mimics sequence was AGCTGGTGTTGTGAATCAGGCCG. Before transfection, HCC cells were seeded into 6-well plates at a density of 6 × 104 cells/cm2 and incubated overnight to achieve 40%–50% confluence. After incubation, Lipofectamine 2000 (Invitrogen, Carlsbad, CA) was used to transfect the cells with circRBM23-targeted siRNA according to the manufacturer's instructions. To force miR-138 overexpression, the cells were transfected with the miR-138 mimics (RiboBio) using a Ribo FECT™ CP Transfection Kit (RiboBio). After transfection, the cells were continuously cultured for 48 h before harvesting.
Cell Counting Kit-8 assay
Cell viability was determined using a Cell Counting Kit-8 (CCK-8; Dojindo, Japan). Approximately 1 × 104 HCC cells were seeded into each well of a 96-well plate and cultured for 72 h. The number of viable cells was determined at 24, 48, and 72 h after seeding by determining optical density values at 450 nm. The data were then used to plot a cell proliferation curve.
EdU assay
A Cell-Light™ EdU assay kit (RiboBio) was used to further assess cell viability. The viability of cells subjected to different treatments was assessed using a FACSCanto II cytometer (BD Biosciences, Germany).
Cell cycle analysis
Cell cycle stages were identified using propidium iodide ReadyProbes® Reagent (Thermo Fisher Scientific, Waltham, MA). After transfection, the cells were collected, washed twice with cold PBS, and then stained with propidium iodide reagent (two drops per 106 cells in 1 mL) for 30 min at room temperature. The cells were then examined using a FACSCanto II cytometer (BD Biosciences).
Wound healing assay
To evaluate cell migration ability, a 200-mL pipette tip was used to create wounds on the surface of cultured HepG2 and Huh7 cells that had reached 90% confluence. Next, the surface was rinsed with culture medium to remove any free-floating cells and debris, and the cells were incubated for 24 h at 37°C. After incubation, the scrape lines were observed and used to assess wound healing. All results represent the mean ± SD of data obtained from three independent experiments.
Cell migration assay
A 24-well Boyden chamber with a polycarbonate membrane (Corning, Corning, NY) was used to detect cell migration. The lower compartment was filled with cell culture medium, and 1 × 105 cells were added into the upper compartment of the chamber. After incubation at 37°C for 24 h, the migrated cells on the lower surface of the chamber were fixed with methanol and stained with 0.1% crystal violet. The number of migrated cells was counted under a light microscope.
Western blot studies
Cells were harvested and lysed using RIPA buffer (Sigma-Aldrich, St. Louis, MO). BCA reagent (Thermo Fisher Scientific) was used to determine the total protein concentration. Aliquots of protein (20 μg) were separated by 10% SDS-PAGE, and the protein bands were electrotransferred onto a PVDF membrane (Merck Millipore, Billerica, MA). Next, the membrane was successively incubated with anti-CCND3 and vimentin primary antibodies (1:1000; Abcam, Cambridge, MA) and a goat antirabbit IgG-HRP secondary antibody (1:2000; Abcam), after which, immunostaining was detected using an enhanced chemiluminescence kit (Thermo Fisher Scientific). Immunostaining of GAPDH was used as an internal reference.
Tumor model
The HepG2 cells were transfected with lentivirus vector of siRNA/overexpression-circRBM23 or negative control. The xenograft model of conditional human HepG2 cells was established. A total of 2 × 106 tumor cells were subcutaneously injected in rear flank of nude mice (6 per group). The tumor sizes were measured 3 d apart and the tumor volumes were calculated: V (cm3) = width2 (cm2) × length (cm)/2.
Statistical analysis
All data analyses were performed using SPSS for Windows, Version 13 software (SPSS, Inc., Chicago, IL). Analysis of variance was used to determine correlations between different groups. Differences between values for circRBM23 were analyzed using the paired t-test. p-Values <0.05 were considered statistically significant. Each experiment was repeated at least three times, and results are presented as the mean ± SD.
Results
Expression of circRBM23and miR-138 in liver tissues and different HCC cell lines
The levels of circRBM23 and miR-138 expression in HCC tissues and paired adjacent liver tissues were analyzed by quantitative RT-PCR. As shown in Figure 1A, the levels of circRBM23 expression were significantly higher in the HCC tissues than in the adjacent liver tissues (p < 0.01), whereas miR-138 expression was significantly decreased in the HCC tissues when compared with the adjacent liver tissues (p < 0.01; Fig. 1B). In addition, the levels of circRBM23 and miR-138 expression were also detected in the different HCC cell lines (HepG2, HHCC, HUH7, and BEL-7402) and the normal liver cell line, HL-7702. The results of this study revealed consistently increased levels of circRBM23 expression and decreased levels of miR-138 expression in all of the HCC cell lines. Moreover, all of the differences were statistically significant when compared with expression levels in the normal liver cell line (p < 0.01; Fig. 1C, D).

Expression of miR-138 and circRBM23 in HCC cell lines and HCC tissues.
Downregulation of circRBM23 decreased cell viability and blocked cell cycle progression
To clarify the role of circRBM23 in HCC cells, circRBM23 siRNA was transfected to HepG2 and Huh7 cells. As shown in Figure 2A, such transfection significantly reduced the level of circRBM23 expression (p < 0.01 vs. the negative control group). The viability of circRBM23 siRNA-transfected HepG2 and Huh7 cells was next examined. CCK-8 assay results showed that the siRNA-transfected cells had become significantly less viable than control cells at 72 h post-transfection (p < 0.01; Fig. 2B). This result was further validated by the EdU assay, in which the number of EdU-positive cells was dramatically reduced in the siRNA-transfected group (Fig. 2C). Furthermore, when compared with control cells, circRBM23 siRNA-transfected cells had lower percentages of S phase cells (p < 0.01; Fig. 2D). These findings demonstrated that circRBM23 siRNA had most likely blocked cell cycle progression from the G0/G1 phase to the S phase.
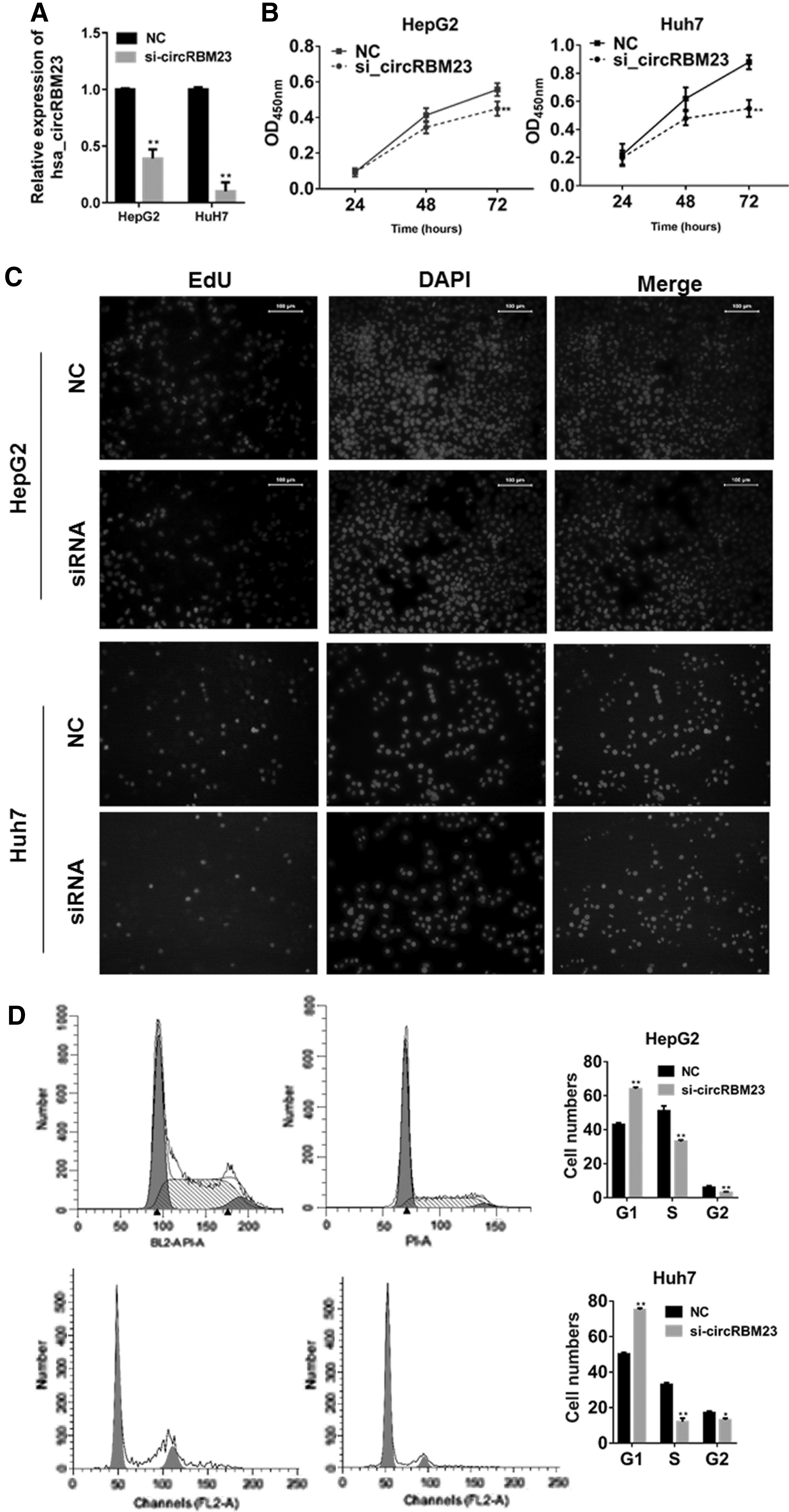
Downregulation of circRBM23 decreased HepG2 and HUH7 cell viability and blocked cell cycle progression.
Downregulation of circRBM23 inhibited HepG2 and Huh7 cell migration
HCC cell migration and invasion, which are distinctive features of malignant cell behavior, were further investigated. As shown by results of this wound healing assay, the reduced expression of circRBM23 in HepG2 and Huh7 cells significantly inhibited wound closure when compared with the wound healing shown by control cells (p < 0.01; Fig. 3A), which indicated that cell migration was inhibited. To further confirm this result, the effects of circRBM23 siRNA transfection on cell migration were investigated using Transwell chambers. The results showed that cells derived from the circRBM23 siRNA-transfected group had lower migration capabilities than cells in the control group (p < 0.01; Fig. 3B). These data indicated that downregulation of circRBM23 inhibited cell migration in vitro.
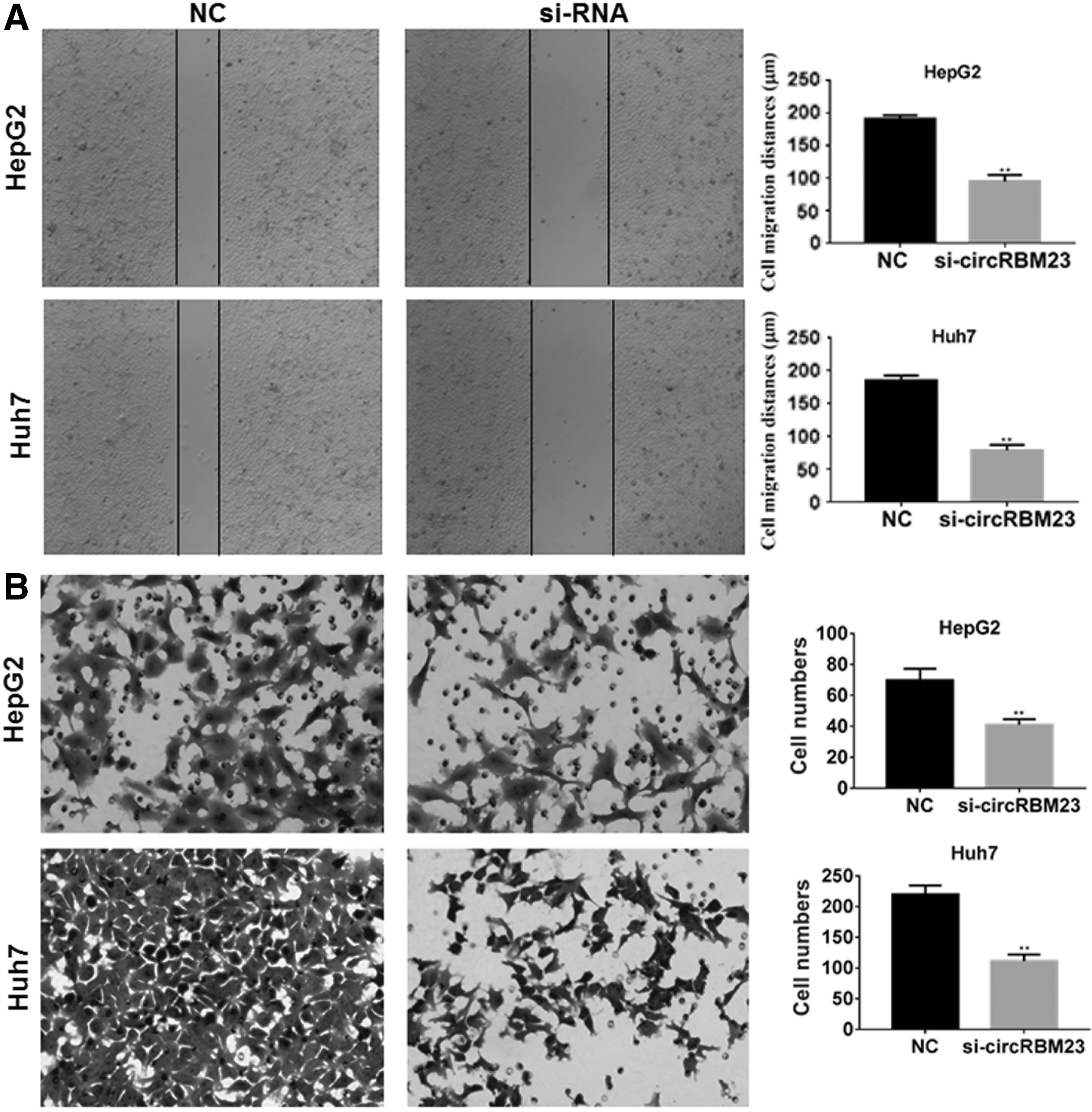
Downregulation of circRBM23 inhibited HepG2 and HUH7 cell migration.
Downregulation of circRBM23 promoted miR-138 expression
Interestingly, bioinformatic analysis revealed putative complementary sequences for miR-138 in human circRBM23, and predicted tumor suppressor miR-138 binding sites were found (
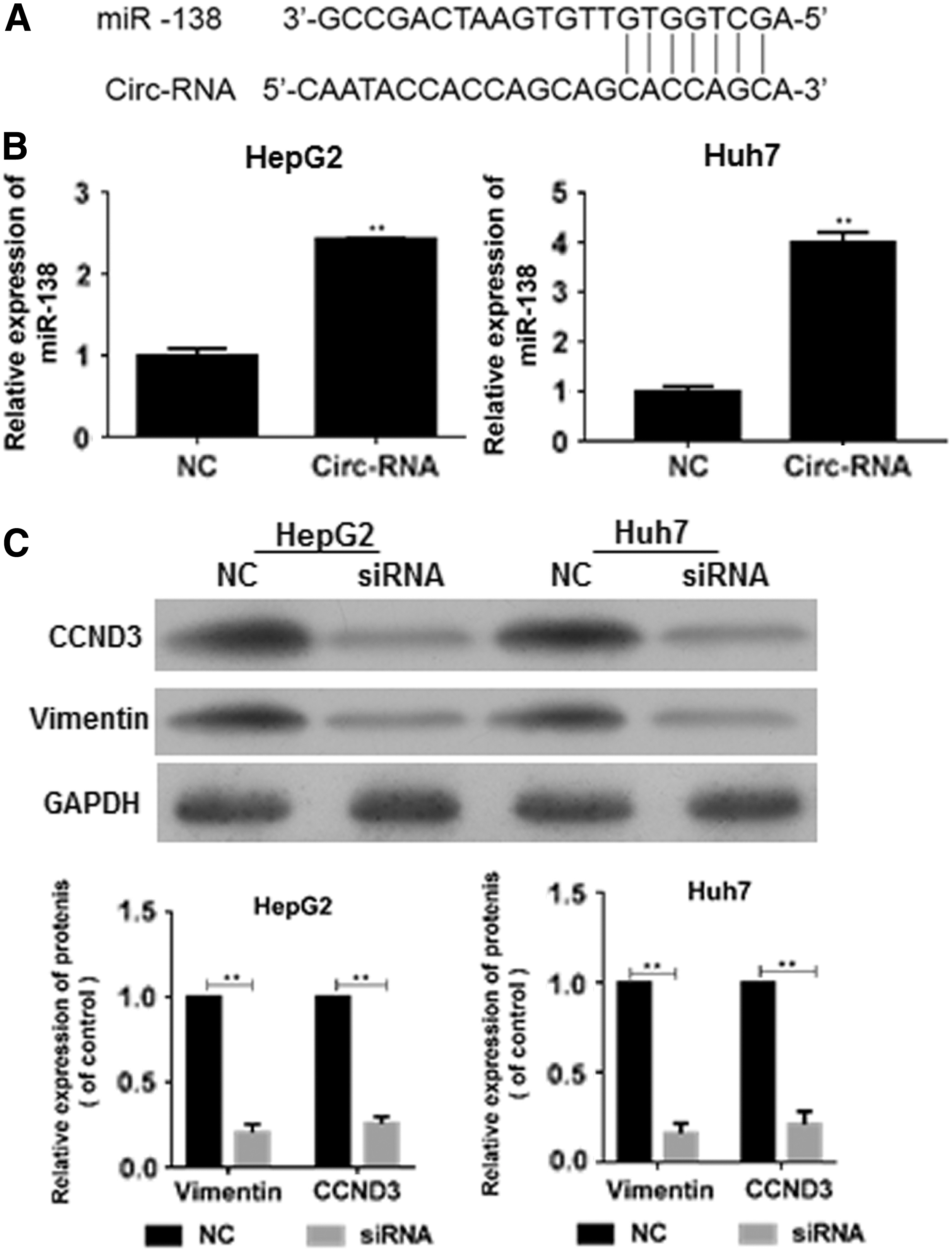
Downregulation of circRBM23 promoted miR-138 expression.
Effects of miRNA-138/circRBM23 on HepG2 and Huh7 cell viability and migration
To further study the role of microRNA-138 in hepatocarcinogenesis, pcDNA-circRBM23 and miR-138 mimic or negative control was cotransfected into HCC cells, after which the RT-PCR analyses showed that miR-138 became significantly overexpressed and suppressed by overexpressed circRBM23, respectively (p < 0.01 vs. the control group; Fig. 5A). Next, the viability of over-circRBM23+ miR-138 mimic transfected HepG2 and Huh7 cells was examined. This study's CCK-8 assay results show that HepG2 and Huh7 cells transfected with the miR-138 mimic were even less viable than HepG2 and Huh7 cells transfected with NC alone. On the other hand, HepG2 and Huh7 cells transfected with the miR-138 mimic and overexpressed circRBM23 displayed significantly enhanced cell vitality at more 72 h post-transfection (p < 0.01; Fig. 5B). The effects of microRNA-138 on HepG2 and Huh7 cell migration were next studied. In both the wound healing and Transwell chamber assays, miR-138 mimic transfection significantly inhibited wound closure, and the transfected cells displayed a lower migration capability, which was restored by overexpression of circRBM23 (p < 0.01; Fig. 5C, D). These data indicate that miR-138 can regulate HepG2 and Huh7 cell viability and migration. Furthermore, Western blot results consistently showed that the levels of vimentin and CCND3 protein expression were inhibited by miR-138 expression but were restored by circRBM23 (Fig. 5E).

Effects of circRBM23/microRNA-138 on HepG2 and Huh7 cell viability and migration.
circRBM23 is required for the growth of HCC in vivo
The xenograft model of human HepG2 cell line was established and nude mice were treated with conditional tumor cells with circRBM23 overexpression or knockdown. It was found that circRBM23 promoted the tumor growth with reduced expression of tumor suppressor miR-138 in tumor tissues, which could be restored by circRBM23 knockdown. (Fig. 6A, B). The expression levels of vimentin and CCND3 were determined and found that circRBM23 can regulate the miR-138-mediated signal pathway in vivo (Fig. 6C).
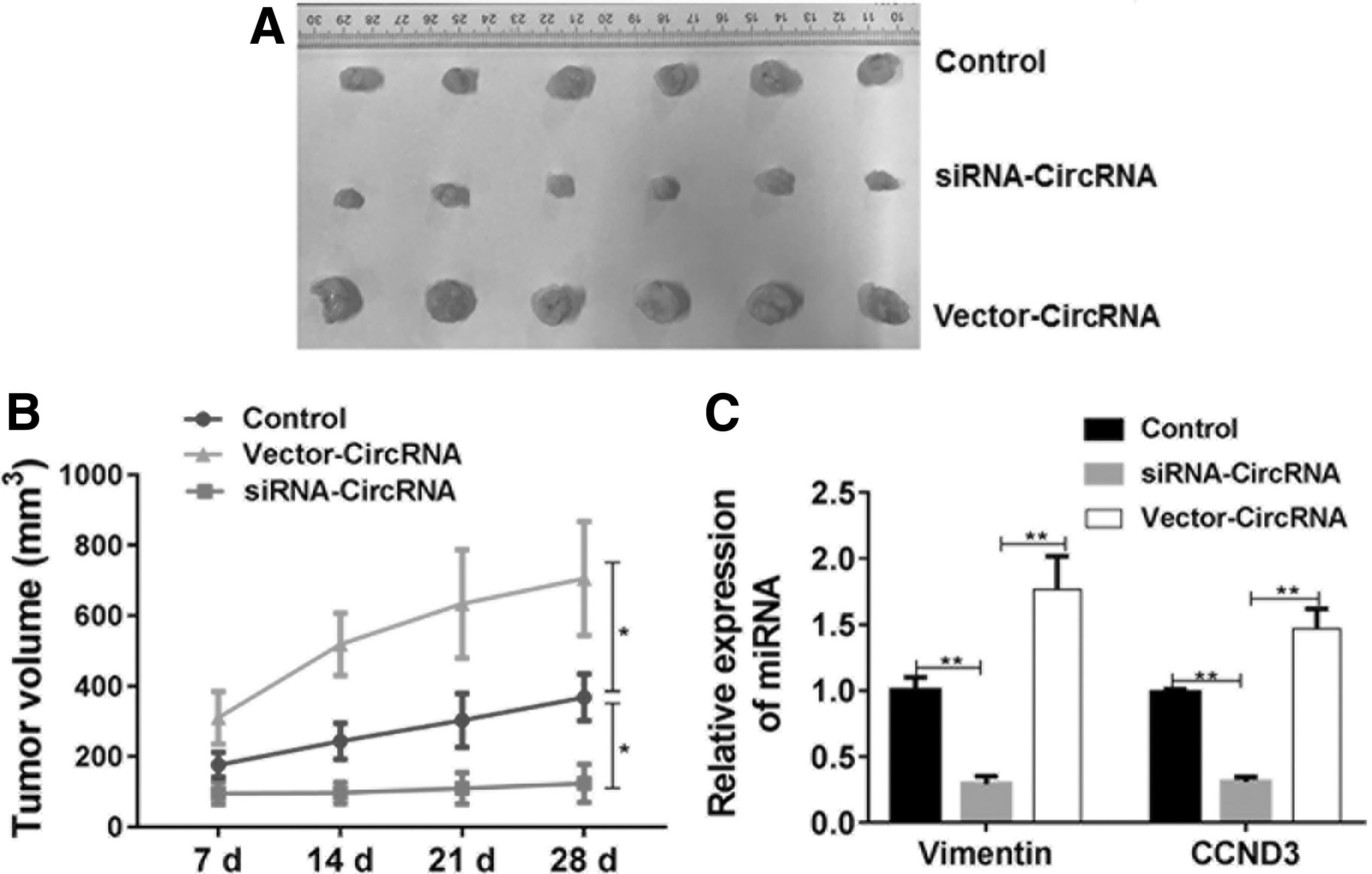
circRBM23 promotes the tumor growth of HCC in vivo. The xenograft model of human HepG2 cell line was established and the nude mice were treated with conditional tumor cells with circRBM23 overexpression or knockdown.
Discussion
The ceRNA hypothesis proposes a complex ceRNA network (ceRNET) formed by different types of RNAs under the mediation of miRNAs. 5 Thus changing the expression level of a specific ceRNA component should lead to obvious changes in content of the entire ceRNET. 4,5 Widely found in eukaryotes, miRNA is a type of endogenous single-stranded small molecular RNA that has been highly conservative during evolution, with the expression characteristic of spatial and temporal specificity in cells. 4,9 Although miRNA does not encode for proteins, its specific complementation with its target gene sequences and ability to regulate the post-transcriptional expression of those genes allow it to participate in various biological processes, such as cell proliferation, differentiation, and apoptosis. 4,9 miRNA is closely related to tumor development, and genome instability is a basic feature of malignant tumors. 18 Thus, tumor development is often accompanied by chromosomal amplification or deletion, resulting in the excessive activation of oncogenes or inactivation of antioncogenes. 19 Many miRNA molecules are located in chromosome variation areas, which are closely related to tumors. Thus, subtle changes in an miRNA level and its regulated targets may lead to major changes in cells. 20 Studies have shown that as an oncogene or antioncogene, miRNA can promote the occurrence of tumors by decreasing the expression of antioncogenes or increasing the expression of oncogenes. 19,21 Therefore, some investigators currently support the concept of oncomirs; that is, if the expression of miRNAs in tumors is upregulated, those miRNAs function as oncogenes that affect the differentiation and apoptosis pathways of cells by negatively regulating tumor antioncogenes so as to promote tumor development. In contrast, the expression of other miRNAs becomes downregulated, and those miRNAs function as antioncogenes. 22 A previous study reported that the expression levels of miR-138 in 77.8% of HCC tissues were downregulated when compared with their expression levels in adjacent nontumor tissues. 16 In this study, decreased levels of miR-138 expression not only in HCC tissues but also in the HCC cell lines were found. Moreover, miR-138 was found to regulate HepG2 cell viability and migration, and the levels of vimentin and CCND3 protein expression were inversely correlated with miR-138 expression. All of these findings indicate that miR-138 functions as an antioncogene in HCC.
As a ceRNA, circRNA functions as a molecular sponge for miRNA. 12 The ceRNET is complex and exquisite, and any tiny disorder can change gene expression and lead to tumor development. 4 However, as an important member of the ceRNET, circRNA is indispensable for maintaining the network's balance. Furthermore, by studying the miRNA sponge role of circRNAs, it may be possible to explain the molecular mechanisms underlying the development of certain tumors. 23 Bachmayr-Heyda et al. 24 discovered that the ratio of circRNA to its corresponding linear RNA in colorectal cancer (CRC) tissues was lower than those in normal tissues. Moreover, that ratio was even lower in tumor cell lines, suggesting that circRNA levels are negatively correlated with cell proliferation. Likewise, the circRNAs with decreased expression in tumor tissues include hsa_circ_001988. The levels of hsa_circ_001988 expression in the tumor tissues of CRC patients are related to the differentiation level of the tumor cells, as well as whether the tumor has invaded the surrounding nerves. 25 However, not all studies have reported a decreased level of circRNA expression in cancer tissues. A study by Xie et al. 26 showed that circRNA_001569 expression was increased in CRC tissues. Those investigators also found that among circ_001569-expressing SW480 and HCT116 cells, the number of cells in their S and G2/M phases was increased, whereas the number of cells in their G1/G0 phase was decreased. Furthermore, the number of apoptotic cells was also decreased, suggesting that the increase in tumor invasiveness caused by circRNA_001569 partly results from the removal of factors that inhibit cell progression and apoptosis. 26 Moreover, circRNA_001569 has also been shown to act on miR-145 to upregulate expression of the miR-145-E2F5 target genes, BAG4 and FMNL2. 26 In this study, increased levels of circRBM23 expression in the HCC tissues and all HCC cell lines were consistently found. This upregulation of circRBM23 in HCC cells can increase their viability by removing factors that inhibit cell cycle progression. Moreover, downregulation of circRBM23 was found to promote the expression of miR-138 and its related target genes, vimentin and CCND3. Distant metastases and invasion of HCC are the main causes for death among liver cancer patients. 27 Therefore, research on how to diminish the invasion and transfer functions of HCC is of great clinical importance. In this study, upregulation of circRBM23 in HCC cells increased their migration ability, which was reduced by the transfection of circRBM23 siRNA, and then restored by the cotransfection of miR-138 inhibitors. Based on these results, it was proposed that downregulation of circRBM23 in HCC tissues can regulate the miR-138-mediated signal pathway by promoting miR-138 expression, and further promote tumorigenesis.
After conducting numerous studies, investigators have gained a preliminary understanding of the basic characteristics and biological functions of miRNA. They now realize that many miRNAs contribute to the progression of HCC and also regulate multiple biological processes. 28,29 However, most HCC-related miRNA studies have primarily focused on the newly discovered relationships between miRNAs and their target genes, whereas few studies have examined the relationship between miRNAs and circRNA. 28,29 Accumulating experimental evidence suggests that circRNA is widely used by cells as a gene regulation tool. Also, through its mechanism of absorbing miRNA and other unclarified mechanisms, ceRNA represents a new approach for regulating and controlling gene expression. Furthermore, its functions increase the complexity of such gene regulation mechanisms. 3,4,8,23 The unique ceRNA characteristic of circRNA opens a new frontier for the development of new medicines. By utilizing circRNA, the activity of a specific miRNA can be inhibited and another specific miRNA can also be transported into target tissues. Also, the high tissue specificity and stability of circRNA are likely to make it a good biomarker for certain diseases. Therefore, the protumor role of circRBM23 indicated the potential prognostic and therapeutic value in HCC.
Footnotes
Acknowledgment
The authors thank all who have supported and participated in this research.
Disclosure Statement
There are no existing financial conflicts.