
Abstract
Mutant-p53 colorectal cancer (CRC) cells are often resistant to radiotherapy. Loss of suppressor activity of wt-p53 on survivin is responsible for the enhanced expression of survivin as a radioresistant factor in tumor cells. Yet, no survivin-modulating drug has been approved for clinical application in CRC. Thus, the search for safe compounds that modulate survivin expression and induce apoptosis irrespective of p53 status may potentiate the efficacy of radiotherapy in mutant-p53 CRC cells. Omega-3 docosahexaenoic acid (DHA) induces apoptosis in malignant cells without cytotoxicity in normal cells. However, little is known whether in vitro concentrations of DHA equal to the human plasma levels are able to modulate expression of survivin and sensitize mutant-p53 CRC cells to γ-irradiation. Radioresistant mutant-p53 HT-29 cells were pretreated with 50- and 100-μM DHA for 48-h before 2-, 4-, 6-, 8-, and 10-Gy of γ-irradiation. Thereafter, proliferation rates were measured after 6 d. HT-29 cells were also pretreated with 50- and 100-μM DHA for 4-h before 2- and 10-Gy of γ-irradiation after which, cell number, survivin expression, caspase-3 activation, apoptosis, and ED50 (γ-irradiation dose causing 50% growth inhibition) were evaluated. Pretreatment of HT-29 cells with 50- and 100-μM DHA for 48-h followed by 2- to 10-Gy of γ-irradiation induced a dose-dependent additive decrease in cell proliferation rate and ED50 values were decreased by 88%, 44%, 41%, and 27% for 500-, 1500-, 2500-, and 5000 cells per well pretreated with 100-μM DHA respectively. Pretreatment of 5 × 105 HT-29 cells per well with 100-μM DHA for 4-h followed by 2- or 10-Gy of irradiation resulted in 53% and 86% decreases in cell numbers, 2- and 5.1-fold activation in caspase-3 followed by 66% and 60% decreases in survivin mRNA levels respectively. DHA in combination with radiation increased total apoptotic rate 48-h post-treatment. DHA decreases survivin expression and induces caspase-3 activation irrespective of p53 status. Significant decreases in ED50 values at concentrations of DHA equal to human plasma levels, suggesting that DHA could be used as an attractive radiosensitizer agent in CRC patients with mutant-p53.
Introduction
Colorectal cancer (CRC) is one of the most common malignancies in the world. 1 Treatment of CRC usually involves surgical removal of the tumor followed by radio- and/or chemotherapy. Radio- and chemotherapy exert their therapeutic effects by targeting malignant cells. However, due to developing chemo- and radioresistant CRC cells, the rate of disease relapse in patients is extremely high. 2 Different factors including mutations of p53 3 and overexpression of survivin have been shown to involve in the radioresistance of CRC cells. 4 –6
P53 mutations happen in nearly 50% of human malignancies, which dramatically increase cellular resistance to γ-radiation. 7 Mutation of p53 induces high level of survivin expression, which plays an important role in resistance to apoptosis induction by chemo- and radiotherapy. 5 As a consequence, one possibility to obtain strong antitumor effects and radiation responses could be using safe natural compounds that exert proapoptotic effects and modulate expression of survivin in mutant-p53 CRC cells. This attractive strategy may diminish necessary radiation dose to sensitize cancer cells to apoptosis and may provide good toleration to treatment in patients with CRC. In this regard, omega-3 polyunsaturated fatty acids (PUFAs) including docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) are harmless, accessible, cheap, extensively used, and may serve as attractive compounds for evaluating their effects on radioresistant tumor cells.
It has been shown that, a number of molecular pathways have been identified for the activity of DHA and EPA against malignant cells 8,9 and these compounds enhance the effect of chemo- and radiotherapy on tumor cells 10 –12 without cytotoxicity on normal cells. 13 –15 In this context, it has been shown that, DHA induces apoptosis irrespective of p53 status 16 and dramatically decreases expression of survivin more efficiently than EPA in CRC cells. 17
In different studies, it has been shown that, PUFAs increase chemo- and radiosensitivity in different kinds of cancer cells such as colorectal adenocarcinoma cells and breast cancer cells. 10 –12 However, little attention has been paid to the evaluation of sensitization effect of DHA on radioresistant p53-mutant CRC cells with high level of survivin (radioresistant factor). With this in mind, we aimed to determine whether in vitro concentrations of DHA (50- and 100-μM) equal to the levels of DHA achievable in the human body following supplementation of the diet with different amount of omega-3 PUFAs/d 18 are able to modulate expression of survivin and sensitize radioresistant homozygous mutant-p53 HT-29 CRC cells 19,20 to γ-irradiation.
Materials and Methods
Cells, culture conditions, and oligonucleotides
HT-29 CRC cells with homozygous for p53 mutation (Arg-273→His)
20
was obtained from the Pasture Institute of Iran. These cells were cultured in RPMI-1640 medium containing 10% fetal bovine serum (FBS), penicillin and streptomycin all from PAA Laboratories (Austria), and 20 mM HEPES and 2 mM
Drug and kits
Pure (99%) fish oil-derived DHA (Fig. 1) was obtained from Sigma-Aldrich Chemical Company (Germany). The trypsin-EDTA mixture, caspase-3 activity and apoptosis assay kits were obtained from Roche. The RNA isolation kit was purchased from Sinaclon Bioscience Company (Tehran, Iran). The cDNA synthesis kit was purchased from Fermentas, Inc. (Burlington, ON, Canada). MTT powder was purchased from Atocel (Austria).
Structure of fish-oil derived docosahexaenoic acid.
Preparation of DHA stock solution
DHA was dissolved in absolute EtOH (ethanol) to provide a 100-mM stock solution. For each experiment, different concentrations of DHA were freshly prepared from the stock solution by serial dilutions in culture medium. Control cells (Vehicle) were treated with ethanol never exceeded 0.5% (v/v) in the experiments.
Ionizing radiation treatments
Theratron Phoenix Cobalt-60 system was used to irradiate cells in cell culture plates. The delivered doses were 2-, 4-, 6-, 8-, and 10-Gy (12 cm length × 9.5 cm width) at room temperature. The source to sample distance was 80 cm and the dose rate was 46.6 Gy/min. Irradiation time correction factors due to the decay of the Cobalt-60 source have been applied. After irradiation, the samples and vehicles were incubated at 37°C in a humidified CO2 incubator. The cells were cultured under the normoxic condition.
Cell count assays
About 5 × 10 5 HT-29 cells per well were plated in six-well plates in 2 mL complete medium and incubated at 37°C for 24 h. Thereafter, the cells were treated with DHA at concentrations of 50- and 100-μM alone for 48 h or pretreated with 50- and 100-μM DHA for 4 h before 2- or 10-Gy of γ-irradiation. Subsequently, after 48 h treatments, total cell numbers were counted under the microscope (Nikon, Tokyo, Japan).
Growth inhibition assays
Nearly, 5 × 10 3 HT-29 cells per well were plated in 96-well plates in RPMI-1640 medium containing 10% FBS for 24 h. After 24 h incubation, the cells were treated with various concentrations of DHA for 48-, 72-, and 96-h. Following each incubation period, MTT solution (5 mg/mL in phosphate-buffered saline [PBS]) was added to each well and the incubation was continued for 4 h at 37°C. Thereafter, the culture media were removed and replaced by 200 μL dimethyl sulfoxide (DMSO) to dissolve the MTT formazan crystals. The absorbance in each well was then measured with a microplate reader (Biotek, Winooski, VT) at 490 nm and a reference wavelength of 620 nm. The growth inhibition rates were calculated as 100% − (Asample − Ablank)/(Acontrol − Ablank) × 100% and the results were expressed as percentages of the control cells (Vehicle). To measure the total growth inhibition and the IC50 values of DHA, the percentages of proliferation rates on the y-axis were plotted against the concentrations of DHA on the x-axis. Finally, all calculations were performed using regression analysis.
Cell proliferation assays
HT-29 cells were plated in 96-well plates at different cell densities (500, 1500, 2500, and 5000 cells/well) in RPMI-1640 medium containing 10% FBS and kept at 37°C for 24 h. After 24 h incubation, the cells were pretreated with DHA at concentrations of 50- and 100-μM for 48 h before 2-, 4-, 6-, 8-, and 10-Gy of γ-irradiation. Thereafter, the cell cultured media were changed with fresh complete medium without DHA and the cells were kept for 6 d at 37°C in a humidified incubator with 5% CO2. Cell cultured media were daily renewed to maintain adequate nutrient concentrations. After 6 d, MTT solution (5 mg/mL in PBS) was added to each well and the incubation was continued for 4 h at 37°C. The culture media were removed and replaced by 200 μL DMSO to dissolve the MTT formazan crystals. The absorbance in each well was then measured with a microplate reader (Biotek) at 490 nm and a reference wavelength of 620 nm. Cell proliferation rates were calculated as (Asample − Ablank)/(Acontrol − Ablank) × 100% and the results were expressed as percentages of the untreated cells. To determine the doses causing 50% growth inhibition by γ-irradiations (ED50), the percentages of the proliferation rates on the y-axis were plotted against the doses of γ-irradiation on the x-axis. Thereafter, all calculations were performed using regression analysis.
RNA purification and reverse transcription PCR
About 5 × 10 5 HT-29 cells per well were plated in six-well plates in 2 mL RPMI-1640 medium containing 10% FBS and incubated at 37°C for 24 h. After 24 h incubation, the cells were treated with DHA at concentrations of 50- and 100-μM alone for 48 h or pretreated with DHA at concentrations of 50- and 100-μM for 4 h before 2- or 10-Gy of γ-irradiation. Subsequently, the cells were kept at 37°C in a humidified incubator with 5% CO2 for 48 h. Thereafter, total RNAs were purified from samples using the RNA preparation kit and used as templates to generate a first cDNA strand according to the manufacturer's protocols. The generated fragments were subsequently used as templates for PCR-amplification of double-stranded cDNA corresponding to a preselected region of the survivin and human glucose-6 phosphate dehydrogenase (GAPDH) coding sequences, using specific primers pairs (Table 1). The ratios of the survivin and GAPDH band densities were calculated using gel documentation (Gel Logic Pro 212 Imaging System; Kodak) and multiplied by 100 (survivin/GAPDH × 100). These ratios represent relative survivin mRNA levels.
List of Oligonucleotides Used in this Study
Annealing temperature in °C (number of cycles).
GAPDH, glucose-6 phosphate dehydrogenase; PCR, polymerase chain reaction.
Caspase-3 activity assays
To evaluate caspase-3 activation, HT-29 cells were treated as mentioned above. Thereafter, the cells were trypsinized and washed with ice-cold phosphate buffer and resuspended in ready to use cold lysis buffer for 25 min, followed by centrifugation at 12,000 g for 10 min at 4°C. Finally, the supernatants were used for caspase-3 activation assay. To this end, caspase-3 derived from cellular lysates was captured by a monoclonal antibody. Next, activated caspase-3 was cleaved proportionally through the addition of a fluorogenic substrate in a reaction buffer, after which free fluorescent 7-amino-4-trifluoromethylcoumarin was measured at an excitation of 380 nm and an emission of 460 nm.
Flow cytometric assays
To evaluate apoptosis and necrosis, HT-29 cells were treated as mentioned above. Next, the cells were trypsinized and washed with ice-cold phosphate buffer and resuspended in 100 μL 1 × binding buffer containing Annexin V-Fluorescein Isothiocyanate (FITC) at a density of 5 × 10 5 cells per milliliter. Thereafter, the cells were mixed and incubated in the dark at room temperature for 25 min. To distinguish cells with membrane damage, propidium iodide (PI) solution was added to the cell suspension before flow cytometric analysis (FACScan). Early apoptosis was defined as cells positive for Annexin V-FITC only. Late apoptosis was defined as cells positive for Annexin V-FITC and PI. Necrotic cells were defined as cells positive for PI only. Total apoptosis was defined as the sum of early and late apoptotic cells.
Dose reduction factor (DRF) was defined as the ratio of apoptosis induced by radiation alone to radiation-induced apoptosis in the presence of DHA. 21
Statistical assessment
The data represent the mean ± standard deviation from two independent experiments with duplicate or triplicate assays. All calculations were performed using SPSS 15 (Chicago, IL). Analysis of variance was used for multiple comparisons. Correlations were assessed by the Pearson correlation analysis. p < 0.05 was considered to be statistically significant.
Results
Effect of DHA and ionizing radiation on cell number
Following 48 h treatments, we found that, DHA (50- and 100-μM) diminished cell number by 78% and 63% in a dose-dependent manner (Fig. 2). Pretreatment of HT-29 cells with 50- and 100-μM DHA for 4 h followed by 2- and 10-Gy of irradiation, induced a dose-dependent additive decrease in cell number compared with either treatment with DHA or γ-irradiation alone 48 h postirradiation. Specifically, pretreatment of HT-29 cells with 100-μM DHA followed by 10 Gy of γ-irradiation resulted in 86% decrease in cell number compared with untreated cells (Fig. 2).
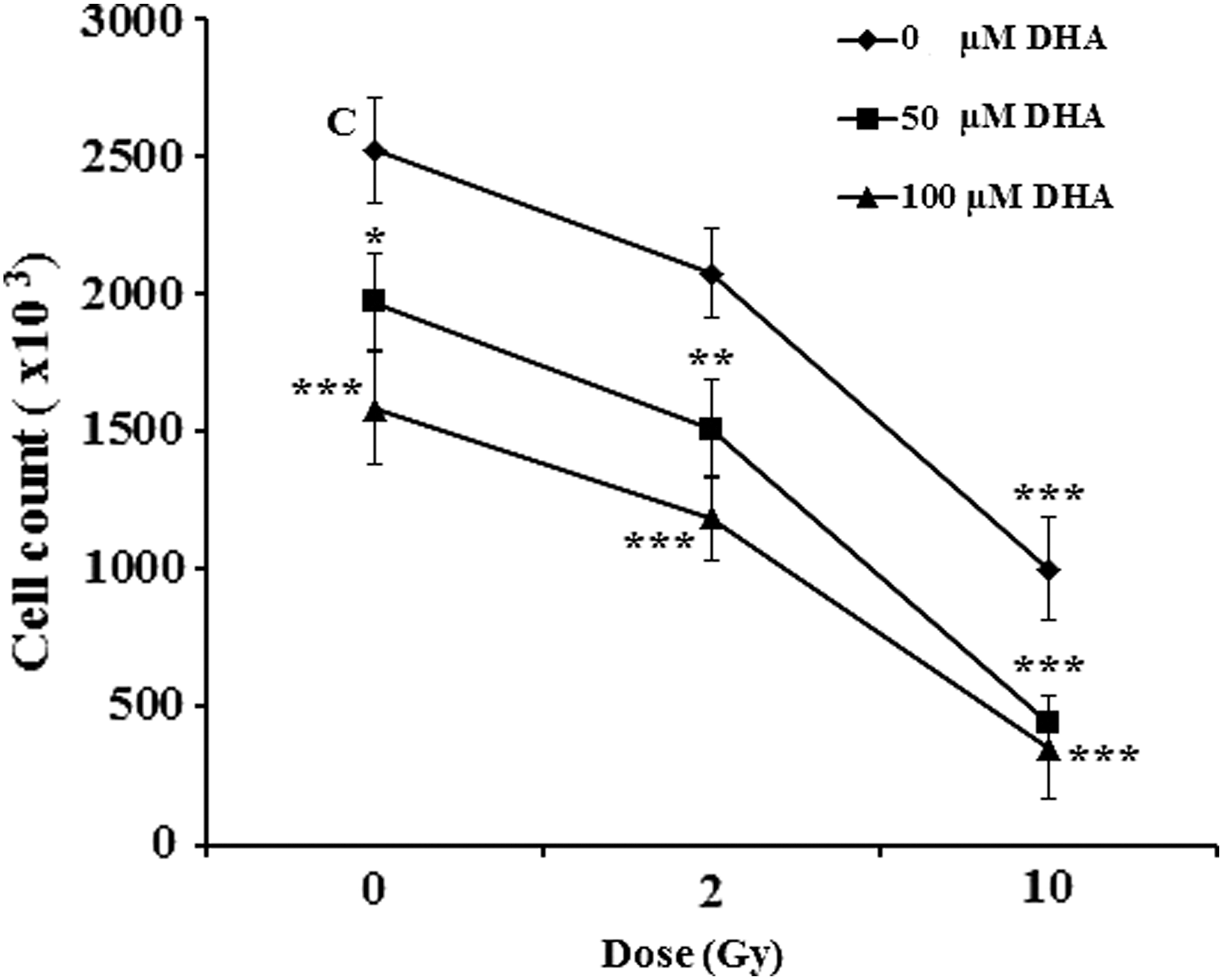
Effect of DHA alone or in combination with ionizing radiation on cell number. 5 × 10 5 HT-29 cells per well were treated either with DHA (50- and 100-μM) alone for 48 h or pretreated with 50- and 100-μM DHA for 4 h followed by 2- or 10-Gy of γ-irradiation. Next, the cell numbers were determined under the inverted microscope 48 h post-treatments. The results represent at least mean of two independent experiments ± SD with duplicate assays. *p < 0.05, **p < 0.01 and ***p < 0.001 versus untreated control cells (C; Vehicle). DHA, docosahexaenoic acid; SD, standard deviation.
DHA effect on growth inhibition
Increase in growth inhibition of HT-29 cells was observed with increasing doses of DHA and durations of exposure to DHA. Significant reduction in IC50 value of DHA was observed after 96 h treatment with DHA compared with other treatments and was measured to be 80.2 ± 11.5 μM (Table 2). We also found that DHA at concentrations of 50- and 100-μM showed no cytotoxicity on peripheral blood mononuclear cells isolated from healthy persons after 48 h treatment.
Evaluation of Growth Inhibition Rates in Docosahexaenoic Acid-Treated Cells
5 × 10 3 HT-29 cells per well were treated with various concentrations of DHA and growth inhibition rates, IC50 and total growth inhibition were measured after 48 to 96 h. Data represent at least means of two independent experiments ± standard deviation with triplicate assays.
p < 0.001 versus untreated control cells (Vehicle).
DHA, docosahexaenoic acid.
Effect of DHA and ionizing radiation on cell proliferation rates
Cell proliferation rates depended on the concentrations of DHA and the doses of γ-irradiation used. DHA in combination with ionizing radiation was more efficient than DHA alone. Pretreatment of HT-29 cells with 50- and 100-μM DHA for 48 h followed by 2- to 10-Gy of γ-irradiation, induced a dose-dependent additive decrease in cell proliferation rate after 6 d. Specifically, pretreatment with 100-μM DHA followed by 10 Gy of γ-irradiation resulted in 70.6% decrease in cell proliferation compared with vehicle (Fig. 3a–d).
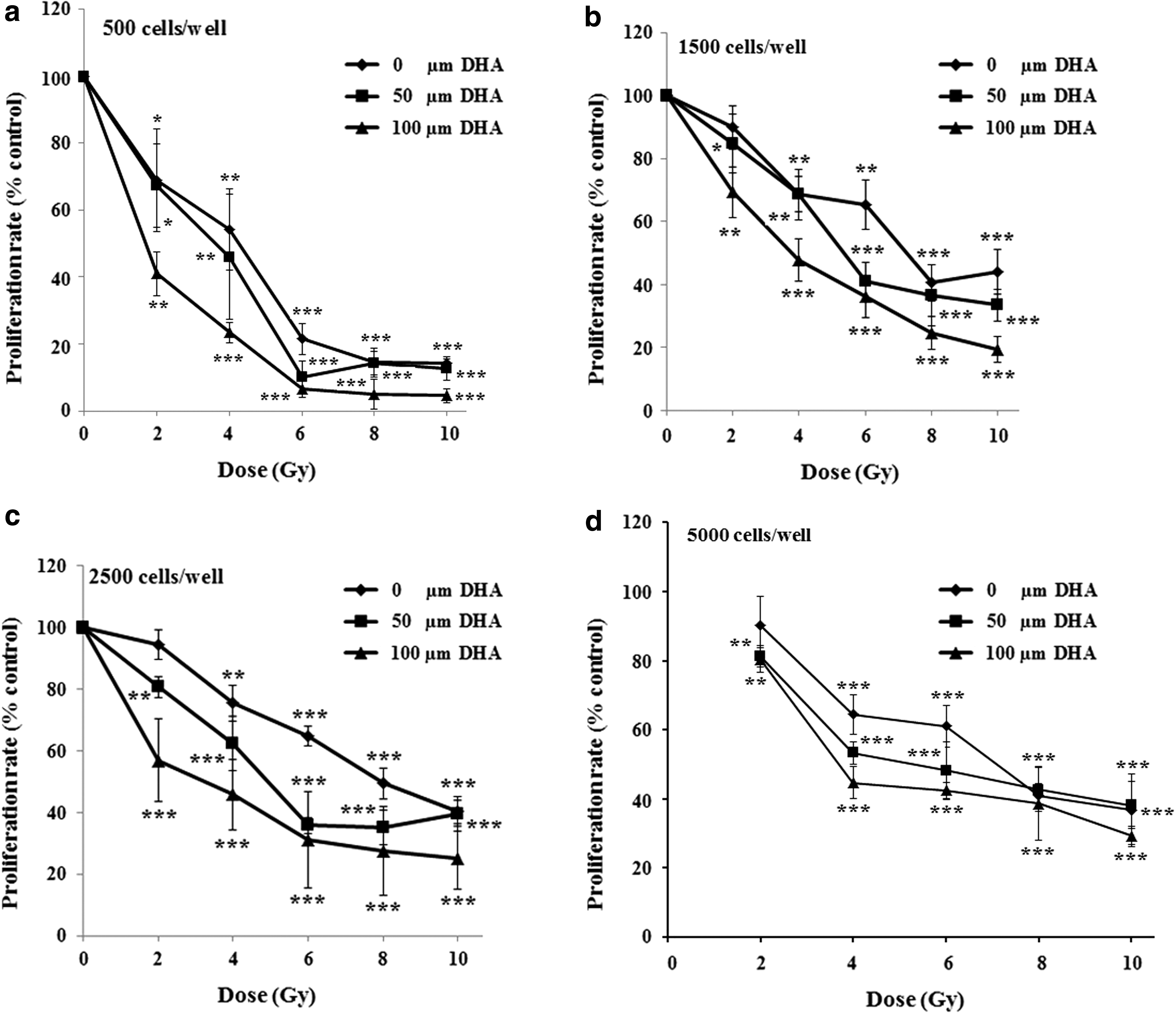
Effect of DHA alone or in combination with ionizing radiation on cell proliferation
Effect of DHA on ED50
Treatment with DHA diminished required doses of γ-irradiation causing 50% growth inhibition (ED50) (Table 3). In this regard, pretreatment with 50-μM DHA followed by 2- to 10-Gy of γ-irradiation induces 23%, 19%, 27%, and 16% decreases in ED50 for 500-, 1500-, 2500-, and 5000 cells per well, respectively. At the same conditions, pretreatments with 100-μM DHA, induces 88%, 44%, 41%, and 27% decreases in ED50 values for 500-, 1500-, 2500-, and 5000 cells per well, respectively.
Effect of Docosahexaenoic Acid Alone or in Combination with γ-Irradiation on ED50 and Lethal Dose
Different cell densities of HT-29 were pretreated with 50- and 100-μM DHA for 48 h followed by 2- to 10-Gy of γ-irradiation and ED50, and lethal dose of γ-irradiation were measured after seven doubling time of cells (6 d). To determine the doses causing 50% growth inhibition by γ-irradiation (ED50) and lethal dose, the percentages of the proliferation rates on the y-axis were plotted against the doses of γ-irradiation on the x-axis. Thereafter, all calculations were performed using regression analysis. Data represent at least means of two independent experiments ± standard deviation with triplicate assays.
DHA, docosahexaenoic acid.
Effect of DHA and ionizing radiation on the survivin mRNA levels
Following 48 h treatment with DHA alone or in combination with γ-irradiation, the relatively high level of survivin expression in untreated HT-29 cells diminished with increasing concentrations of DHA and γ-irradiation (Fig. 4a, b). We also found that pretreatment with 50-μM in combination with 2- or 10-Gy of γ-irradiation decreased survivin mRNA levels compared with DHA alone. Interestingly, treatment with 100-μM DHA alone resulted in highest decrease in survivin mRNA levels compared with other treatments. Treatment of cells with γ-irradiation alone had little effect on survivin mRNA levels compared with vehicle and the level of survivin mRNA was higher in cells treated with 10 Gy compared with 2 Gy of γ-irradiation.
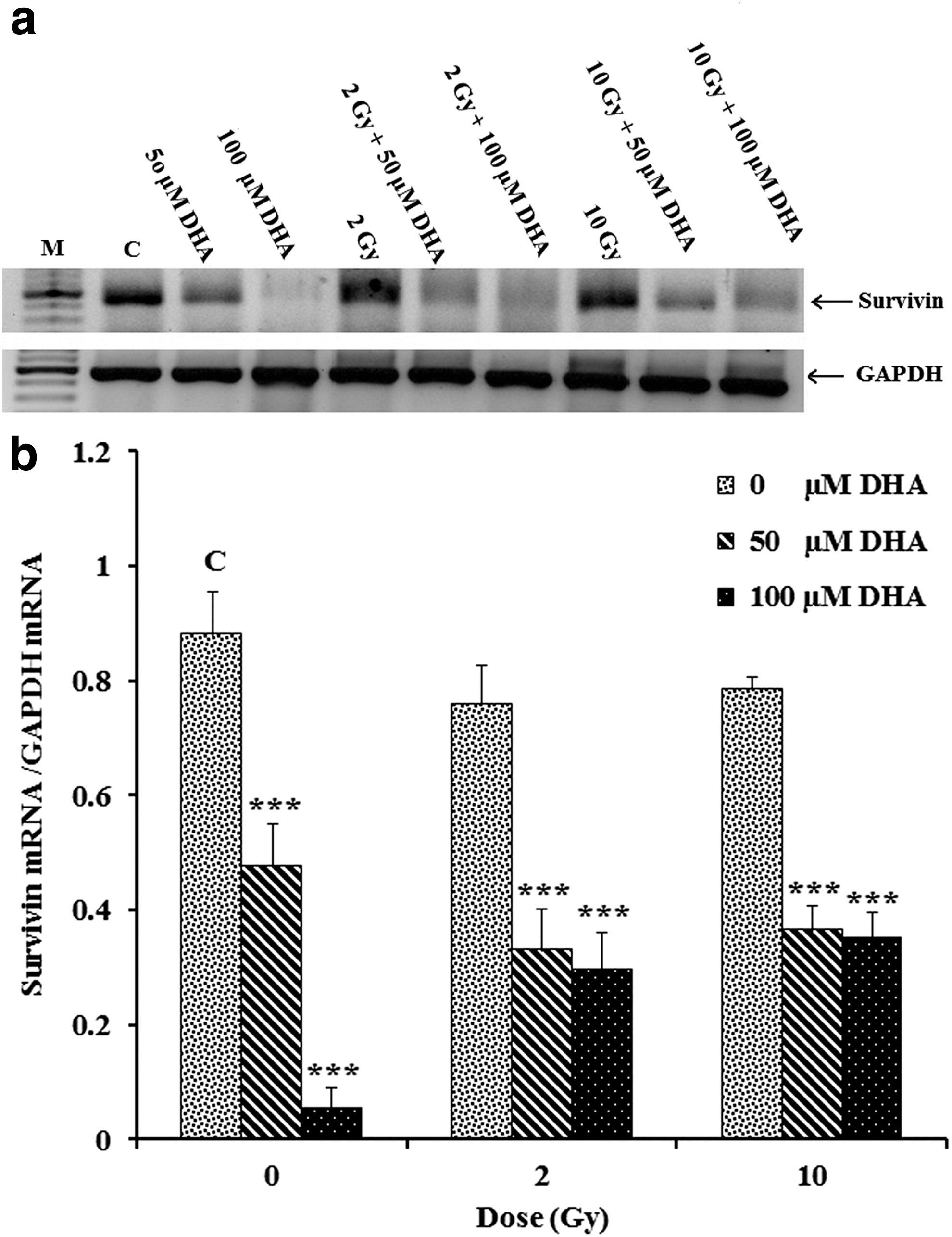
RT-PCR detection of survivin mRNA in HT-29 cells treated with DHA alone or in combination with ionizing radiation. About 5 × 10
5
HT-29 cells per well were treated either with DHA (50- and 100-μM) alone for 48 h or pretreated with 50- and 100-μM DHA for 4 h followed by 2-or 10-Gy of γ-irradiation, after which total RNAs were isolated 48 h post-treatments and subjected to the RT-PCR detection of survivin mRNA
Effect of DHA and ionizing radiation on caspase-3 activation
To evaluate whether the growth inhibition that was observed in HT-29 cells is accompanied by caspase-3 activation, we evaluated caspase-3 activation in treated cells. By doing so, we found that pretreatment with DHA (50- and 100-μM) followed by γ-irradiation (2- or 10-Gy) increased caspase-3 activation. Specifically, pretreatment with 100-μM DHA in combination with 10 Gy resulted in 5.1-fold increase in the caspase-3 activation compared with vehicle (Fig. 5). We also found that enzymatic caspase-3 activity was lower in cells treated with 2- or 10-Gy of γ-irradiation compared with other treatments.
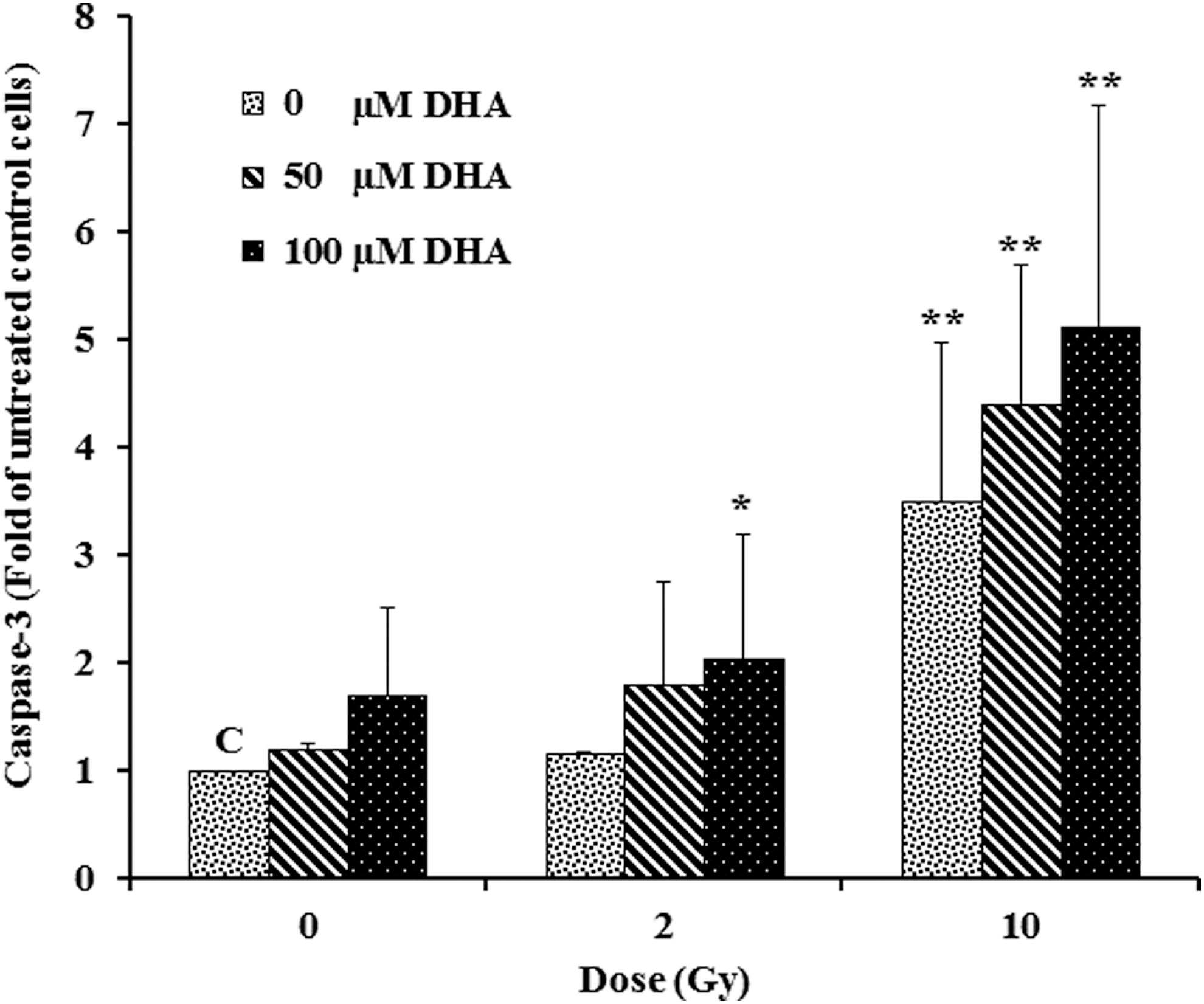
Effect of DHA alone or in combination with ionizing radiation on caspase-3 activation. About 5 × 10 5 HT-29 cells per well were treated either with DHA (50- and 100-μM) alone for 48 h or pretreated with 50- and 100-μM DHA for 4 h followed by 2- or 10-Gy of γ-irradiation, after which caspase-3 activation was measured using FIENA method 48 post-treatments. The results represent mean ± SD from two independent experiments with duplicate assays. *p < 0.05 and **p < 0.01 versus untreated control cells (C; Vehicle). DHA, docosahexaenoic acid; SD, standard deviation.
Effect of DHA and ionizing radiation on apoptosis induction
To investigate apoptosis and necrosis induction, samples were stained with Annexin V conjugated with FITC and PI and subjected to the flow cytometer. Induction of apoptosis by pretreatment with DHA (50- and 100-μM) in combination with γ-irradiation was confirmed, which showed a decrease in live cells by induction of apoptosis. In addition to apoptosis, a necrotic process was also involved in HT-29 cells death. Specifically, pretreatment with 100-μM of DHA followed by 10-Gy of γ-irradiation induced an increase in the number of necrotic cells (35.9%) compared with untreated cells (Table 4). Comparison of necrosis induction between DHA and γ-irradiation revealed that γ-irradiation induces significant increases in necrotic cell death compared with the cells treated with DHA only (Table 4).
Effect of Docosahexaenoic Acid and γ-Irradiation Alone or in Combination on Apoptosis and Necrosis Induction
5 × 10 5 HT-29 cells per well were treated either with DHA at concentrations of 50- and 100-μM alone for 48 h or pretreated with DHA at concentrations of 50- and 100-μM for 4 h before 2- or 10-Gy of γ-irradiation. Subsequently, after 48 h, the cells were harvested and stained with Annexin V-FITC/PI. Data represent at least means of two experiments ± SD.
p < 0.05, ** p < 0.01 and *** p < 0.001 versus untreated control cells (Vehicle).
DHA, docosahexaenoic acid; FITC, Fluorescein Isothiocyanate; PI, propidium iodide; SD, standard deviation.
The DRF calculated for the aforementioned concentrations of DHA in combination with different doses of γ-rays is indicative of its potent radiosensitizing capability and revealed that pretreatment of cells with 100-μM DHA for 4 h before 2 Gy of γ-irradiation is more effective than other treatments at the same conditions (Table 5).
Effect of γ-Irradiation in Combination with Docosahexaenoic Acid on Dose Reduction Factor
5 × 10 5 HT-29 cells per well were pretreated with DHA at concentrations of 50- and 100-μM for 4 h before 2- or 10-Gy of γ-irradiation. Subsequently, 48 h post-treatments, the cells were harvested and induction of apoptosis (Early and Late) was determined by flow cytometer.
DRF was calculated as the ratio of apoptosis induced by radiation alone to radiation-induced apoptosis in the presence of DHA.
DHA, docosahexaenoic acid; DRF, dose reduction factor.
Correlations between survivin expression, caspase-3 activation, and apoptosis
In treated cells, mRNA of survivin level was downregulated by DHA and γ-irradiation alone or in combination. There were associations between expression of survivin with caspase-3 activation and apoptosis. The decreases in survivin mRNA levels were accompanied by increases in caspase-3 activation and apoptosis compared with vehicle. Significant correlations were observed between survivin mRNA levels with caspase-3 activation and apoptosis. Furthermore, we found that, there were significant correlations between caspase-3 activation and apoptosis (Table 6).
Evaluation of Correlation Between Survivin Expression, Caspase-3 Activation, and Apoptosis
5 × 10 5 HT-29 cells per well were treated either with DHA at concentrations of 0-, 50- and 100-μM alone for 48 h or pretreated with DHA at concentrations of 50- and 100-μM for 4 h before 2- or 10-Gy of γ-irradiation. Subsequently, after 48 h, the cells were harvested and survivin mRNA levels, caspase-3 activation, and total apoptotic rates were determined. Next, correlations between variables were calculated using Spearman correlation analysis.
DHA, docosahexaenoic acid.
Discussion
P53-mutant CRC cells are more resistant to radio- and/or chemotherapy. In this study, we evaluated the effects of DHA on HT-29 cells that serves as a model for homozygous mutant-p53 CRC cells with radioresistant properties. 20 It is interesting to note that, DHA at concentrations of 50- and 100-μM equal to the levels of DHA achievable in the human body following supplementation of the diet with different amount of omega-3 PUFAs/d 18 decreased cell numbers and cell proliferation rates and increased sensitivity of mutant-p53 HT-19 cells to γ-irradiation irrespective p53 mutation. These results were obtained using DHA concentrations that were below the maximum tolerated dose of omega-3 PUFAs reported in human, 22 suggesting that DHA would be a safe and nontoxic compound to sensitize CRC cells to radiotherapy in patients with homozygous mutant-p53 CRC cells.
In a similar study, Cai et al. showed a simple additive effect of DHA with radiation in HT-29 cells. 23 The discrepancy between Cai et al. work and our work could be attributed to the different radiation doses and DHA concentrations used in our study compared with Cai et al. work.
PUFAs induce oxidative stress to facilitate tumor cell killing. 24 The same mechanism has been attributed to the ionizing radiation. 25 Therefore, additional damage of tumor cells can be expected by combining DHA with ionizing radiation such as ones in our study. This cooperation of DHA and irradiation is encouraging for a potential utilization of DHA/γ-irradiation in clinical practice.
To evaluate cell viability postradiotherapy, we performed the MTT survival assay instead of the clonogenic assay. Although the clonogenic assay is commonly used to evaluate survival of irradiated cancer cells, this method has some disadvantages such as the time taken to form colonies, inability to measure survival in cells that do not grow as colonies, risk of contamination of cells over long period of cultivation, and low sample throughput. Therefore, as an alternative method, MTT assay, that is based on mitochondrial activity of live cells with several advantages including high sensitivity, rapid, low cost, and high reproducibility with ability to measure metabolic activity in cells that do not grow as colonies, was used in our study. In this regard, several studies have been reported regarding the comparability of MTT and clonogenic assay. 26,27
Flow cytometry assays showed dramatic increases in apoptosis and necrosis rates after treatment with DHA/γ-irradiation. It has been shown that PUFAs and γ-irradiation may induce apoptosis through generation of reactive oxygen substance and lipid peroxidation products. 28,29 Additionally, lipid peroxide generation was shown to induce necrosis through oxidative nucleic acid base modification and DNA strand breaks. 30,31 In line with these findings, our data showed that DHA in combination with γ-irradiation increased apoptosis and necrosis.
In this study, DHA alone induced apoptosis in HT-29 cell-line, which is p53 deficient. 30,31 Our study showed that DHA induces apoptosis through a p53-independent pathway. This result is in agreement with previous report showing that DHA inhibits growth of CRC cells independent of p53 mutational status. 16 As, this protein is frequently mutated in most human malignancies, which is associated with poor prognosis and resistance to ionizing radiation, 32 –34 this could demonstrate the advantage of using DHA as an attractive radiosensitizer agent in mutant-p53 tumor cells compared with other compounds.
Survivin protects cells from apoptosis by binding to activated form of caspase-3 via the inhibitor of apoptosis domain, thereby inhibiting its activity. 35 DHA in combination of γ-irradiation decreased survivin mRNA levels, which were followed by caspase-3 activation and apoptosis. Based on our descriptive results, induction of caspase-3 activation through inhibition of the survivin expression appears to be one of the molecular pathways leading to DHA apoptotic effect in HT-29 cells. In this context, researchers showed that DHA decreases survivin expression through proteasomal regulation of beta-catenin levels in CRC cell lines. 36
While this study lacks a mechanistic insight as to how DHA exerts modulation of survivin, it does provide an interesting basis to extent the study with in vivo experiments.
We also found that, the level of survivin mRNA was higher in the cells treated with 10 Gy compared with 2 Gy of γ-irradiation. In agreement with our result, Jin et al. showed that γ-irradiation induces upregulation of survivin in HepG2 cells in a dose-dependent manner. 37 Interestingly, when DHA in combination with γ-irradiation was employed, the levels of survivin mRNA dramatically decreased compared with untreated cells, which suggests that DHA in cooperation with γ-irradiation turns the profile of high level of survivin expression in HT-29 cells into that of normal cells.
Conclusions
DHA decreases survivin mRNA level and activates caspase-3 irrespective of p53 status. Significant decreases in ED50 at concentrations of DHA equal to human plasma levels, representing a safe radiosensitizer compound for radioresistant mutant-p53 CRC cells. The additional properties of DHA on angiogenesis, inflammation, metastasis, invasion, epigenetic regulation, 38 ease of oral administration, protective effect on normal tissues during radiotherapy, 39 and lack of toxicity in normal cells, might further justify its potential as an attractive radiosensitizer drug in patients with radioresistant mutant-p53 CRC cells.
Footnotes
Acknowledgment
The authors acknowledge Dr. Talya Dayton from Hubrecht Institute for her valuable suggestions with respect to the article.
Disclosure Statement
No competing financial interests exist.