
Abstract
Background:
Paeonol, a phenolic component from the root bark of Paeonia moutan, has been identified to possess antitumor effects. However, the effect of paeonol and the mechanism of CXCL4/CXCR3-B signals in paeonol-induced breast cancer cell remain unknown.
Materials and Methods:
After MDA-MB-231 cells were pretreated with paeonol or DMSO, the proliferation activity was detected by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide), Hoechst, Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL), and Annexin-V/propidium iodide staining flow cytometry. Western blot and immunohistochemistry of human breast cancer and noncancerous tissues were performed to determine the molecular alteration of CXCL4/CXCR3-B signals.
Results:
Compared with the control, paeonol-treated breast cancer cells had low proliferation activity and high apoptotic index, indicating that paeonol induces breast cancer cell apoptosis. Western blot and immunohistochemistry showed that paeonol increased CXCR3-B signal, downregulated CXCL4, heme oxygenase (HO-1) with a corresponding increased BACH1, and decreased nuclear factor E2-related factor 2 (Nrf2).
Conclusions:
Thus, CXCL4/CXCR3-B may be involved in the mechanism of apoptosis induced by paeonol in breast cancer cells by regulating the expression of BACH1 and Nrf2 to downregulating HO-1 and promote apoptosis. Therefore, the authors suggest paeonol has a significant growth inhibitory effect on breast cancer cells, which may be related to the induction of apoptosis.
Introduction
B
Chemokines and their receptors participate in the development of cancers, including tumor cell proliferation, angiogenesis, invasion, and metastasis, as well as affect the penetration of tumor immune cells. CXCR3 is a G protein-coupled receptor 9 (GPR9) that belongs to the CXC chemokine receptor family and consists of CXCR3-A and CXCR3-B isoforms in human. 5 CXCL4 binds specifically to CXCR3-B, whereas CXCL9, CXCL10, and CXCL11 bind to both CXCR3-A and CXCR3-B. 6,7
CXCR3-A and CXCR3-B control opposite biological roles using distinct intracellular signals. 7 –9 Generally, activation of CXCR3-A enhances tumor cell growth, invasion, and survival, whereas CXCR3-B appears to cause proliferation suppression, apoptosis, and angiostasis. Report has revealed that renal cancer cell proliferation is inhibited when CXCR3-B is overexpressed. 10 Moreover, renal cancer cells promote growth and migration via CXCR3-A-controlled signal in the absence or low presence of CXCR3-B. 11
Heme oxygenase (HO-1), a cytoprotective enzyme, plays a critical role in maintaining cellular homeostasis during stress conditions to break down heme into biliverdin, ferrous iron, and carbon monoxide. 12 Recently, HO-1 has been implicated in cancer promotion. 13 In cancers, highly expressed HO-1 and nuclear factor E2-related factor 2 (Nrf2), its positive regulator, play a crucial role by regulating prosurvival and antiapoptotic pathways for tumor cell growth. 14
HO-1 is activated by small Maf proteins and Nrf2 heterodimers via binding to Maf recognition elements, while its transcription is suppressed by BACH1 heterodimers and small Maf proteins such as MafK. 15 Thus, there is either an increased nuclear import or export of BACH1 and Nrf2 to control HO-1 based on the received signal. Both mRNA and protein level HO-1 are overexpressed in renal cancer tissues expressing low amounts of CXCR3-B. 11 CXCR3-B proliferation inhibition events in breast cancer cells are controlled by decreasing HO-1 expression to promote increased apoptosis. 16,17
Through research advancement, natural products of medicinal plants have revealed their anticancer potentials that may be used in cancer chemotherapy. Paeonia moutan is a plant that commonly grows in the Southwestern and Northwestern parts of China and has long been used as traditional Chinese herbal medicine. The root bark of Paeonia moutan contains a secondary bioactive component paeonol. Paeonol is a phenolic composite that possesses a number of pharmacological and physiological properties such as antioxidation, 18 antiallegic, 19 antioxidant, 20 anti-inflammation, 21,22 and antiangiogenic. 23 The administration of paeonol for both in vivo and in vitro studies has revealed antiproliferative and apoptosis-inducing effects in several cancer cell lines. 24,25
There is evidence that paeonol's anticancer effect suppressed the growth of human colorectal cancer cell line HT-29, breast cancer gene cell line T6-17, human hepatoma cell line Bel-7404, human erythromyeloid cell line K562, and cervical cancer cell line Hela. A previous study demonstrated that paeonol has anticancer effect by downregulating Bcl-2 and upregulating Bax, caspase 8, and caspase 3 via induction of apoptosis. 26 However, the therapeutic effect of paeonol via CXCL4/CXCR3-B-mediated signals and the relation to the modulation of BACH1, Nrf2, and HO-1 transcription molecules need to be clarified. This study has demonstrated that paeonol has anticancer and apoptotic effects, which may activate CXCL4/CXCR3-B-mediated growth inhibitory signals to increase BACH1 and decrease Nrf2 expression to downregulate HO-1 and promote apoptosis.
Materials and Methods
Chemicals and equipment
Paeonol was obtained from Beijing Pharmaceutical University (Beijing, China). Mouse anti-CXCR3-B, anti-CXCL4, anti-BACH1, anti-Nrf2, and anti-HO-1 monoclonal antibodies and streptavidin biotin peroxidase immunohistochemical reagent kits were obtained from Santa Cruz Bio, Inc. (Santa Cruz, CA). Horseradish peroxidase-labeled rabbit anti-goat immunoglobulin G (IgG) antibody and actin polyclonal antibody were purchased from Biosynthesis Bio. (BIOSS, Beijing). The enhanced chemiluminescence kit was purchased from Amersham Pharmacia Biotech. The MDA-MB-231 cell line was obtained from Cancer Institute of Chinese Academy of Medical Sciences. All other chemicals used were of analytical reagent grade.
Tissue samples
Breast cancer and noncancerous (control) tissue samples were collected from 114 patients without preoperative chemotherapy and/or radiotherapy, who underwent surgery at the First People's Hospital affiliated to Jiamusi University. The protocol for tissue sample collection was approved by the Research and Ethics Committee of the hospital. Informed consent to use the tissue samples was signed by patients before surgery. The specimen consisted of 114 breast cancer tissues and 30 matched noncancerous (control) tissues obtained from 30 of the 114 patients as paired specimens.
Methods
Immunohistochemical method was used to detect the expression of CXCL4, CXCR3-B, BACH1, Nrf2, and HO-1 in breast cancer and noncancerous tissues
Briefly, 4 μm formalin-fixed, paraffin-embedded mounted sections were deparaffinized, rehydrated, and rinsed three times in phosphate-buffered saline (PBS). The slides were heated in citrate solution for 40 min, cooled, and rinsed three times in PBS. Subsequently, they were incubated with 5% bovine serum albumin at 37°C for 30 min and washed again in PBS three times. The slides were incubated at 4°C with primary antibody CXCL4 (1:100 dilution) or CXCR3-B (1:100 dilution) or BACH1 (1:50 dilution) or Nrf2 (1:100 dilution) or HO-1 (1:100 dilution) overnight. The next day, the slides were incubated with biotinylated anti-rabbit IgG and (strept)avidin/biotin complex (SABC) at 37°C for 20 and 30 min, respectively. The slides were thoroughly rinsed in between incubations with PBS three times. Next, they were developed for 5 min in 3,3′-diaminobenzene (DAB) and counterstained with Mayer's hematoxylin at room temperature. Subsequently, they were washed in water, a solution of HCL in ethanol, and PBS. They were then incubated in PBS at 37°C for 40 min to 1 h. Finally, the slides were fixed in graded series of ethanol and xylene, coverslipped, and mounted under the microscope. The brown and blue stained cells under the microscope represent positive and negative cells, respectively.
Cell culture
MDA-MB-231 cells were cultured in RPMI-1640 supplemented with 10% fetal bovine serum and 1% penicillin and streptomycin. The cultured cells were kept in CO2 incubator at 5% CO2, 37°C in tissue culture flask.
Cytotoxic assay
Briefly, cells were seeded at 100 μL with 5 × 104 cells/well in 96-well plates overnight at 37°C. After a 48-h 37°C incubation with paeonol at different concentrations (4.6 × 10−5, 9.4 × 10−5, 1.8 × 10−4, 3.7 × 10−4, 7.5 × 10−4, 1.5 × 10−3 mM/mL), cells were incubated further for 4 h with 20 μL MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) (5.0 mg/mL). Following removal of the medium, 200 μL of DMSO was used to solubilize the formazan crystals in the cells. At absorbance wavelength of 490 nm, the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) formazan level was measured using Multiskan MK3 reader (Thermo Electron Corporation, San Jose, CA). Using the formula below, the cell proliferation inhibition rate was calculated.
The 50% (IC50) of drug causing inhibition of cell growth rate was then determined.
The cell cycle and apoptosis were detected by flow cytometry
Overnight 1 × 106 cells/dish were treatment with DMSO (control) or paeonol 3.6 × 10−4 mM/mL for 24 h, 48 h. Harvested cells were rinsed three times with PBS, fixed at 4°C for 30 min with 70% ethanol. After processing with RNase and propidium iodide staining for 30 min, flow cytometry assay of DNA composition and cell cycle were carried out. The percentage of apoptotic cell number and cell cycle were computed and the data analyzed by CellQuest software (Becton Dickinson, San Jose, CA).
Detection of apoptosis by Hoechst staining
The 1 × 106 MDA-MB-231 cells/dish dispensed overnight were treated with DMSO or paeonol (3.6 × 10−4 mM/mL) for 24 h, 48 h at 37°C. Subsequently, they were fixed, rinsed with wash buffer, and treated with Hoechst 33342 in accordance with the manufacturer's instruction manual.
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
Overnight cells seeded at 2 mL with 5 × 105 cells/per dish were treated with DMSO or paeonol and incubated at 37°C, 24 h, 48 h. Cells were fixed in 4% formaldehyde at room temperature for 30 min. They were rinsed with PBS and immersed in 3% H2O2 dissolved in methanol at room temperature for 10 min. Next, cells were then immersed in permeable fluid (0.1% Triton dissolved in sodium citrate) for 2 min and blotted dry with absorbent paper after two PBS washes. The cells were incubated in a dark wet reaction with 50 μL TdT at 37°C, 60 min, rinsed three times in PBS, and blotted dry. Fifty microliters of streptavidin-HRP was added and incubated at 37°C in a moist dark reaction for 30 min. Finally, after rinsing three times with PBS, DAB staining was carried out and the cells were mounted under the microscope.
Western blot expression of CXCR3-B, CXCL4, BACH1, Nrf2, and HO-1
The SDS-PAGE run protein samples were electrophoretically transferred to PVDF membranes. Milk-blocked PVDF membranes were subsequently incubated with a primary antibody (CXCL4, CXCR3-B, BACH1, Nrf2, HO-1, or β-actin), washed three times with TBST, and further incubated with the horseradish peroxidase-conjugated secondary antibody, respectively. The reactive protein bands were enhanced by chemiluminescence kit and exposed to X-ray detection film.
Statistical analysis
All values are recorded as mean ± standard deviation. Group differences were determined using one-way analysis of variance and Duncan's multiple range test. p < 0.05 was deemed statistically significant.
Results
Immunohistochemical expression of CXCL4, CXCR3-B, HO-1, BACH1, and Nrf2 in human breast cancer and noncancerous tissues
To determine the protein expression of CXCL4/CXCR3-B-mediated signaling events, 114 human breast cancer and 30 noncancerous (control) tissues were immunohistochemically evaluated. It was revealed as shown in Figure 1 that compared with the noncancerous (control) tissues, CXCL4 and HO-1 were highly expressed, while the expression of CXCR3-B was low. BACH1 was low in the nucleus with high cytoplasmic expression with a corresponding high to low nuclear and cytoplasmic Nrf2 expression, respectively, in the cancerous tissues.

The immunohistochemical expression of CXCL4, CXCR3-B, HO-1, BACH1, and Nrf2 in human breast cancer and noncancerous (control) tissue (S-P × 400).
The effect of paeonol on cell proliferation activity
To test the cytotoxic effect of paeonol on breast cancer cells, it was administered to MDA-MB-231 at different concentrations for 24 and 48 h. It was revealed that the IC50 after 24 h was more than 1.5 × 10−3 mM/mL and 48 h was about 3.6 × 10−4 mM/mL. Treatment with paeonol after 48 h had significant direct inhibition (p < 0.05) on cell proliferation (data are not shown). Subsequently, paeonol 3.6 × 10−4 mM/mL for 24 and 48 h was used for further analysis.
Hoechst and Terminal deoxynucleotidyl transferase dUTP nick end labeling assay for morphology assessment.
Considering the fact that the apoptotic process is normally correlated with DNA cleavage and morphological alteration, the authors investigated the efficacy of paeonol on nuclear morphology of MDA-MB-231. Following 48 h treatment compared with the control, paeonol resulted in fragmented and condensed nuclei population (Fig. 2).
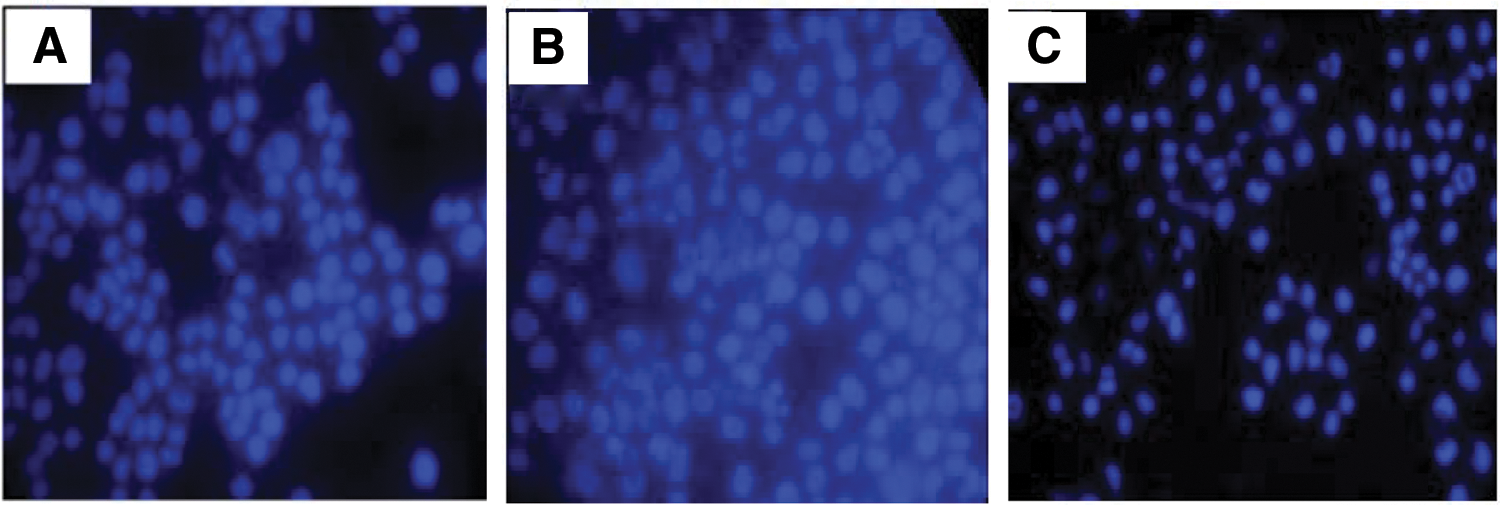
Nuclear staining assessment by Hoechst on MDA-MB-231 cells treated with ( × 400)
To buttress this, Terminal deoxynucleotidyl transferase dUTP nick end labeling staining was performed, which resulted in significant positive Terminal deoxynucleotidyl transferase dUTP nick end labeling cells characterized by fragmented nuclei, cytoplasmic shrinkage, condensed nuclei, and apoptotic bodies in the paeonol-administered cells in a time-dependent manner, as shown in Figure 3. These results attest that the inhibition by paeonol on MDA-MB-231 cells was due to apoptosis.

Morphological and apoptotic index changes of apoptosis on MDA-MB-231 cells using Terminal deoxynucleotidyl transferase dUTP nick end labeling stain ( × 400).
Paeonol induces cell cycle arrest and apoptosis on MDA-MB-231 cells
To find out the cell arrest exhibited by paeonol in the induction of apoptosis on MDA-MB-231 cells, cell cycle and Annexin V were conducted using flow cytometry. After treatment with paeonol, G0/G1 cell population was significantly increased from 48.3% in the control group to 64.7% and 68.7% in paeonol group at 24 h and 48 h, respectively. Moreover, the percentage of apoptotic cells of sub-G1 phase also increased markedly from 4.2% in the control group to 8.9% and 19.6% at 24 h and 48 h in the paeonol group, respectively. However, in the S and G2/M phases, percentage of cell population decreased when the control group was compared with the paeonol group. The results indicate that paeonol induced G0/G1 arrest on MDA-MB-231 cell (Fig. 4).
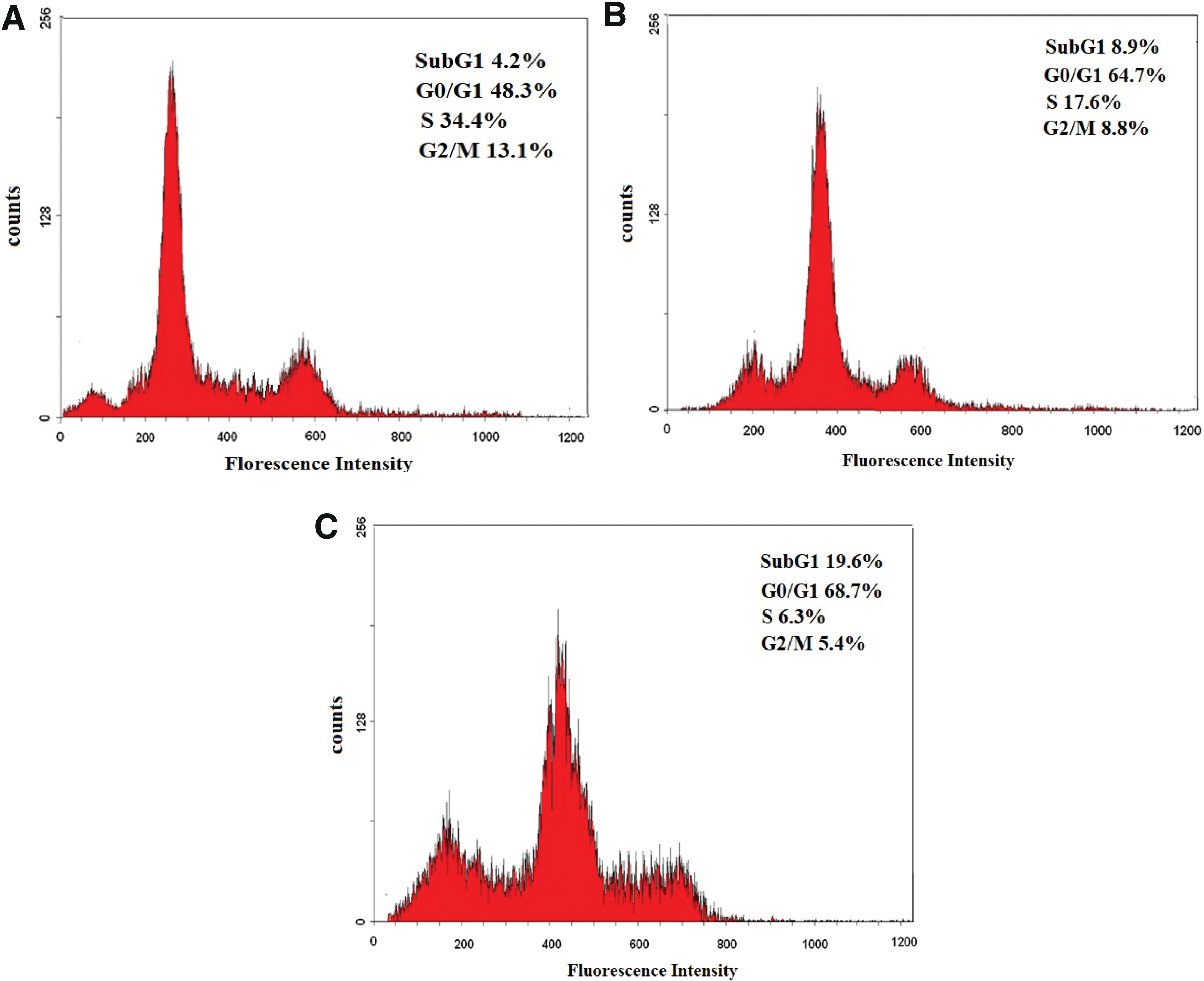
Cell cycle analysis of MDA-MB-231 cells treated with DMSO or paeonol by flow cytometry.
The authors subsequently confirmed and quantified the presence of apoptotic cells with the Annexin V-FITC Apoptosis detection kit. The results showed that the total apoptotic cell rate (early and late) increased from 9.27% (1.73% +7.54%) in the control to 33.29% (9.45% + 23.84%) at 24 h and to 44.89% (3.16% + 41.67%) at 48 h after paeonol treatment, respectively (Fig. 5).
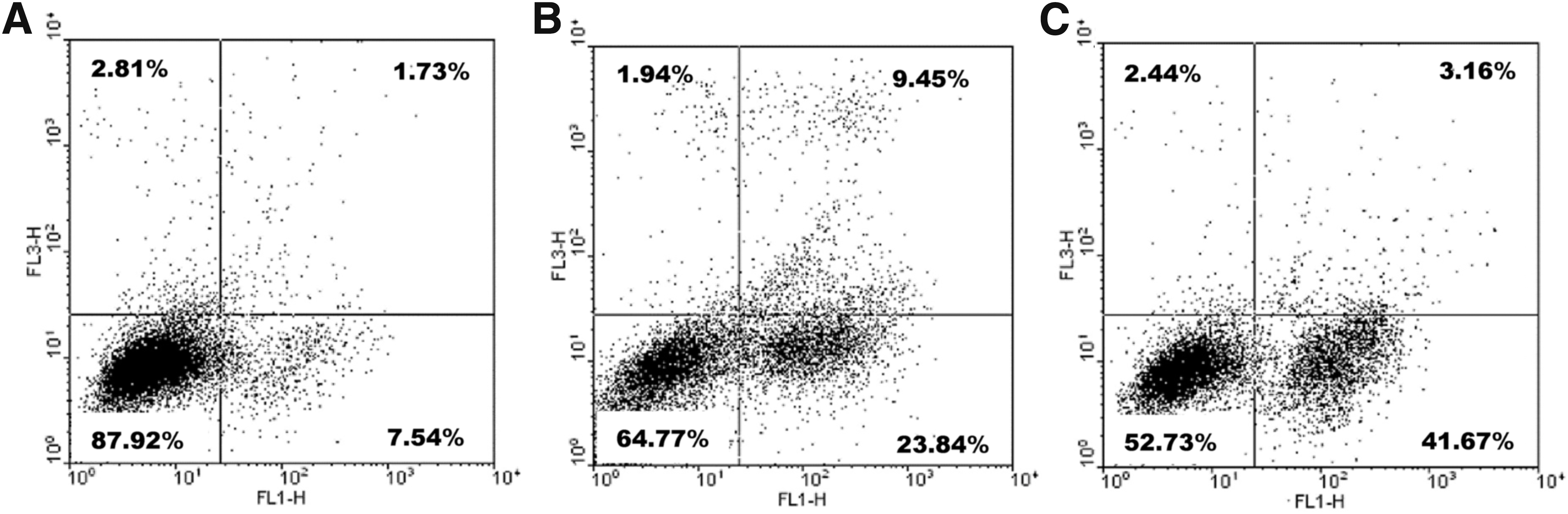
Annexin V/propidium iodide apoptosis rate detection on MDA-MB-231 cells using flow cytometry.
Effect of paeonol on CXCL4, CXCR3-B, HO-1, BACH1, and Nrf2 expression by Western blot
To determine whether CXCL4/CXCR3-B signals are involved in the antitumor effect of paeonol via the induction of apoptosis, Western blot analysis was carried out on MDA-MB-231 cells treated with DMSO and paeonol. The authors observed that compared with the control group, paeonol-treated groups resulted in increased CXCR3-B with a corresponding decreased CXCL4 and HO-1 expression. Moreover, BACH1 expression was upregulated with downregulated Nrf2 (Fig. 6).
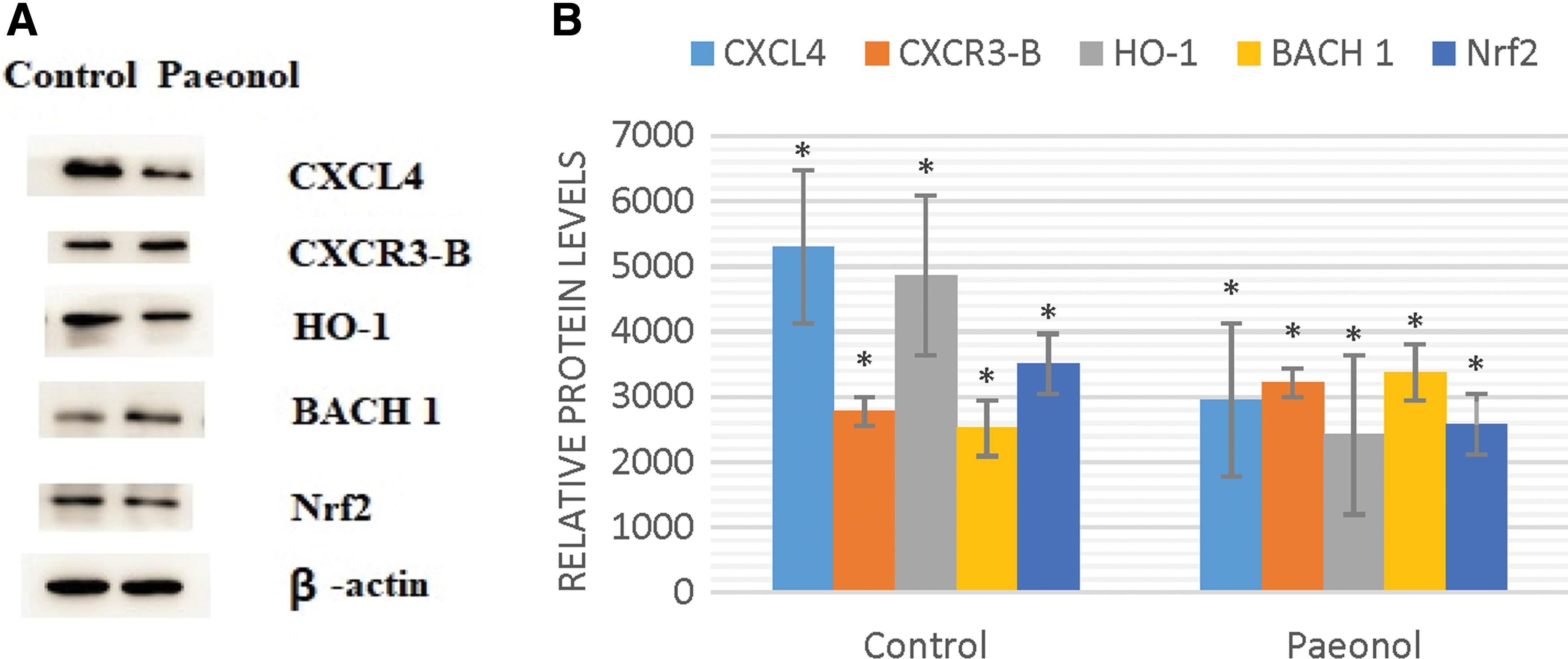
Western blot analysis for CXCL4, CXCR3-B, HO-1, BACH1, and Nrf2 in MDA-MB-231 cell line.
Discussion
Paeonol has been described with several medicinal properties and as an anticancer agent. However, the effect of paeonol on CXCL4/CXCR3-B-mediated signaling events has not been clarified in breast cancer cells. Regulation of apoptosis and cell cycle activities are linked to the mechanism of antiproliferation approaches exhibited by several natural compounds. These results revealed that paeonol has an anticancer and apoptotic effect that significantly suppressed MDA-MB-231 cell proliferation. Paeonol-treated cells showed classic morphological characteristics indicative of apoptosis such as chromatin condensation, fragmented nuclei, decreased cell size, cytoplasmic shrinkage, apoptotic bodies, and high stain to Annexin V apoptotic marker. Furthermore, before the G1 phase, the internucleosomal degradation of DNA produced an apoptotic peak (Sub-G1 phase), which attests that the suppressive effect was through induction of cell apoptosis. The G1 arrest prevents the reproduction of genetic lesions to offspring cells by allowing damaged DNA to be repaired. 27
Apoptosis is a kind of programmed cell death involved in developmental and degenerative stages by which organisms get rid of harmful cells that are dangerous to their survival. Due to different kinds of drugs and cell types, the apoptotic mechanisms induced by drugs are also complex. 28 Several studies have revealed that paeonol promotes apoptosis in esophageal cancer cell lines and human gastric cancer cell lines by suppressing Bcl-2 expression and modulating caspase-3 events, and induced cell death related to apoptosis by modulation expression of Bcl-2 and Bax in human colon cancer cell lines and mouse HepA-hepatoma. 29 –32 The authors previously demonstrated that paeonol exhibits anticancer effect by downregulating Bcl-2 and upregulating Bax, caspase 8, and caspase 3 via induction of apoptosis. 14
To know the apoptotic mechanism, they examined the CXCL4/CXCR3-B-mediated signals that play a vital role in the modulation of apoptosis. The induction of CXCR3-B via CXCL4 promotes p38 MAPK activation and inactivates ERK1/2; this promotes Nrf2 export from the nucleus, localizes BACH1 to the nucleus, and decreases HO-1 to reduce breast cancer cell proliferation via increased apoptosis. 33 In this report, paeonol-induced apoptosis activated CXCL4/CXCR3-B-mediated signals as there was an increased CXCR3-B and decreased HO-1 antiapoptotic molecule to promote apoptosis. Moreover, it was correlated with an upregulated BACH1 and decreased Nrf2. The transcription of HO-1 is strongly regulated by negative and positive regulator of BACH1 and Nrf2. 34 –36 Tumor cells express a variety of chemokines and receptors that are associated with disease progression and poor prognosis. 37 –42 Generally, the activation of CXCR3-A enhances tumor cell growth, whereas CXCR3-B inhibits tumor cell proliferation. Report shows that tumor tissues express low CXCR3-B, 10 and breast cancer CXCR3-B is inhibited when the Ras-Raf pathway is activated. 37 In prostate cancer, abnormal CXCR3-A expression and decreased CXCR3-B promote tumor invasion and metastasis by reversing the inhibitory function to a proliferative one. 43 Tumor cell proliferation may benefit from reduced CXCR3-B-mediated weak signaling events 37 and the upregulation of HO-1 antiapoptotic molecule. 13,44 Moreover, there is also evidence that HO-1 inhibition decreases survival of acute myeloid leukemia and colon carcinoma and promotes apoptosis of renal cancer cells, while HO-1 activation favors proliferation of several cancers, such as melanoma, hepatoma, lung, and thyroid carcinoma. 44,45
In this study, after administration of paeonol, the upregulated CXCR3-B suppressed the transcription of HO-1 via the moderation of BACH1 and Nrf2 and promoted apoptosis to inhibit breast cancer cell growth. This may have occurred as a result of the apoptosis processes induced by the drug and might better the prognosis of breast cancer patients. The results are consistent with a study on CXCR3-B overexpression plasmid, which lead to decreased HO-1 via the regulation of BACH1 and Nrf2. 46 Several anticancer agents may induce tumor cell apoptosis, and the induction of apoptosis in tumor cells is normally approved as a valid measure for effective cancer therapeutic agent. Moreover, the activators of Nrf2 are more favorable for suppressing cancers, while Nrf2 inhibitors are valuable therapeutic drugs. 47
In this report, paeonol-activated CXCL4/CXCR3-B-mediated signals promoted proliferation suppression via induced apoptosis on breast cancer cells. The anticancer mechanism might be associated with the G0/G1 cell arrest, upregulation of CXCR3-B, with decreased HO-1 antiapoptotic molecule through the moderation of BACH1 and Nrf2 to promote apoptosis. In a body, this study has shown that paeonols' anticancer and apoptotic effect can induce CXCL4/CXCR3-B-mediated signals to promote growth inhibition and apoptosis of breast cancer cells. It is associated with increased BACH1 and decreased Nrf2 to downregulate HO-1. Therefore, the authors suggest that paeonol has a significant chemotherapeutic effect on breast cancer.
Footnotes
Acknowledgment
This research was funded by the Natural Scientific Foundation of HeiLongJiang province under grant number [H2016089].
Disclosure Statement
There are no existing financial conflicts.