
Abstract
Cholangiocarcinoma (CCA) is a highly malignant and aggressive tumor of the bile duct that arises from epithelial cells. Chemotherapy is an important treatment strategy for CCA patients, but its efficacy remains limited due to drug resistance. Salubrinal, an inhibitor of eukaryotic translation initiation factor 2 alpha (eIF2α), has been reported to affect antitumor activities in cancer chemotherapy. In this study, the authors investigated the effect of salubrinal on the chemosensitivity of doxorubicin in CCA cells. They showed that doxorubicin induces CCA cell death in a dose- and time-dependent manner. Doxorubicin triggers reactive oxygen species (ROS) generation and induces DNA damage in CCA cells. In addition, ROS inhibitor N-acetylcysteine (NAC) pretreatment inhibits doxorubicin-induced CCA cell death. Importantly, these data demonstrate a synergistic death induction effect contributed by the combination of salubrinal and doxorubicin in CCA cells. It is notable that salubrinal promotes doxorubicin-induced ROS production and DNA damage in CCA cells. Taken together, these data suggest that salubrinal enhances the sensitivity of doxorubicin in CCA cells through promoting ROS-mediated DNA damage.
Introduction
Cholangiocarcinoma (CCA), is the second most common hepatic malignant tumor beside hepatocellular carcinoma, arising from the epithelium of intrahepatic and extrahepatic biliary ducts. 1 –3 Although chemotherapy is a common stratagem used for CCA therapy, the worldwide morbidity of CCA has unexpectedly increased since 1980s and the prognosis of patients with 5-year survival rate is <10%. 2,4 Accumulating evidences reveal that the development of chemotherapeutic drug resistance is a major reason for the poorly therapeutic effects of CCA chemotherapy. 5 –8 Doxorubicin, a widely used efficiently chemotherapy agent in a broad range of tumors, 9,10 functioned by intercalating double-stranded G-C sequences of DNA and inhibiting the synthesis of nucleic acids, resulting in DNA damage and epigenome and transcriptome deregulation. 11,12 However, the response rate of doxorubicin is not satisfactory in CCA owing to drug resistance. 12 Hence, improving the drug sensitivity of doxorubicin might to be an important strategy to enhance the survival rate of CCA patients. 13
Salubrinal, a small molecular compound, inhibits the activity of eukaryotic translation initiation factor 2 alpha (eIF2α) through the selective inhibition of eIF2α dephosphorylation. 14 The authors and others have previously demonstrated that salubrinal could affect the sensitivity of some chemotherapy agents, such as cisplatinum and doxorubicin in several types of tumor cells, including hepatocellular carcinoma and breast cancer cells. 15 –17 However, the effects of salubrinal on the chemotherapy sensitivity of doxorubicin in CCA have not been explored.
In this study, the authors investigated the effects of salubrinal on the chemosensitivity of doxorubicin in CCA cells. They found that salubrinal improved CCA cell death upon doxorubicin treatment through increasing reactive oxygen species (ROS) production and promoting DNA damage. These results implied a potential synergistic effect of salubrinal in combination with doxorubicin on CCA cells, suggesting the combination of salubrinal and doxorubicin might be a novel option to CCA therapy.
Materials and Methods
Chemicals and antibodies
Doxorubicin and eIF2α inhibitor salubrinal (Sal) were purchased from Sigma Chemical Company. ROS scavenger N-acetylcysteine (NAC) was purchased from BBI Life Science. Antibodies against cleaved PARP (c-PARP), cleaved caspase-3 (c-Caspase-3), p-CHK1, p-CHK2, p-H2AX, eIF2α, and p-eIF2α were purchased from Cell Signaling Technology. Antibody GAPDH was purchased from Santa Cruz Biotechnology.
Cell culture
CCA cell lines RBE and QBC939 were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin in a humidified incubator containing 5% CO2 and 95% ambient air at 37°C. 18
Western blot analysis
Experimental cells were lysed with Triton lysis buffer (20 mM Tris, pH 7.4, 137 mM NaCl, 10% glycerol, 1% Triton X-100, 2 mM EDTA, 1 mM PMSF, 10 mM NaF, 5 mg/mL aprotinin, 20 mM leupeptin, and 1 mM sodium orthovanadate) and centrifuged at 12,000 g for 15 min. The concentrations of protein samples were measured using the BCA assay, then denatured with 4 × sodium dodecyl sulfate (SDS)-loading buffer (200 mM Tris, pH 6.8, 8% SDS, 400 mM DTT, 0.4% bromophenol blue, and 40% glycerol) at 100°C for 5 min. The protein samples were subjected to Western blot analysis with standard sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) as previously described. 19 The proteins were electrolytically transferred to NC membrane and blocked within 5% nonfat milk, dissolving TBST at room temperature for 1 h. Then, the membrane was incubated overnight at 4°C with primary antibodies against c-PARP (1:1000), p-CHK1 (1:1000), p-CHK2 (1:1000), p-H2AX (1:1000), or GAPDH (1:200). Subsequently, the relevant secondary antibodies labeled with IRDye 700 were employed to recognize the target protein before being developed with the Odyssey system.
LDH release assay
RBE or QBC939 cells (4 × 103) were seeded into 96-well plates and cultured overnight, and then treated with doxorubicin diluted in RPMI-1640 containing 1% FBS for indicated times. The released LDH was examined by LDH cytotoxicity assay kit in accordance with the manufacturer's instructions, and the absorbance was determined with BioTek microplate reader at 450 nm. In addition, the cell death ratio was calculated as follows: cell death ratio = 100 × (experimental release-spontaneous release)/(maximum release-spontaneous release). The value of maximum release was tested with the treatment of maximum release solution in the kit. 20 The experiments in this assay were performed thrice.
ROS assay
The intracellular ROS assay was measured as recommended by the manufacturer's instructions. Briefly, ∼2 × 105/well RBE or QBC9393 cells were seeded into six-well plates, cultured overnight, and treated with corresponding chemical dissolved in RPMI-1640 containing 1% FBS for 24 h. 21 Then, experimental cells were cultured with serum-free culture medium containing HCFH-DA and incubated for 1 h. Next, the stained cells were washed with serum-free culture medium and pictures were taken using fluorescence microscope. Finally, ROS was detected by BioTek microplate reader (excited at 488 nm and emitted at 525 nm).
Statistical analysis
All experiments were performed thrice independently. These quantitative data were shown as the mean ± standard deviation. Statistical analysis was evaluated using Student's t-test. p < 0.05 was considered statistically significant.
Results
Doxorubicin induces CCA cell death
To investigate the potential role of doxorubicin in the induction of cell death in human CCA cells, RBE and QBC939 cells were used to determine the cytotoxic effects of doxorubicin. LDH analysis showed that doxorubicin treatment induced RBE and QBC939 cell death in a dose- and time-dependent manner (Fig. 1A, C). To further determine whether doxorubicin could influence cell apoptosis in CCA cells, Western blot assay was used to measure the c-PARP and c-caspase-3 in RBE and QBC939 cells. The results showed that doxorubicin treatment induced c-PARP in a dose- and time-dependent manner (Fig. 1B, D). These data suggested that doxorubicin can induce human CCA cell death.
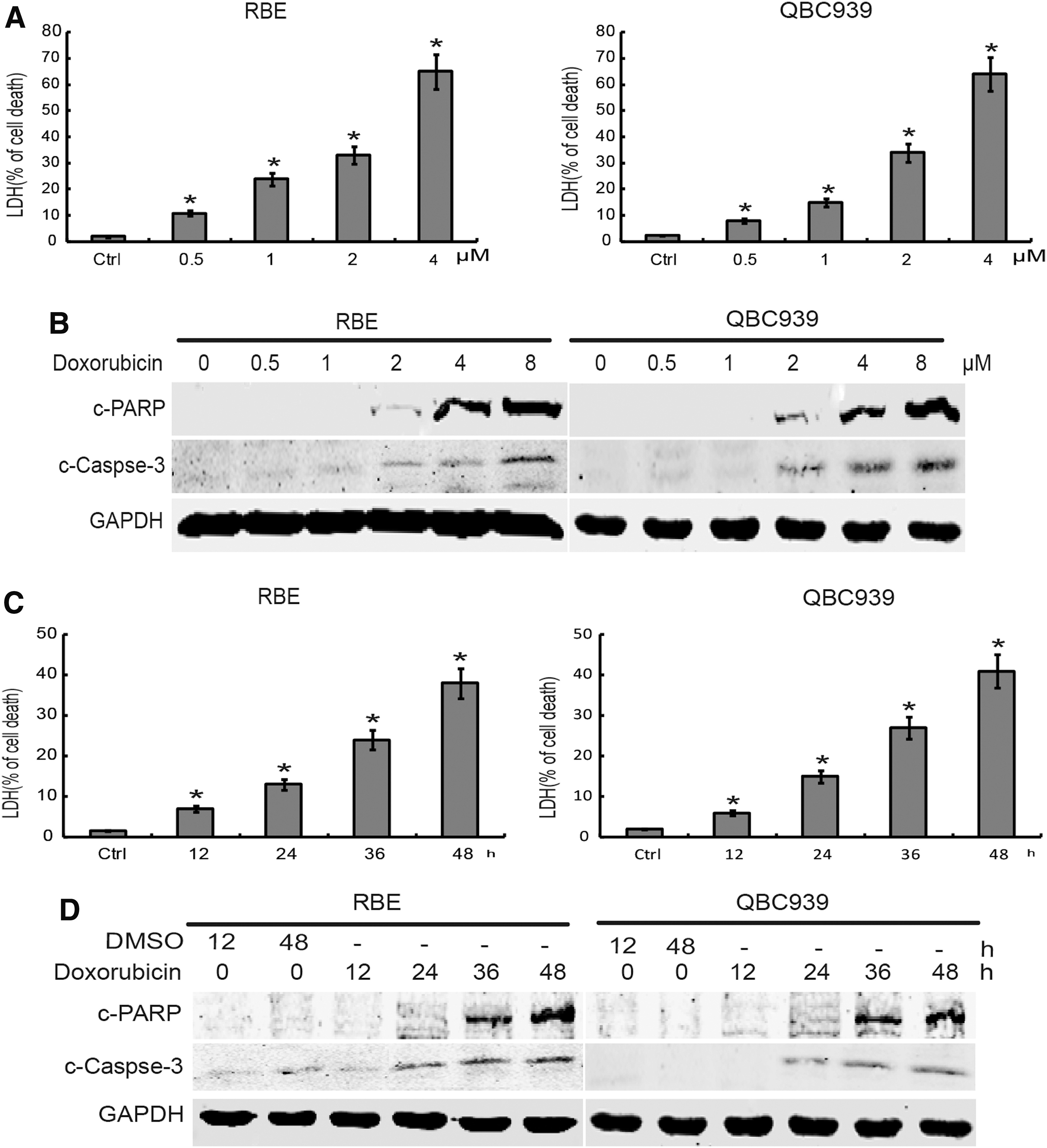
Doxorubicin induces the CCA cell death.
Doxorubicin initiates ROS production and DNA damage in CCA cells
To evaluate whether doxorubicin induced ROS production in CCA cells, ROS assay was performed in RBE and QBC939 cells. The results showed that doxorubicin treatment significantly elevated ROS levels in RBE and QBC939 cells (Fig. 2A). To explore whether doxorubicin treatment caused DNA damage in CCA cells, DNA damage biomarkers were evaluated with immunoblotting. As shown in Figure 2B, the abundance of phospho-H2AX, phospho-CHK1, and phospho-CHK2 significantly increased upon doxorubicin treatment in RBE and QBC939 cells. These data indicated that doxorubicin induces ROS production and DNA damage in CCA cells.
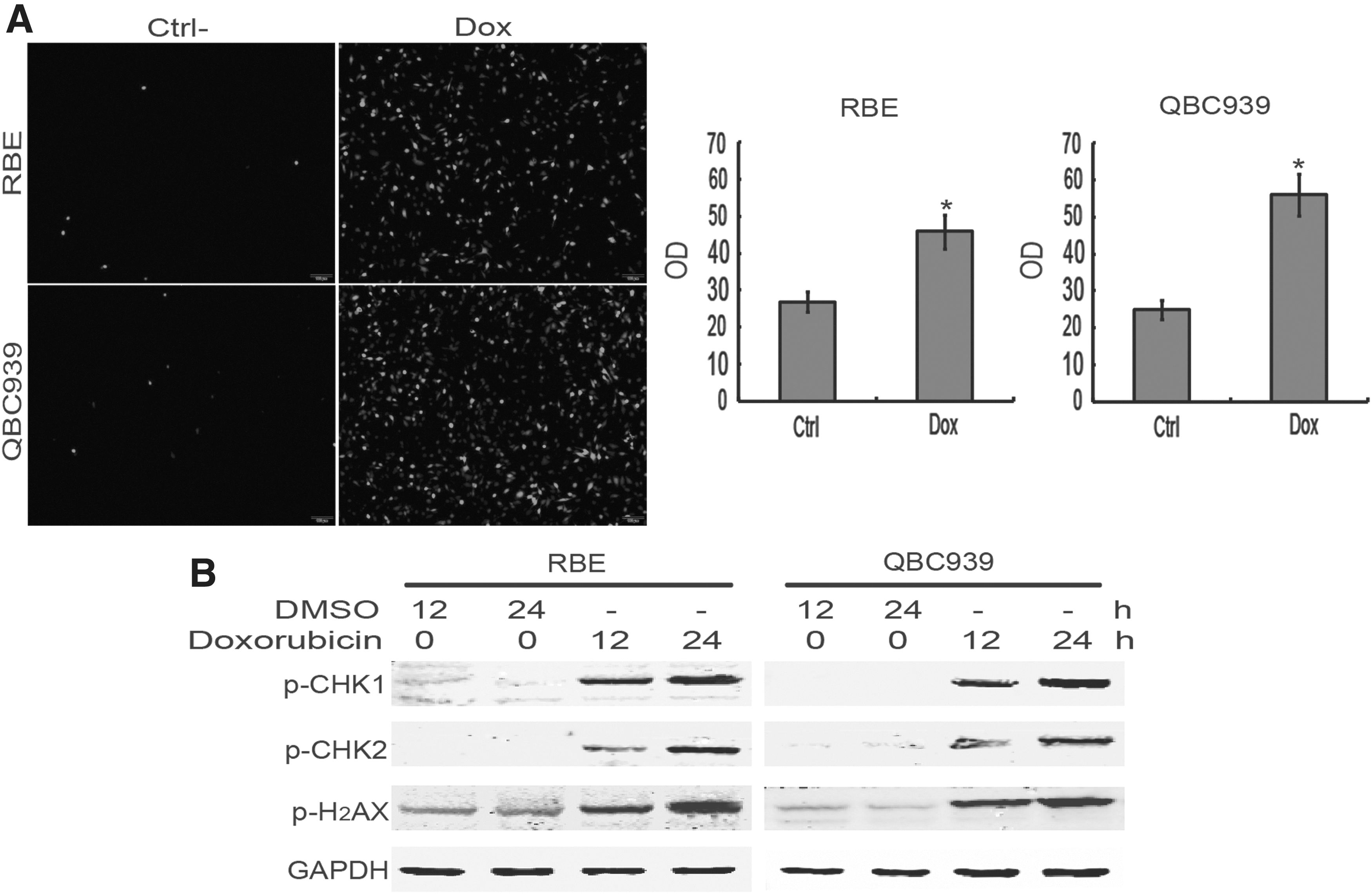
Doxorubicin initiates ROS production and DNA damage in CCA cells.
NAC inhibits doxorubicin-initiated CCA cell death
To investigate the effect of ROS induced by doxorubicin on the cytotoxicity of human CCA cells, the antioxidant NAC was used to suppress ROS. As shown in the data, NAC pretreatment significantly reduced doxorubicin-induced LDH release (Fig. 3A) and PARP and caspase-3 cleavage (Fig. 3B) in RBE and QBC939 cells, suggesting that a reduction of ROS can alleviate doxorubicin-induced CCA cell death. Moreover, the Western blot results showed that doxorubicin-induced DNA damage was obviously inhibited by NAC in doxorubicin-treated cells (Fig. 3C), indicating NAC can protect RBE and QBC939 cells against doxorubicin-induced DNA damage. These results together revealed that ROS promoted the death of CCA cells upon doxorubicin treatment.

NAC inhibits doxorubicin-initiated CCA cell death.
Salubrinal increases doxorubicin chemosensitivity in CCA cells
To investigate the effect of salubrinal on the cytotoxicity in CCA cells, the death of CCA cells was tested upon combination treatment of salubrinal and doxorubicin. These results showed that salubrinal pretreatment obviously increased doxorubicin-induced LDH release (Fig. 4A) and PARP and caspase-3 cleavage (Fig. 4B) in RBE and QBC939 cells. Thus, these data indicated that salubrinal increased the chemosensitivity of doxorubicin in CCA cells.
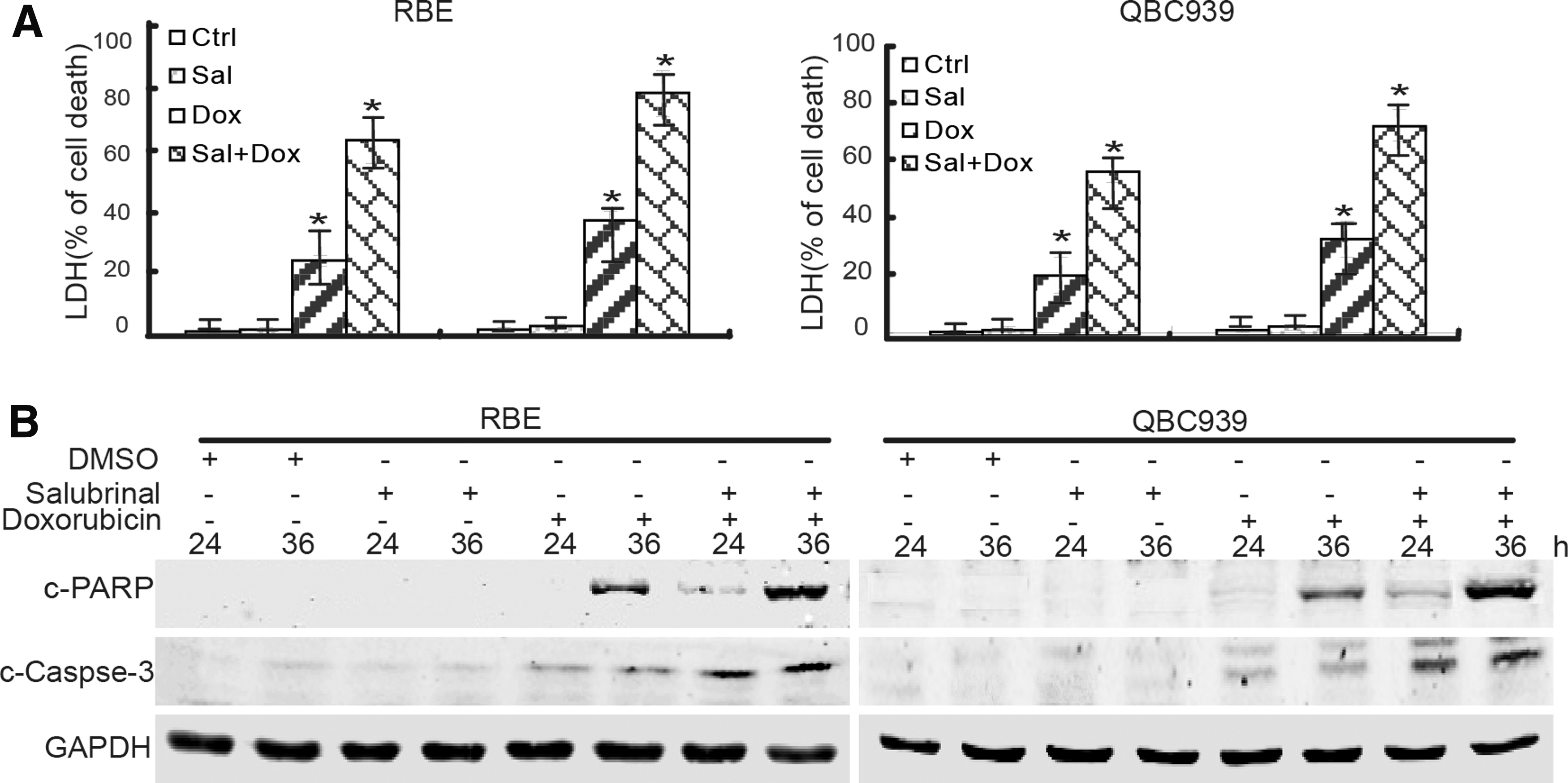
The eIF2α inhibitor increases doxorubicin chemosensitivity in CCA cells.
Salubrinal promotes the cytotoxicity of doxorubicin by increasing DNA damage
Further investigations were conducted to explore the mechanisms of salubrinal in promoting doxorubicin-mediated human CCA cell death. These data showed that salubrinal pretreatment increased ROS levels in doxorubicin-treated RBE and QBC939 cells (Fig. 5A, B). Moreover, Western blot results showed that salubrinal pretreatment increased the expression of phosphor-H2AX, phosphor-CHK1, and phosphor-CHK2 in RBE and QBC939 cells (Fig. 5C), indicating salubrinal promoted DNA damage and DNA damage response. Taken together, these results indicated that salubrinal can increase the chemosensitivity of doxorubicin by increasing ROS production and promoting DNA damage in CCA cells.
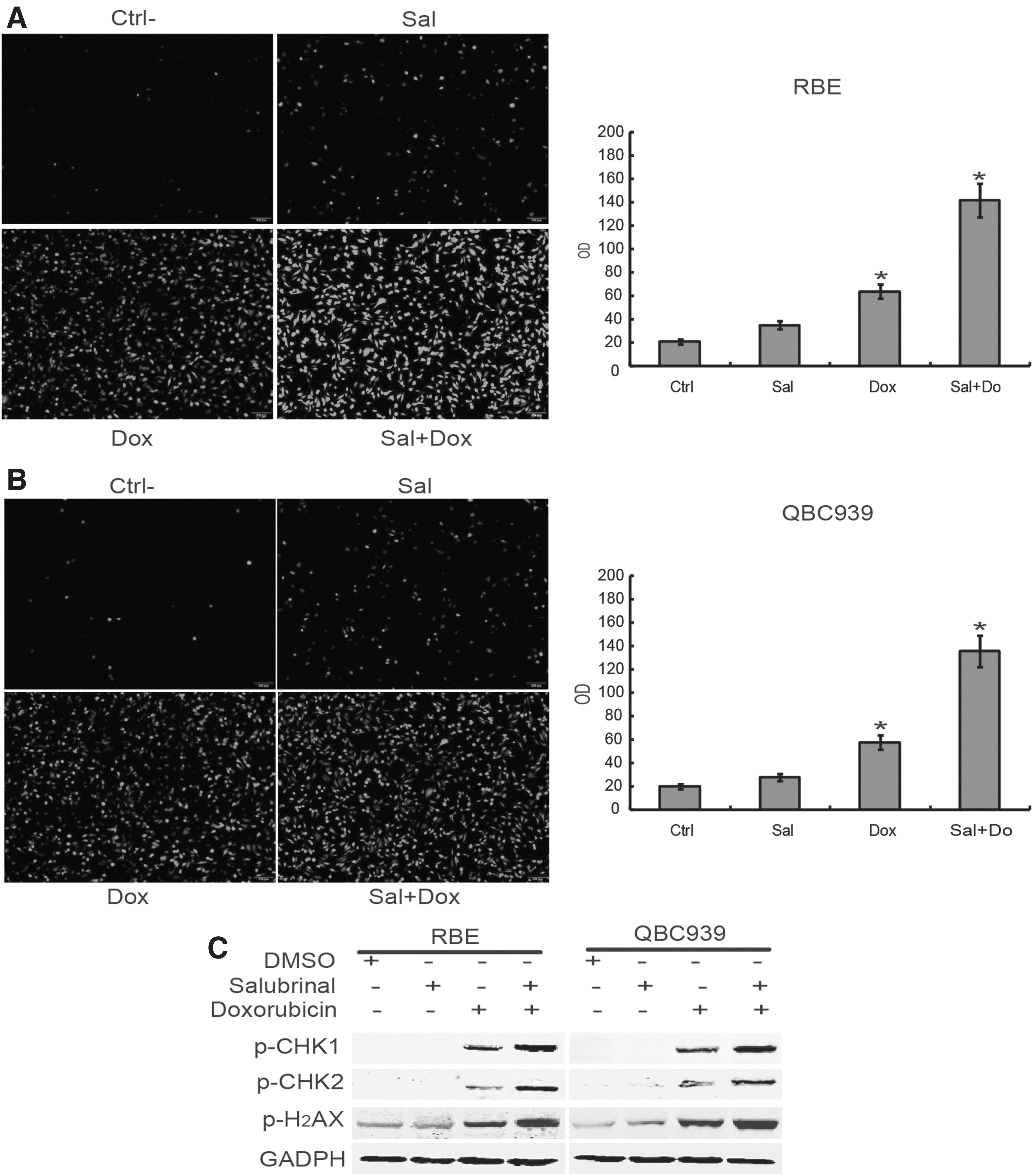
Salubrinal promotes cytotoxicity in of doxorubicin by increasing ROS and DNA damage.
Discussion
CCA is one of the most aggressive and lethal malignancies in humans. 22 For CCA patients, chemotherapy is an essential therapeutic method, especially for those without the chance of surgery treatment. A key factor in the development of chemotherapy treatment is to selectivity improve drug sensibility. 23 Salubrinal, the inhibitor of eIF2α, was reported to affect the sensitivity of some chemotherapy agents in some types of cancer cells. 24 However, whether salubrinal could affect the cytotoxicity of doxorubicin in CCA cells is still unknown. In this study, the authors demonstrated that salubrinal promoted doxorubicin-medicated CCA cell death through increasing ROS production and promoting DNA damage.
These data showed that doxorubicin induced CCA cell death in a dose- and time-dependent manner. Accumulating evidences have demonstrated that ROS induction plays an important role in the cytotoxicity of doxorubicin. 10,25 –27 Furthermore, the authors found that doxorubicin treatment causes an increase of ROS in CCA cells. Moreover, the findings that ROS scavenger NAC markedly decreased the LDH release and reduced PARP cleavage strongly suggest that ROS is involved in doxorubicin-induced CCA cell death.
Considering that DNA damage is one of the most serious effects of ROS overproduction, the authors wondered whether DNA damage exerts a role in doxorubicin-induced CCA cell death. In this study, they found that doxorubicin treatment induces DNA damage in CCA cells. Mechanistically, these results suggest that DNA damage induction is implicated in CCA cell death upon doxorubicin treatment. In addition, doxorubicin-induced DNA damage was significantly inhibited by NAC in CCA cells, indicating that doxorubicin triggered DNA damage depending on ROS induction.
The previous studies found that the eIF2α pathway is deregulated in CCA cells and eIF2α inhibitor salubrinal can inhibit the proliferation of CCA cells.
15,28
Moreover, the authors have demonstrated that salubrinal increased the antitumor activity of rapamycin in CCA cells, but reduced cisplatin chemosensitivity in hepatocellular carcinoma cells.
15,17
A recent report revealed that salubrinal increased doxorubicin sensitivity in breast cancer cells.
16
In this study, the authors wondered whether salubrinal could affect the chemosensitivity of doxorubicin in CCA cells. They found that salubrinal obviously increased the cytotoxicity of doxorubicin in CCA cells, demonstrated by elevated LDH release and PARP cleavage. These results suggest that salubrinal can enhance the antitumor efficiency of doxorubicin in CCA cells. However, the phosphorylation of eIF2α changed obviously after salubrinal treatment (Supplementary Fig. S1; Supplementary Data are available online at
Considering ROS plays an indispensable role in doxorubicin-induced CCA cell death, the authors assumed that salubrinal might promote the antitumor efficiency of doxorubicin through affecting ROS generation. This assumption is supported by the data that salubrinal pretreatment resulted in increased ROS induction in doxorubicin-treated CCA cells. It is notable that salubrinal treatment induced ROS production without DNA damage in CCA cell induction. In addition, salubrinal promoted doxorubicin-mediated DNA damage in CCA cells. Based on these findings, the authors proposed that salubrinal exerts a promotion role, at least in part, in doxorubicin chemosensitivity through increasing ROS induction and DNA damage. However, further studies are needed to uncover the detailed mechanisms for salubrinal-mediated ROS production in CCA cells.
In conclusion, the authors found that doxorubicin induces CCA cell death through initiating ROS production and DNA damage, which could be aggravated by eIF2α inhibitor salubrinal. These findings shed light on the molecular basis of salubrinal in combination with doxorubicin, which might be a new strategy for the treatment of CCA.
Footnotes
Acknowledgments
This work was supported by grants from Science and Technology Department of Sichuan Province Foundation (2017JY0134), Health and Family Planning Commission of Sichuan Province Foundation (16PJ539), the National Natural Science Foundation of China (81472312), Innovation Team of Education Department of Sichuan Province (16TD0021), Luzhou City, Southwest Medical University Foundation (2016LZXNYD-T02, 2015LZCYD-S01-14/15, 2015LZCYD-S01-8/15), Sichuan Province, Luzhou City, and Southwest Medical University Foundation (14JC0082, 14JC0038, 14ZC0070).
Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.