
Abstract
Cancer Biotherapy and Radiopharmaceuticals
officially retracts the article entitled, “Downregulation of Long Noncoding RNA HOTAIR and EZH2 Induces Apoptosis and Inhibits Proliferation, Invasion, and Migration of Human Breast Cancer Cells” by Lu Han, Hai-Chao Zhang, Li Li, Cai-Xia Li, Xu Di, and Xin Qu (Cancer Biother Radiopharm. 2018;33(6);241–251; doi: 10.1089/cbr.2017.2432) due to inconsistencies in some of the images in the article. These inconsistencies were originally questioned in a comment on the PubPeer platform in 20201 but were then reported as being part of a large paper mill in September 2021.2
The corresponding author, Dr. Lu Han, did not respond to requests for the raw data as it pertained to the work for further examination.
Submission of manuscripts from a paper mill is a violation of the journal's standard protocols and is considered an infraction against the rigorous standards of scientific publishing.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves and does not tolerate any misconduct.
References
1. PubPeer.
2. Smut C. The scientific sea of miR- and exosome-related knowledge. For Better Science. Sept 22, 2021;
Background
The incidence of breast cancer among females worldwide represents the second leading cause of cancer-related deaths among women. 1,2 According to statistics reported by the American Cancer Society, during the year 2014, ∼233,000 new cases accompanied by 40,000 deaths have been estimated to have occurred due to breast cancer. 3 Breast cancer remains a significant stumbling block from many perspectives, including clinical and societal, posing a major scientific challenge, largely due to metastasis that accounts for the foremost cause of breast cancer deaths worldwide. 4,5 Therefore, it is of great significance that reliable cost-effective biomarkers continue to be discovered and developed to evaluate the prognosis of patients with metastatic breast cancer. 6
Genome-wide experiments have discovered that mammalian genomes are universally transcribed, producing a large number of regulatory nonprotein-coding RNAs (ncRNAs). 7,8 A previous study highlighted there to be a deregulation of ncRNA gene expression in human diseases, including that of cancer, 9 which provides support to the notion that investigations into ncRNAs could be a promising area for breast cancer treatment.
As a type of RNA molecule, ncRNAs have an array of important functions and are abundant in number. Furthermore, ncRNAs are not translated into a variety of proteins and can generally be assorted into two classes based on their sizes: long ncRNAs (lncRNAs) and small ncRNAs. 7,10 A principal fraction of the transcriptome in higher organisms comprises an extensive repertoire of lncRNAs, which are expressed in a developmental stage-specific manner. 11 LncRNAs are key factors in a variety of biological processes, including that of gene regulation, genomic imprinting, genome packaging, dosage compensation, and chromatin organization. 12,13 HOTAIR (HOX transcript antisense intergenic RNA) is a long intergenic ncRNA that suppresses gene expression in an epigenetic manner through the enhancer of a histone methyltransferase (HMT), enhancer of zeste homologue 2 (EZH2). 14
An investigation into the effects of lncRNAs on the development of breast cancer was previously conducted, which revealed that HOTAIR could promote metastasis and reprogram the chromatin organization of cancer cells. 15 A recent study highlighted an association between the upregulation of HOTAIR in metastatic tissue and the functionality of HOTAIR, suggesting it could promote the invasion and motility of melanoma cells. 16 Furthermore, the lncRNA HOTAIR has at times been considered to be a biomarker of poor prognosis in various types of malignant tumors. 17 Although HOTAIR has been implicated in several cancers, its involvement in breast cancer is yet to be properly elucidated. Thus, the aim of this study was to investigate the effects of lncRNA HOTAIR and EZH2 on the proliferation, invasion, migration, and apoptosis of human breast cancer cells.
Materials and Methods
Cell culture
Normal human mammary epithelial cell line HBL-100 and breast cancer cell lines MCF-7, MDA-MB-231, and SKBR-3 were all purchased from the cell bank of Chinese Academy of Sciences (Shanghai, China) and cultured in Dulbecco's minimum essential medium (DMEM, 12400-024; Gibco Company, Grand Island, NY) consisting of 10% fetal bovine serum (FBS, 11012-8611; Gibco Company, Hangzhou, China) and 1% double antibody (100 U/L penicillin and 100 mg/L streptomycin, 15140-122; Gibco Company). All the cell lines were cultured in 5% CO2 incubators at 37°C. The cells were passaged every 2–3 d and cells in logarithmic growth phase were eventually selected for further experiments.
Cell transfection and grouping
Breast cancer MCF-7, MDA-MB-231, and SKBR-3 cells in the logarithmic growth phase were inoculated into a six-well plate; after 24 h had elapsed, the cells were continually cultured in an antibiotic-free medium. When cell density had reached 40%–60%, the medium was replaced with a serum-free Opti-MEM culture medium. When cell density reached 70%, the transfection process was performed accordingly. The complex mixed by transfection reagent and 200 μL small interfering RNA (siRNA) was prepared based on the criteria of Lipofectamine 2000 transfection kit (11668019; Invitrogen Thermo Fisher Scientific, Waltham) and added to the medium. Transfected siRNA free of interference was regarded as the control.
After transfection, the cells were placed in a 5% CO2 incubator at 37°C for 6 h and then cultured in a 1.5 mL normal medium with a serum comprising 10% FBS. The cells were subsequently assigned into the blank group (not transfected), negative control group (NC group, transfected with nonspecific siRNA), siRNA-HOTAIR group, siRNA-EZH2 group, and siRNA-HOTAIR+siRNA-EZH2 group.
The primer sequences of siRNA were synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The upstream sequence of siRNA-NC was 5′-UUCUCCGAACGUGUCACGUUU-3′, and the downstream sequence was 5′-ACGUGACACGUUCGGAGAAUU-3′; The upstream sequence of siRNA-HOTAIR was 5′-UUUUCUACCAGGUCGGUAC-3′, and the downstream sequence was 5′-AAUUCUUAAAUUGGGCUGG-3′. The upstream sequence of siEZH2 was 5′-TTCATGCAACACCCAACACT-3′, and the downstream sequence was 5′-GAGAGCAGCAGCAAACTCCT-3′.
Reverse transcription–quantitative polymerase chain reaction
Normal human mammary epithelial cell line HBL-100 and breast cancer cell lines MCF-7, MDA-MB-231, and SKBR-3 were collected. The total RNA of the aforementioned cells was extracted using Trizol reagent (A33254; Thermo Fisher Scientific, Waltham), of which the concentration and purity were determined in connection with an ultra micro spectrophotometer (NanoDrop2000; Thermo Fisher Scientific). Based on the first-strand synthesis reverse transcription kit (#k1622; Thermo Fisher Scientific), a total of 10 μL cDNA was obtained in connection with the reverse transcription of the sample RNA. Next, 65 μL of diethyl pyrocarbonate was added to the obtained cDNA for dilution and fully mixed.
The reverse transcription–quantitative polymerase chain reaction (RT-qPCR) system and PCR reaction procedure were set as follows: 5 μL SsoFast EvaGreen Supermix (2 × ), 0.5 μL forward primer (10 μM), 0.5 μL reverse primer (10 μM), and 4 μL cDNA. The PCR amplification conditions were set as follows: predenaturation for 1 min at 95°C, denaturation for 30 s at 95°C, extension for 60 s at 72°C, and annealing for 5 s at 58°C, repeated 30 cycles that were followed by final extension for 5 s at 65°C ∼95°C. The RT-qPCR primers were designed and synthesized by Huada Gene Scientific and Technological Co., Ltd. (Shenzhen, China) (Table 1). The expression of glyceraldehyde-3-phosphate dehydrogenase was employed as the reference for the calculation of the relative expression of HOTAIR based on the relative quantitative formula 2−ΔΔCt, followed by a statistical analysis of the obtained result.
EZH2, enhancer of zeste homologue 2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HOTAIR, HOX transcript antisense intergenic RNA.
MTT assay
Breast cancer MCF-7, MDA-MB-231, and SKBR-3 cells in the logarithmic growth phase were collected and then resuspended in cell growth medium. The cell density was adjusted to 1.0 × 104 cells/mL followed by inoculation of the cells into a 96-well plate (100 μL each well). Next, following 24 h of cell culture, once cell adherence was observed, the cell culture medium was discarded.
Cells in each group were transfected with three preset holes for each group. After 6 h of culture, the cells underwent a subsequent culture process for 24, 48, and 72 h, respectively, in normal medium. The cultured solution in each hole was obtained and washed thrice with sterile phosphate buffered saline (PBS), followed by the removal of the remaining FBS and phenol red to avoid FBS interference with phenol red in the complete medium. Next, 10 μL MTT solution (M2128-1G; Sigma-Aldrich, Ronkonkoma, NY) was added to each hole and incubated for 4 h under conditions void of light. The culture medium in the 96-well plate was carefully drawn, with each 100 μL dimethyl sulfoxide solution added to each hole (D2650-100ML; Sigma-Aldrich) under conditions void of light.
The cells in flat plates were dissolved over a period of 15 min by means of vibration using an oscillator without lighting, followed by detection using a microplate reader. The optical density (OD) value of each hole was detected at a wavelength of 570 nm using a full wavelength microplate reader (51119200; Thermo Scientific™), with the breast cancer cell survival rates among the various groups subsequently calculated in accordance with the following formula: cell survival rate (%) = OD value of experimental group/OD value of normal group × 100%. Meanwhile, the cells were photographed under an inverted microscope allowing for closer observations of the changes to the cell morphology at the start of culture and after the silence of HOTAIR.
Scratch test
Each well of the six-well plate was seeded with 5 × 105 cells with the different siRNA added to each group accordingly with three holes preset for each group. After a 6-h period of culturing had elapsed, the medium was replaced by a normal culture medium, and a straight line was drawn on the cell surface under the guidance of the straight edge of a sterile gun. After three PBS rinses, the cells were photographed under an inverted microscope (200 × ) with the scratch distance measured at both 0 and 48 h after culture.
Transwell assay
Matrigel (356234; Corning, Corning City, NY) was diluted with precooled serum-free DMEM at a ratio of 1:10, and after fully mixing, 100 μL diluted Matrigel was added to each of the upper chambers, maintained at room temperature for 2 h, and rinsed with 200 μL serum-free DMEM medium before use.
After digestion and resuspension using serum-free DMEM, the cells was calculated and diluted to 3 × 105 cells/mL. The upper Transwell chamber was dispensed with 100 μL diluted cells; meanwhile, 600 μL DMEM containing 10% serum (as a chemokine) was added to the Transwell lower chamber. According to the Transwell introduction, the Transwell chambers were collected after 24 h of incubation, followed by 5 min of methanol fixation and two PBS washes. The Transwell chambers were photographed and observed under a microscope after eosin staining, with the total number of transmembrane cells tallied.
Flow cytometry
The breast cancer cells were transfected, treated, and then centrifuged at 800 rpm with a centrifugal radius of 15 cm for 5 min. Following two PBS washes, based on the instructions of the Annexin-V-FITC Cell Apoptosis Detection Kit (K201-100; Biovision, San Francisco), the cells were resuspended in 200 μL binding buffer followed by the addition of 10 μL of PI dye liquid and 5 μL of Annexin V-FITC. After incubation under dark conditions at room temperature for 15 min, flow cytometry was applied to determine the DNA concentration of the cells (Becton, Dickinson and Company, NJ) at a wavelength of 488 nm. The experiment was repeated thrice.
Dual-luciferase reporter gene assay
MCF-7 cells in breast cancer were collected and transfected with plasmid DNA (PTEN-wild type [WT] or PTEN-mutant type [MUT]) and HOTAIR (50 nM) or NC. After 24 h of transfection had elapsed, a multimode reader was used to carry out the experiment based on the instructions of Dual-Luciferase Assay Report Gene Experiment Kit (BioTek, Winooski, VT). Firely luciferase and Renilla luciferase were regarded as the relative luciferase activities. The experiment was repeated thrice.
Statistical analysis
SPSS 21.0 software (IBM Corporation, New York, NY) was used for statistical data analysis. Each experiment was performed in triplicate or more. Measurement data were presented as mean ± standard deviation. Comparisons among multiple groups were assessed by means of one-way analysis of variance. Student-Newman-Keuls test was conducted for pairwise comparison. p < 0.05 was considered to be statistically significant.
Results
HOTAIR and EZH2 are both highly expressed in breast cancer cells
Compared with normal human mammary epithelial cell line HBL-100, breast cancer cell lines MCF-7, MDA-MB-231, and SKBR-3 exhibited notable increases of HOTAIR and EZH2, which were determined to be 25.6 and 6.3 times, 11.2 and 4.3 times, and 15.8 and 3.8 times, respectively, compared to the expression of HOTAIR and EZH2 of the HBL-100 cells. The highest expression of HOTAIR and EZH2 in MCF-7 cells was recorded in the three breast cancer cells, and as a result of this, the subsequent experiments were performed using the breast cancer cell line MCF-7. The expressions of HOTAIR were significantly downregulated in the MCF-7 cells after HOTAIR and EZH2 interference, with a distinct effect observed in regard to the interference of siRNA interference in three breast cancer cells.
Compared with the control group, no significant difference was observed regarding the expressions of HOTAIR and EZH2 in the NC group, while comparisons with the control and NC groups revealed there to be reduced HOTAIR mRNA in the siRNA-HOTAIR and siRNA-HOTAIR+siRNA-EZH2 groups (both p < 0.05). Elevated expressions of HOTAIR were detected in the siRNA-EZH2 group when compared with the siRNA-HOTAIR group (p < 0.05). Furthermore, no notable differences were observed in the siRNA-HOTAIR+siRNA-EZH2 group when compared with the siRNA-HOTAIR group (p > 0.05).
In addition, analysis of EZH2 indicated that when compared with the control group and NC group, EZH2 mRNA expressions in the siRNA-EZH2 and siRNA-HOTAIR+siRNA-EZH2 groups were significantly decreased (both p < 0.05). Compared with the siRNA-EZH2 group, the EZH2 mRNA expressions were significantly increased in the siRNA-HOTAIR group (p < 0.05), while no distinct difference was observed among the siRNA-HOTAIR+siRNA-EZH2 groups (p > 0.05) (Fig. 1). The aforementioned results suggested that the expression of HOTAIR in MCF-7 cells were the highest, with elevated levels of HOTAIR expression recorded in the siRNA-EZH2 group.
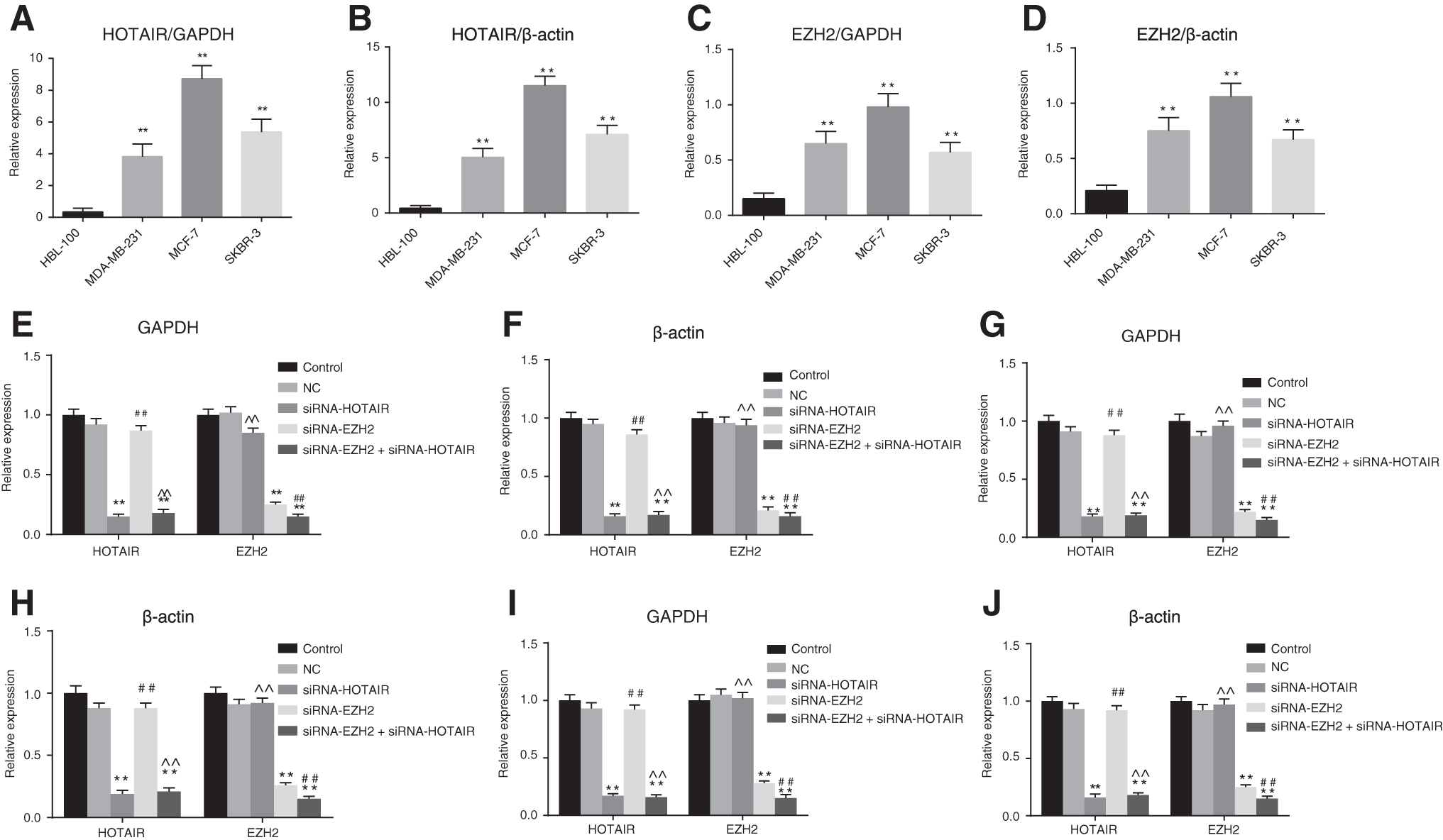
The results illustrated that HOTAIR and EZH2 were both overexpressed in breast cancer cells.
Downregulation of HOTAIR or EZH2 leads to abnormal morphology of the breast cancer cells
The photographs obtained under an inverted microscope revealed that, the cell morphology and density of the three cells had changed significantly before and after HOTAIR and EZH2 interference. The cells were observed to be regular in cellular morphology, as well as clear and bright in both the control and NC groups. In the siRNA-HOTAIR group, the cells were observed to be irregular in cell morphology with membrane shrinkage and decreased density and proliferation, with a greater number of dead cells observed compared with that of the control group.
Similar results were observed in the siRNA-EZH2 group, with that of the siRNA-HOTAIR group. In the siRNA-HOTAIR+siRNA-EZH2 group, compared with cells in groups of siRNA-HOTAIR and siRNA-EZH2, the cells exhibited irregular cell morphology with signs of membrane shrinkage, with a more significant degree of decreased density and proliferation (Fig. 2).

The results of cell morphological observation showed that downregulation of HOTAIR and EZH2 presented abnormal morphology of breast cancer cells. EZH2, enhancer of zeste homologue 2; HOTAIR, HOX transcript antisense intergenic RNA.
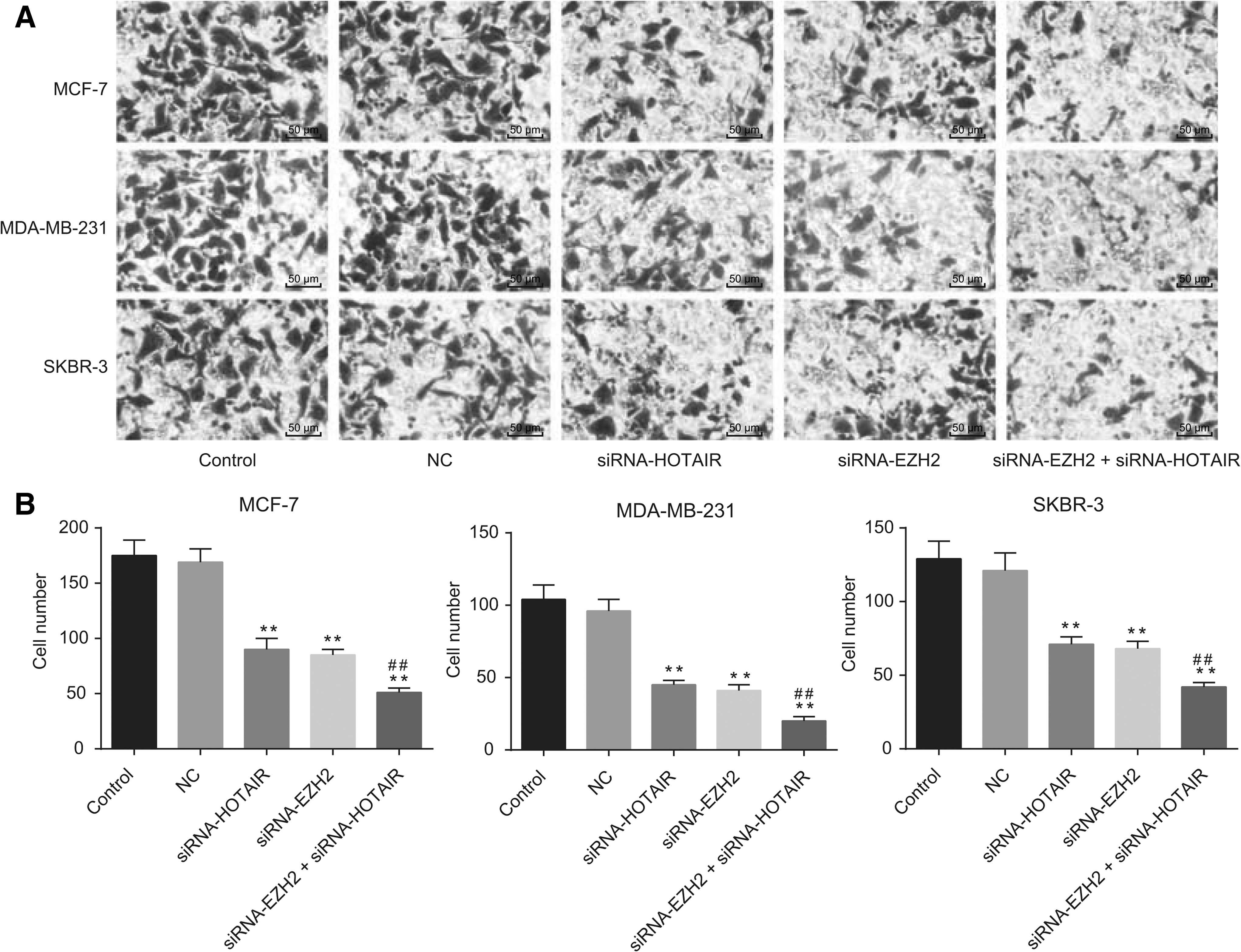
The combined downregulation of HOTAIR and EZH2 exhibits the most effective inhibition of breast cancer cell proliferation
The employed MTT assay revealed that the interference of HOTAIR and siRNA-EZH2 inhibited the proliferation of breast cancer in the MCF-7 cells in a time-dependent manner, whereby the inhibitory effect was enhanced with increase in time. The inhibitory effect on cell proliferation peaked at 72 h, while no significant difference between the control group and the NC group was observed (p > 0.05). Compared with the control and NC groups, the siRNA-HOTAIR group displayed distinct cell proliferation, while similar results in the siRNA-EZH2 and siRNA-HOTAIR groups were observed (all p < 0.01). Comparisons between the siRNA-HOTAIR and siRNA-EZH2 groups revealed a more distinct degree of cell proliferation inhibition in the siRNA-HOTAIR+siRNA-EZH2 group (all p < 0.01) (Fig. 3).

The results of MTT assay indicated that the downregulation of HOTAIR and EZH2 inhibited proliferation of breast cancer cells.
The combined downregulation of HOTAIR and EZH2 exhibits the most effective inhibition of breast cancer cell invasion and migration
Scratch test results revealed that the migration of the three breast cancer cells after HOTAIR and EZH2 interference was significantly decreased when compared to the control and NC groups (p < 0.01); no significant difference was observed in the NC group when compared to the control group (p > 0.05). Compared with the siRNA-HOTAIR group, no significant difference was detected in the siRNA-EZH2 group, while comparisons to the control and NC groups revealed there to be a reduced rate of cell migration in the siRNA-EZH2 and siRNA-HOTAIR+siRNA-EZH2 groups (all p < 0.01).
Transwell assay results indicated that after silencing HOTAIR, invasion of cells through the Matrigel and the chamber membrane in the siRNA-HOTAIR group was markedly decreased, compared with the control and NC groups (p < 0.01). The invasive ability was notably reduced in the siRNA-EZH2 group (p < 0.01), while no significant difference was detected when comparing the siRNA-HOTAIR and siRNA-EZH2 groups (p > 0.01). Decreased invasive ability was noted in the siRNA-HOTAIR+siRNA-EZH2 group when compared with the siRNA-HOTAIR group (p < 0.01). However, no significant difference was detected between the control and NC groups (p > 0.05) (Figs. 4 and 5).

Scratch test results revealing that downregulation of HOTAIR and EZH2 inhibited migration of breast cancer cells.
Transwell assay results suggesting that downregulation of HOTAIR and EZH2 inhibited invasion of breast cancer cells.
The combined downregulation of HOTAIR and EZH2 together has the most significant enhancement of breast cancer cell apoptosis
Flow cytometry results demonstrated that after siRNA-interfered HOTAIR and EZH2, the apoptosis among the three breast cancer cell lines was promoted, while no obvious difference between the control group and the NC group was noted (p > 0.05). In comparison with the control and NC group, cell apoptosis in the siRNA-HOTAIR group was significantly elevated, which was also observed in the siRNA-EZH2 group (p < 0.01). Compared with the siRNA-EZH2 and siRNA-HOTAIR group, the siRNA-HOTAIR+siRNA-EZH2 group displayed a more distinct level of improved cell apoptosis (both p < 0.01) (Fig. 6).
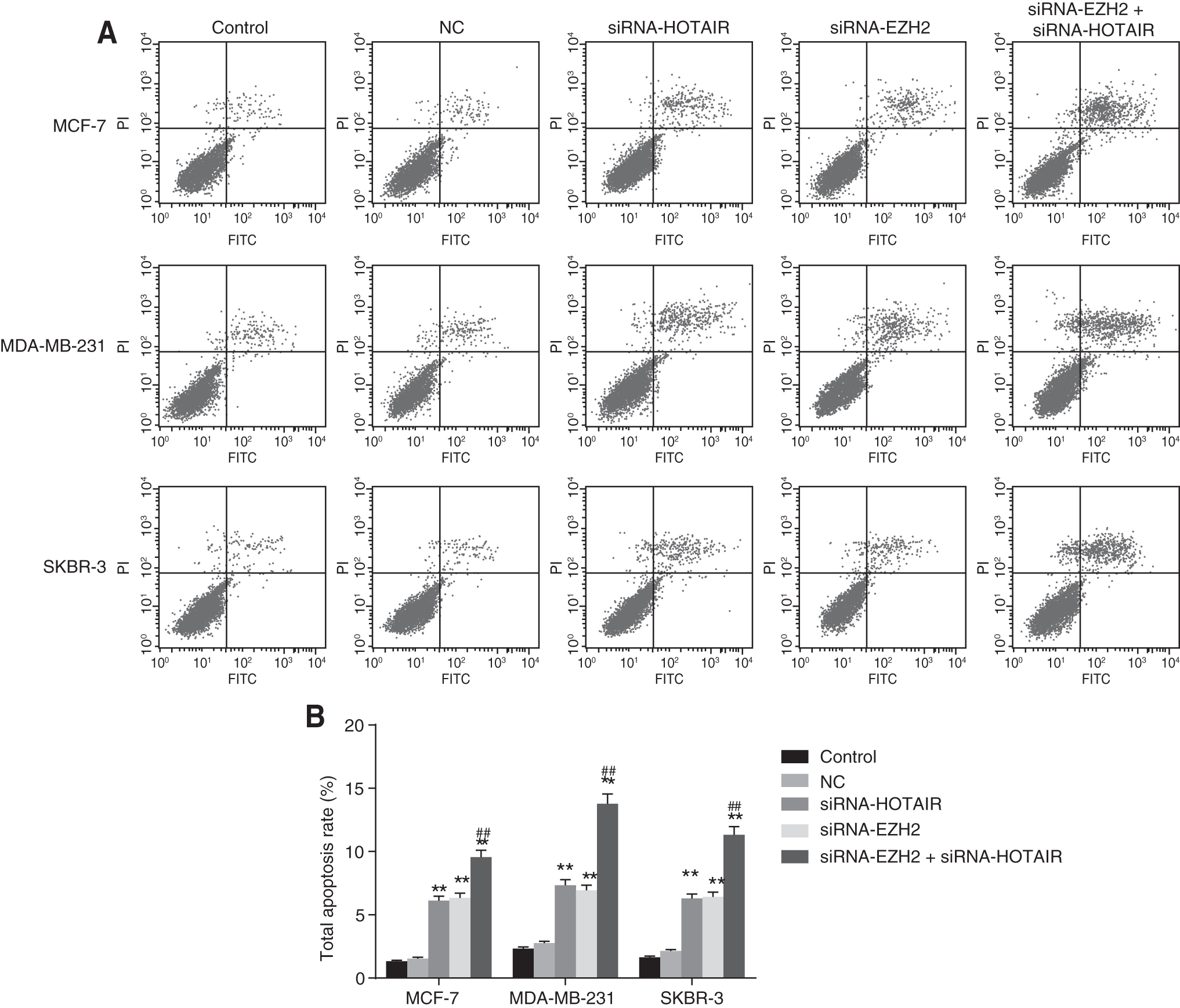
The results of flow cytometry showed that apoptosis of breast cancer cells MCF-7, MDA-MB-231, and SKBR-3 was promoted by siRNA-interfered HOTAIR and EZH2.
HOTAIR inhibits PTEN expression by recruiting EZH2 in breast cancer
The Cancer Genome Atlas (TCGA) database indicated that the PTEN expression was low in breast cancer. The binding site of HOTAIR and PTEN was predicted in connection with the bioinformation website (
Meanwhile, the bioinformation predication results demonstrated that HOTAIR could bind specifically to EZH2 (Fig. 7). All the aforementioned results presented indicated that HOTAIR could act to inhibit PTEN expression by recruiting EZH2 in breast cancer.
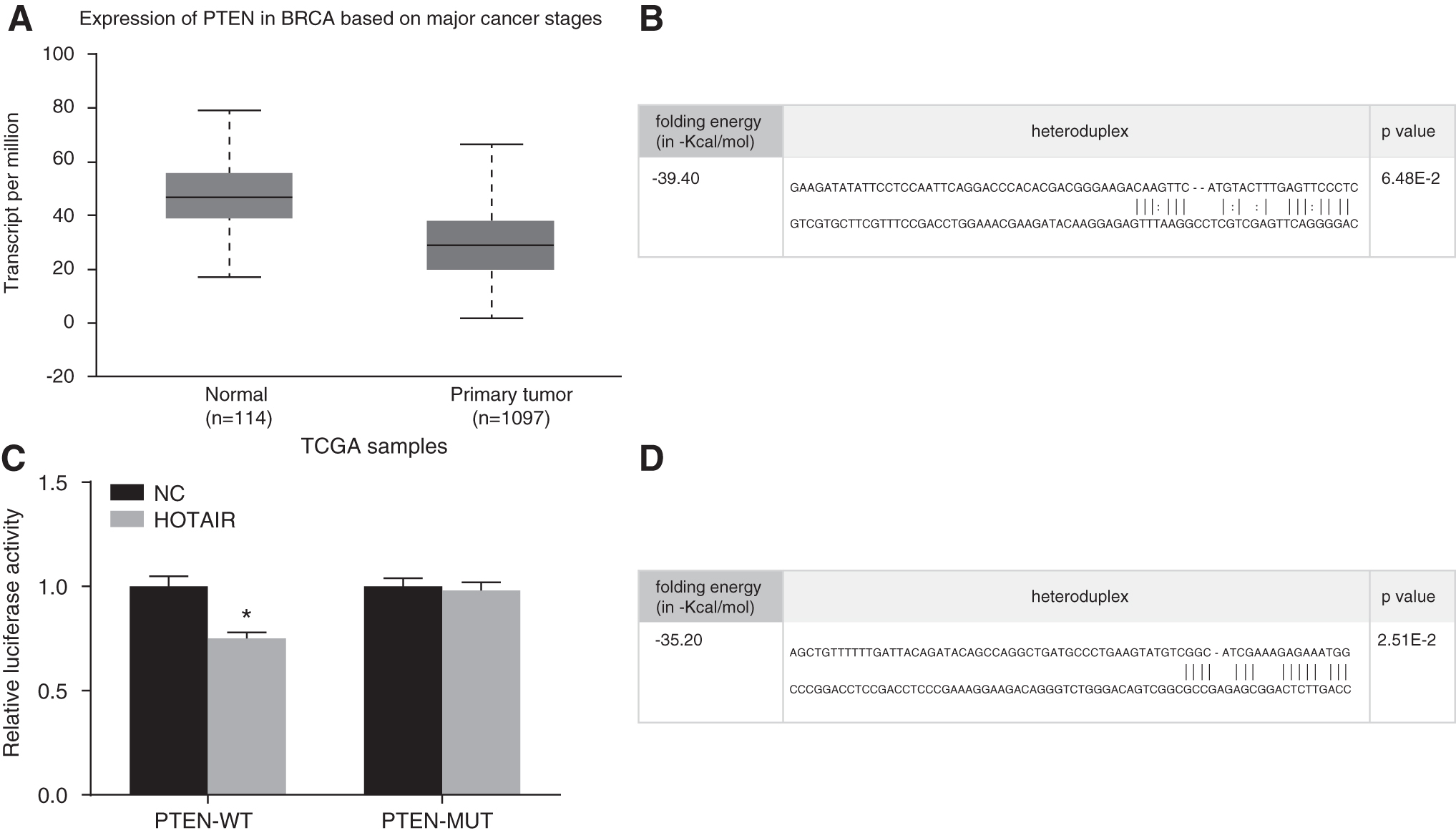
Bioinformation predication and dual-luciferase gene reporter results demonstrating the inhibition of PTEN expression by HOTAIR's recruiting of EZH2.
Discussion
Representing the most common cancer faced by the female population, with the incidence increasing with age, breast cancer has been reported to be rising in morbidity in various parts of the world. 18,19 The etiology of breast cancer is one of clinical heterogeneity, with distinct molecular aberrations, requiring a series of treatments that are accompanied by results with varying outcomes. 20,21 Breast cancer deaths are generally brought on by the hematogenous spread of pathological cells from the primary site before migration to distant tissues and subsequent metastasis. 22 Thus, there is a strong clinical requirement for more accurate prognoses of metastasis, which may save a large number of patients with breast cancer from unnecessary adjuvant systemic therapy and other related costly medical treatments. 23
During this study, the authors aimed to evaluate the role by which lncRNA HOTAIR and EZH2 act to influence proliferation, invasion, migration, and apoptosis of breast cancer cells. Based on the results of a series of experiments, evidence was provided, demonstrating there to be elevated expression of HOTAIR and EZH2 in breast cancer cells, while indications were presented suggesting that downregulation of HOTAIR and EZH2 could act to suppress the invasion and migration, while acting to promote the apoptosis of breast cancer cells, highlighting a potential biomarker for the treatment of breast cancer.
Initially, RT-qPCR was applied to determine the expressions of HOTAIR and EZH2 in normal human mammary epithelial cell line HBL-100 and breast cancer cell lines MCF-7, MDA-MB-231, and SKBR-3, the results of which revealed there to be higher expression of HOTAIR and EZH2 in breast cancer cells, particularly in the MCF-7 cells. Expression of HOTAIR has been evaluated in various types of human cancer as well as in other diseases, with its overexpression implicated in the poor survival observed among patients with thyroid carcinoma, solid tumors, and acute leukemia. 24 Previous studies have revealed there to be distinct upregulation of HOTAIR in nonsmall-cell lung cancer (NSCLC) tissues, which act to promote NSCLC cell metastasis and invasion. 25
Evidence has been presented suggesting that HOTAIR expression was upregulated in pancreatic tumors, which has since been correlated with the incidence of more aggressive tumors when compared to nontumor tissues. 26 Further study into HOTAIR and EZH2 in breast cancer cells, which were silenced by RNA interference, is required. During this study, the authors detected that compared with the blank and NC groups, there were more apoptotic cells and decreased proliferation of breast cancer cells in the siRNA-HOTAIR group, providing evidence that HOTAIR silencing could inhibit the proliferation of breast cancer cells.
Consistent with the authors' results, a previous study demonstrated that HOTAIR knockdown in pancreatic cancer cells could induce apoptosis, alter cell cycle progression, and decrease cell proliferation, demonstrating the expanded function of HOTAIR. 26,27 The knockdown of HOTAIR, in cervical cancer cells, was previously found to inhibit proliferation, while promoting apoptosis. 28 HOTAIR is involved in epigenetic regulation owing to its interaction with Polycomb Repressive Complex 2, which induces trimethylation of histone H3 on lysine27 (H3K27me3), resulting in a reduction in the expression of multiple genes. 29
Silencing of HOTAIR has also been determined to decrease the binding and recruitment abilities of H3K27me3 and EZH2 gene locus with HOTAIR as well as cell cycle-related genes remaining active for transcription through the modulation of covalent histones and silencing of HOTAIR expression. 30 Liu et al. indicated that the knockdown of EZH2 could significantly inhibit proliferation, while improving the rate of apoptosis of hepatocellular carcinoma cells. 31 Investigations into the knockdown of HOTAIR revealed there to be a weakening in the recruitment and binding abilities of EZH2, 30 which interestingly was observed in this study, adding further validation to the notion that HOTAIR can recruit EZH2.
Most importantly, the authors estimated the effects of HOTAIR and EZH2 silencing on breast cancer cell migration through the application of a Scratch test and invasion by means of a Transwell assay. The authors' findings illustrated that the invasion and migration of the three breast cancer cells were markedly inhibited after HOTAIR and EZH2 expressions were inhibited. By recruiting the gene silencing machinery to the promoters of target gene, HOTAIR can indirectly reprogram chromatin state leading to gene silencing. 12,32 Furthermore, previous studies provided support to the notion that increased expressions of lncRNA HOTAIR can promote the migration, proliferation, and invasion of ovarian cancer cells by modifying chromatins. 33,34
Notably, previous studies have suggested that the knockdown of lncRNA HOTAIR could reverse epithelial–mesenchymal transition and tumor invasion in cases of colorectal cancer. 35 Verification has been previously presented, indicating that the knockdown of HOTAIR results in reduced expression of the multidrug-resistant protein 1 in K562-imatinib cells, leading to increased sensitivity to imatinib treatment, as well as markedly attenuating the activation of PI3K/Akt. 36,37
Reports have indicated that as a kind of HMT, EZH2 exhibits abundant levels in numerous types of cancer, including breast cancer. 38 EZH2 inhibits gene transcription and mediates epigenetic downregulation of genes that are related with cell proliferation, migration, and invasion. 39 Thus, it was subsequently concluded that the downregulation of HOTAIR and EZH2 results in inhibition of the processes of proliferation, invasion, and migration, while acting to promote the apoptosis of breast cancer cells. Taken together, the key findings of this study has presented significant evidence that the knockdown of HOTAIR can inhibit the proliferation, invasion, and migration, while inducing the apoptosis of breast cancer cells by recruiting EZH2.
These results highlight the promise of HOTAIR as a novel predictor of poor prognosis for patients suffering from breast cancer, thus indicating that the reduction of lncRNA HOTAIR and EZH2 can inhibit the development of breast cancer, which may be of significant clinical use in identifying more effective therapeutic strategies for patients with breast cancer. Although the occurrence mechanism of the binding of HOTAIR and PTEN was not investigated during this study, due to time and financial restraints, the authors aim to further explore the mechanisms behind the findings of this study, in addition with larger sample sizes required for future studies to further confirm the findings in relation to the effects associated with HOTAIR.
Footnotes
Acknowledgment
The authors would like to acknowledge the reviewers for their helpful comments on this article.
Disclosure Statement
No competing financial interests exist.