
Abstract
MiR-30d-5p, a member of the microRNA family, was recently reported to regulate androgen receptor signaling in prostate cancer (PCa). Ecto-5′-nucleotidase (NT5E/CD73) is a pivotal regulator of tumor migration and has angiogenetic properties. However, the undiscovered function of miR-30d-5p and whether it targeted NT5E in PCa remain uncertain. In this study, the authors observed miR-30d-5p was significantly downregulated in PCa tissues and cell lines compared with the adjacent normal tissues and normal prostate cells, respectively. The lower expression of miR-30d-5p was found to be inversely correlated with the NT5E expression in PCa tissues. Subsequently, the biological function of miR-30d-5p was evaluated in PCa in vitro. The results indicated that miR-30d-5p overexpression inhibited PCa cell growth and invasion by MTT, Transwell assays, respectively, as well as induced cell cycle G0/G1 phase arrest and apoptosis using flow cytometry analysis. In addition, miR-30d-5p directly bound to the 3′UTR (3′ untranslated region) of NT5E in DU-145 and PC-3 cells by luciferase reporter assay. Furthermore, enforced NT5E expression alleviated miR-30d-5p inhibition of PCa cell growth and invasion in DU145 cells. Taken together, these data indicated that miR-30d-5p may be a potential therapeutic target for the treatment of PCa by serving as a tumor suppressor, by negatively regulating NT5E.
Introduction
Being an extremely heterogeneous disease, prostate cancer (PCa) is the second most frequently diagnosed cancer in men and ranked as the fourth most common malignancy in the world. 1 Metastasizing to bone and soft tissues is considered a major deleterious event in patients with PCa. 2 A recent study estimated that 180,890 men were diagnosed with PCa and 26,120 patients succumbed to PCa in 2016 in the United States. 3 Initially, the cancer is androgen dependent and exposed to androgen deprivation therapy. 4 However, most of patients eventually turn into castrate-resistant PCa and become insensitive to further androgen deprivation. 5 Currently, it still remains largely unclear how PCa acquires proliferative ability and becomes highly metastatic upon androgen deprivation. Hence, exploring the molecular mechanism underlying the development and progression of PCa will provide propitious to develop novel gene therapeutic agents and contribute to identification of diagnostic markers.
As the most abundant family of small single-stranded noncoding RNA gene products, microRNAs (miRNAs) play roles in the maintenance of cellular machinery in steady state. 6 These miRNAs were found to be critical to attenuate the stability and translation of mRNAs by base pairing to partially complementary sites in the 3′UTR (3′ untranslated region) of their target genes, and involved in multiple fundamental physiological and pathological procedures. 7,8 In recent years, dysregulation of miRNAs has been shown to regulate tumor growth, metastasis, diagnosis, and prognosis. 8 For example, the comprehensive expression analysis of a large number of miRNAs could reflect the developmental lineage and differentiation state in human cancer. 6 The role of miR-21 to function as a tumor gene was confirmed in colorectal cancer, 9 gastric cancer, 10 and nonsmall cell lung cancer (NSCLC). 11 MiR-30d-5p, a member of the miRNA family, has been reported to regulate androgen receptor signaling in PCa. 12 The levels of miR-30d-5p were also found to be negatively associated with the activity of androgen receptor in PCa. 12 Moreover, downregulation of miR-30d-5p could abrogate tumor growth and motility by targeting cyclin E2 in NSCLC. 13
The Ecto-5′-nucleotidase (NT5E/CD73) mRNA encodes a membrane-bound protein that involved in the catalytic process to hydrolyze extracellular nucleotides to bioactive nucleoside intermediates. 14 In addition, NT5E is a pivotal regulator in tumor migration and angiogenetic properties, and involved in depressing anticancer immune responses by activating adenosine receptors. 15,16 Mechanism studies have revealed that NT5E is a direct target of two miR-30 family members (miR-30a-5p and miR-30b) in NSCLC and gallbladder carcinoma (GC), respectively. 16,17 Interestingly, the miRNA-mRNA interaction accounts for miR-30a-5p- and miR-30b-inhibited growth and carcinogenesis of NSCLC and GB tumor cells. 16,17 It also seems that another member of the miR-30 family, miR-30d-5p, might present similar behavior by directly binding to NT5E and plays an important role in the pathogenesis of PCa.
To validate their hypothesis, the authors first determined the expression levels of miR-30d-5p and NT5E in PCa tissues and cell lines and then investigated the biological effects of miR-30d-5p in PCa in vitro using a series of functional assay. In addition, bioinformatics analysis and dual-luciferase reporter assay were used to reveal the underlying mechanisms by which miR-30d-5p regulates PCa. These findings will provide a novel biomarker for therapeutic strategies, diagnosis and prognosis for PCa.
Materials and Methods
Patients and tissue samples
In total, 40 pairs of tumor tissues and adjacent normal tissues were obtained from PCa patients during surgery at Renmin Hospital of Wuhan University (Hubei, China). All obtained samples were immediately frozen in liquid nitrogen and kept at −80°C before RNA extraction. All the patients had not received any radiotherapy or chemotherapy before surgery. The clinicopathological features of prostate tumors of the patients were evaluated histologically by independent pathologists according to the criteria of EAU guidelines 18 and summarized in Table 1. Informed consent was obtained from the included patients. All procedures performed in this study were approved by the Ethics Committee of Renmin Hospital of Wuhan University.
PSA, prostate specific antigen.
Cell culture and transfection
Human PC cell lines (LNCaP, VCaP, DU145, and PC3) and a normal prostate epithelium cell line RWPE were purchased from ATCC (Manassas, VA). All cell lines were cultured in RMPI-1640 (Invitrogen) supplemented with 10% of fetal bovine serum (Invitrogen), 100 units/mL penicillin (Invitrogen), and 100 g/mL streptomycin (Invitrogen), which were all maintained in a humidified atmosphere of 5% CO2, at 37°C.
The synthesized oligos for miR-30d-5p mimics (miR-30d-5p) and a corresponding negative control (NC mimics) were obtained from GenePharma (Shanghai, China). The NT5E overexpression plasmid and the corresponding control were purchased from Genecopoeia (Guangzhou, China). For cell transfection, DU145 and LNCaP were seeded in six-well plates and the transfection was performed using Lipofectamine™ 2000 (Invitrogen) according to the instructions provided by the manufacturer. After 48 h of transfection, cells were harvested for the following assays.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was extracted from PCa clinical tissue samples or cell lines using TRIzol reagent (Invitrogen) following the manufacturer's protocol. For miR-30d-5p detection, cDNA synthesis and subsequent quantification were performed using predesigned TaqMan assays (Applied Biosystems, Foster City, CA). Small nuclear RNA U6 was used as an endogenous control. For NT5E gene expression detection, cDNA synthesis was conducted using a PrimeScript RT Reagent Kit (Takara Biotechnology Co., Ltd.) and quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using a 7500 sequence detection system (Applied Biosystems), and normalized to GAPDH (glyceraldehyde 3-phophate dehydrogenase) expression. The relative expression levels of miRNA and mRNA were analyzed with the 2−ΔΔCt method.
Western blotting
The total proteins were extracted from PCa cells using a lysis buffer (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer's protocol and quantified by BCA assay (Beyotime, Shanghai, China). Equal amounts of protein were separated by 10% sodium dodecyl sulfate–polyacrylamide gels (SDS-PAGE) and transferred onto PVDF (polyvinylidene fluoride) membranes (Millipore). The membranes were blocked with 5% nonfat dry milk for 1 h and then probed with primary antibodies against NT5E (1:500; Proteintech) and GAPDH (1:5000; Cell signaling Technology) overnight at 4°C, followed by incubation with peroxidase-conjugated secondary antibodies for 2 h at room temperature. The bands were visualized by an enhanced chemiluminescence kit (Beyotime). GAPDH was used as an internal control.
Cell viability assay
Cell viability of PCa cells was characterized by Cell Counting Kit-8 (CCK-8) (Dojindo, Kumamoto, Japan) following the manufacturer's instructions. DU145 and LNCap cells were equally plated in 96-well plates (2500 cells/well) 48 h after transfection and maintained for 5 d. At an interval of every 24 h, 10 μL of CCK-8 was added to the plates. After 4 h of incubation, the optical density was detected at a wavelength of 490 nm on SpectraMax 250 microplate reader (Molecular Devices). Each experiment was performed in triplicate.
Colony formation assay
Cell proliferation was evaluated based on colony numbers using the colony formation assay. In brief, DU145 and LNCaP cells were seeded in six-well plates at a density of 200 cells per well and cultured for 7 d. Then cells were fixed with methanol for 30 min, followed by staining with 0.1% crystal violet (Beyotime) for 10 min. After being washed with water and air-dried, the number of colonies (containing >50 cells/colony) were counted under a light microscope. Triplicate plates were determined in each group.
Flow cytometry assay
Cell cycle distribution of PCa cells was analyzed by flow cytometry with propidium iodide (PI) staining. Briefly, cells (1 × 105 cells/dish) were seeded on 6-cm dishes and cultured for 3 d. The cells were then fixed by precooled 70% ethanol for 30 min at 4°C and stained with a PI solution (100 μg/mL) containing 10 μg/mL of DNase-free RNase, followed by flow cytometer analysis (FACSCalibur; BD Biosciences). Similarly, apoptosis of PCa cells was detected by Annexin V-APC/7-AAD apoptosis detection kit according to the manufacturer's instructions (KeyGEN Biotech, Nanjing, China).
Cell migration assay
Cell migration was assessed using a wound-healing assay in DU145 and LNCaP cells. Briefly, cells were seeded in six-well plates and transfected with miR-30d-5p or NC mimics, or co-transfected with miR-30d-5p and NT5E plasmid. After 48 h of transfection, a wound field was made using a sterile 200 μL pipette tip among cells in each well. The mobilized cells were observed by a Leica DMI 6000B microscope (Leica Microsystems, Inc., IL) at 0 and 48 h, respectively. The relative migration ability was evaluated by determining the width of wound healing.
Bioinformatics analysis and dual-luciferase reporter assay
The potential target genes of miR-30d-5p were found by using the online software programs TargetScan (
Statistical analysis
All statistical analyses were performed using GraphPad Prism 6.0 software (GraphPad Software, Inc., La Jolla, CA). All quantitative data were expressed as mean ± standard deviation of at least three experiments. The correlation between miR-30d-5p and NT5E expression in PCa tissues was determined using Pearson's correlation coefficient by GraphPad Prism 6.0 software. The significance of differences among the groups was estimated by Student's t-test or one-way ANOVA. Values of p < 0.05 were considered statistically significant.
Results
Expression of miR-30d-5p and NT5E in PCa tissues
qRT-PCR assay was first used to determine the expression of miR-30d-5p and NT5E in 40 paired clinical PCa and adjacent normal tissues. As shown in Figure 1A, the expression of miR-30d-5p was significantly downregulated in PCa tissues compared with matched normal tissues (p < 0.001). On the contrary, PCa samples exhibited higher levels of NT5E mRNA compared to the paired nontumor tissues (p < 0.001, Fig. 1B). Statistical analysis further demonstrated that the expression of miR-30d-5p was negatively correlated with NT5E expression in PCa tissues (p = 0.0206, Fig. 1C). These results indicated that miR-30d-5p and NT5E may serve an important role in PCa.
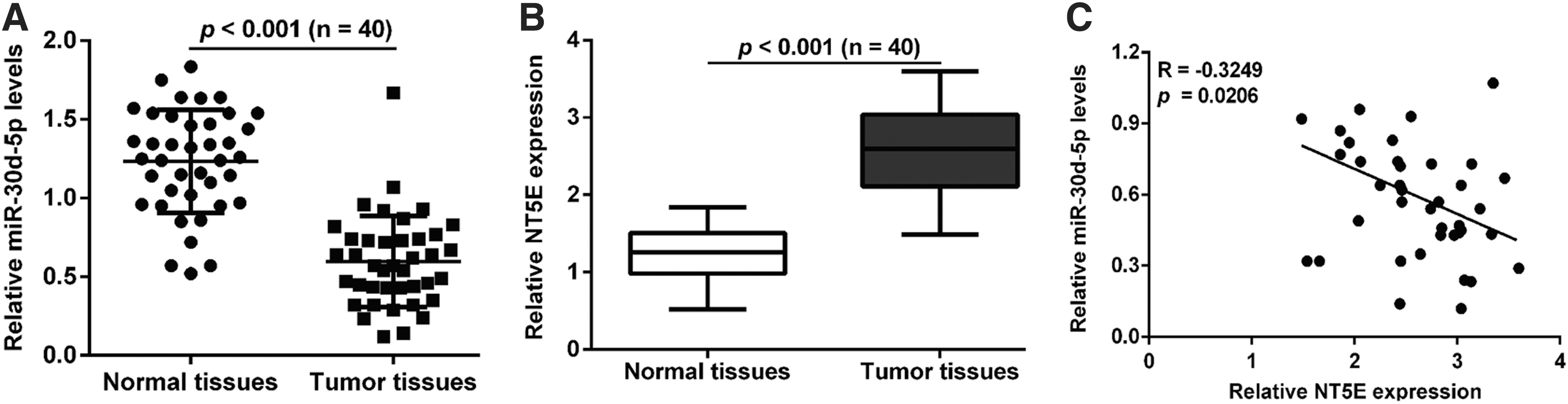
MiR-30d-5p and NT5E were aberrantly expressed in PCa tissues compared with adjacent normal tissues. qRT-PCR was performed to determine the expression of
Inhibitory effects of miR-30d-5p on cell viability and proliferation in PCa
Next, the authors used qRT-PCR to measure the endogenous miR-30d-5p in PCa cell lines, including prostate specific antigen (PSA)-negative DU145 and PC3, as well as PSA-positive LNCaP and VCaP cells. As shown in Figure 2A, miR-30d-5p was significantly downregulated in all examined PCa cell lines, compared to that in noncarcinoma human prostate cell line, RWPE (p < 0.01). Notably, PSA-positive LNCaP and PSA-negative DU145 presented lower miR-30d-5p expression compared with VCaP and PC3, respectively, and thus were selected for overexpressing miR-30d-5p through the transfection of miR-30d-5p mimics. After 48 h of transfection, qRT-PCR further confirmed that miR-30d-5p mimic transfection remarkably upregulated miR-30d-5p levels in both LNCaP and DU145 cells (p < 0.001, Fig. 2B). Ectopic forced expression of miR-30d-5p resulted in dramatic suppression of cell viability and proliferation capacity of PCa cells compared with NC mimic-transfected cells using CCK-8 assay (p < 0.01, p < 0.001, Fig. 2C, D) and colony formation assay (p < 0.01, Fig. 2E), respectively.

MiR-30d-5p suppressed cell viability and proliferation in PCa cells.
Effects of miR-30d-5p on cell cycle distribution and apoptosis in PCa
To examine the mechanism underlying the inhibition of cell growth, the cell cycle distribution and apoptosis were detected in PCa cells after transfection with miR-30d-5p mimics or NC mimics using flow cytometry analysis. As shown in Figure 3A, upregulation of miR-30d-5p markedly increased the percentage of cells in G0/G1 phase (from 54.30% ± 1.00% to 66.33% ± 1.99%, p < 0.01), and decreased the percentage of cells in S (from 28.67% ± 1.03% to 18.89% ± 0.71%, p < 0.01) and G2/M phase (from 17.03% ± 0.59% to 14.77% ± 1.33%, p < 0.05) in LNCap cells. Similarly, the percentage of cells in G0/G1 phase (51.17% ± 1.17% vs. 65.62% ± 0.62%, p < 0.001) was significantly elevated, and was reduced in S (28.74% ± 0.51% vs. 18.95% ± 0.25%, p < 0.01) and G2/M phase (20.09% ± 0.99% vs. 15.43% ± 0.64%, p < 0.01) in the miR-30d-5p group compared with the NC mimics' group in DU145 cells. These results indicated that the upregulation of miR-30d-5p induced cell cycle G0/G1 phase arrest in PCa cells. In addition, the authors analyzed the effect of miR-30d-5p overexpression on apoptosis in PCa cells. As depicted in Figure 3B, the upregulation of miR-30d-5p significantly promoted cell apoptosis in both LNCaP (p < 0.01) and DU145 (p < 0.05) cells.
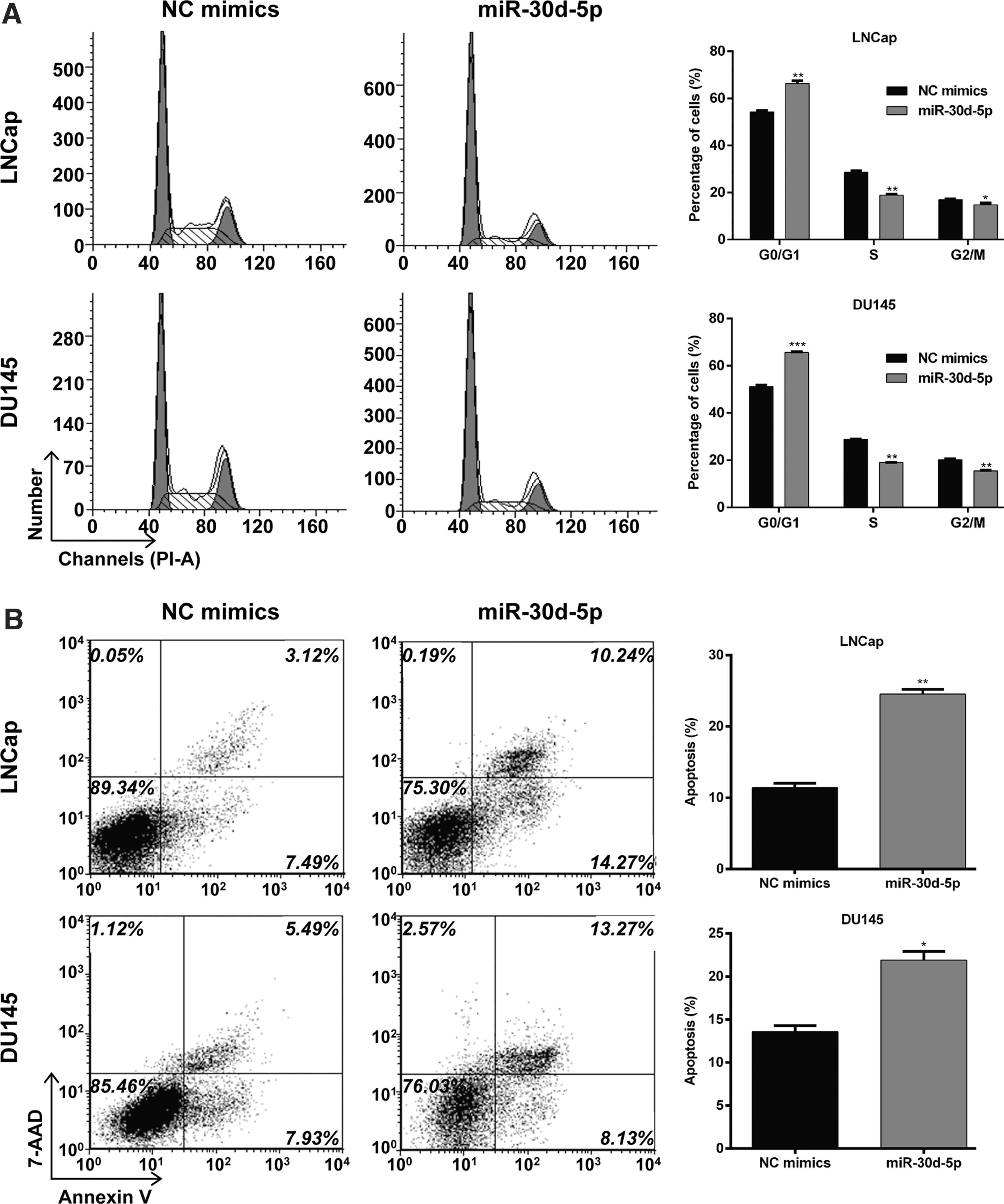
MiR-30d-5p induced cell cycle G0/G1 phase arrest and apoptosis in PCa cells.
Upregulation of miR-30b-5p suppressed PCa cell migration ability
Furthermore, the potential role of miR-20b on the migration ability of PCa was evaluated by performing the wound-healing assay. As shown in Figure 4A and B, continuous rapider movement was observed in NC mimic-transfected LNCaP and DU145 cells compared with miR-30d-5p mimic transfection. Quantitative analysis further confirmed that the wound area in the miR-30d-5p group was significantly decreased compared to the NC mimics' group in LNCaP (p < 0.001) and DU145 (p < 0.001) cells. These results suggested that miR-30d-5p upregulation resulted in significant suppression in cell migration ability in PCa cells.
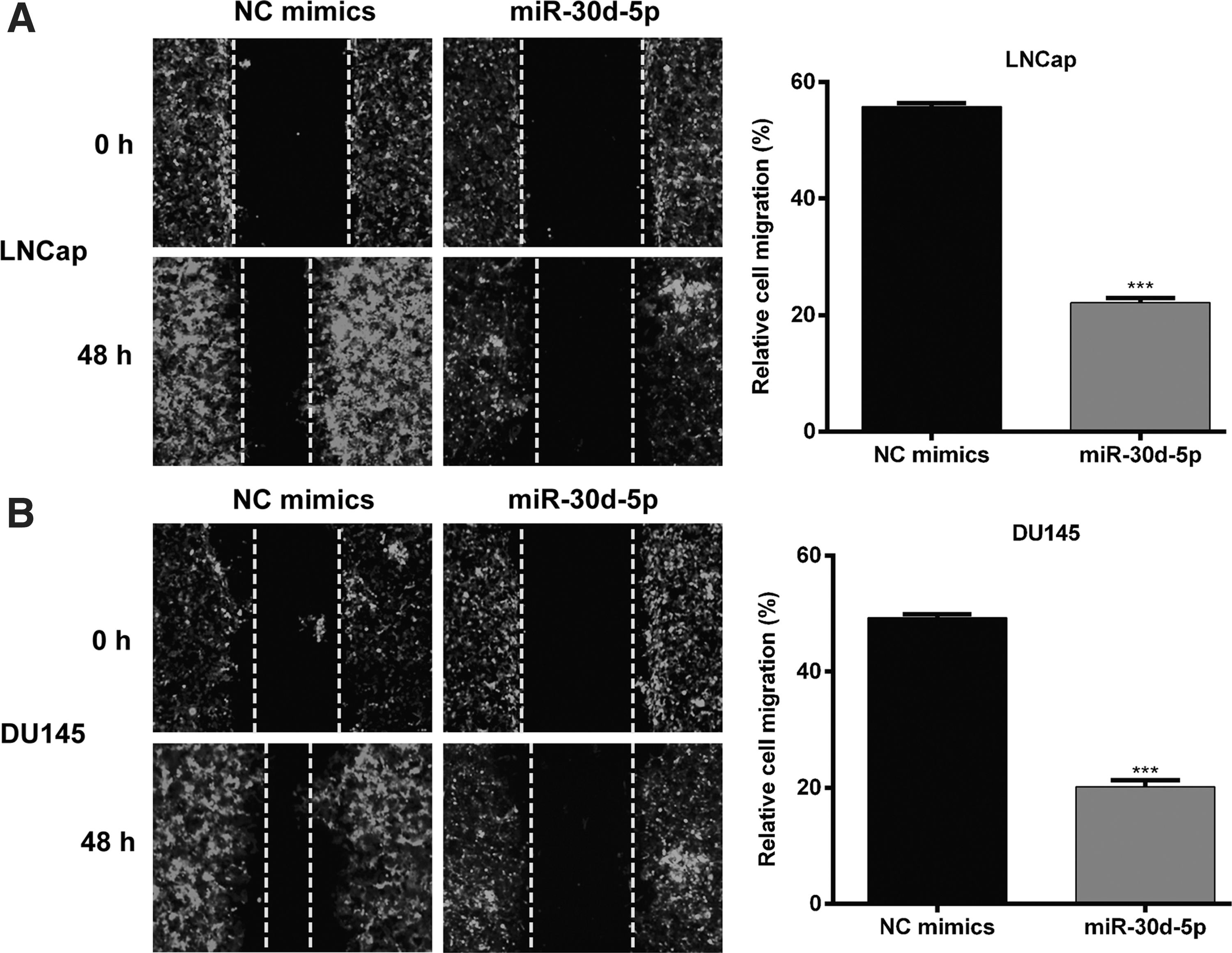
MiR-30d-5p suppressed cell migration ability in PCa cells.
NT5E is a direct target gene of miR-30b-5p in PCa cells
Through publicly available databases search, the 3′UTR of NT5E was predicted to have a complementary sequence with miR-30d-5p (Fig. 5A). To validate this prediction, the NT5E WT and NT5E MUT containing miR-30d-5p matching sequence were constructed and dual-luciferase reporter assay was performed in LNCaP and DU145 cells. As shown in Figure 5B, miR-30d-5p significantly reduced the luciferase activity of NT5E WT, but did not reduce the activity of NT5E MUT in LNCaP (p < 0.01) and DU145 (p < 0.05) cells. In addition, the authors examined whether miR-30d-5p inhibits endogenous NT5E expression in LNCaP and DU145 cells using Western blotting. Compared with the NC mimic transfection, endogenous NT5E protein expression levels were significantly downregulated in LNCaP and DU145 cells transfected with miR-30d-5p (Fig. 5C), which further confirmed the inverse correlation between miR-30d-5p and NT5E. Collectively, the results provide further evidence supporting the hypothesis that NT5E is a target of miR-30d-5p in PCa.
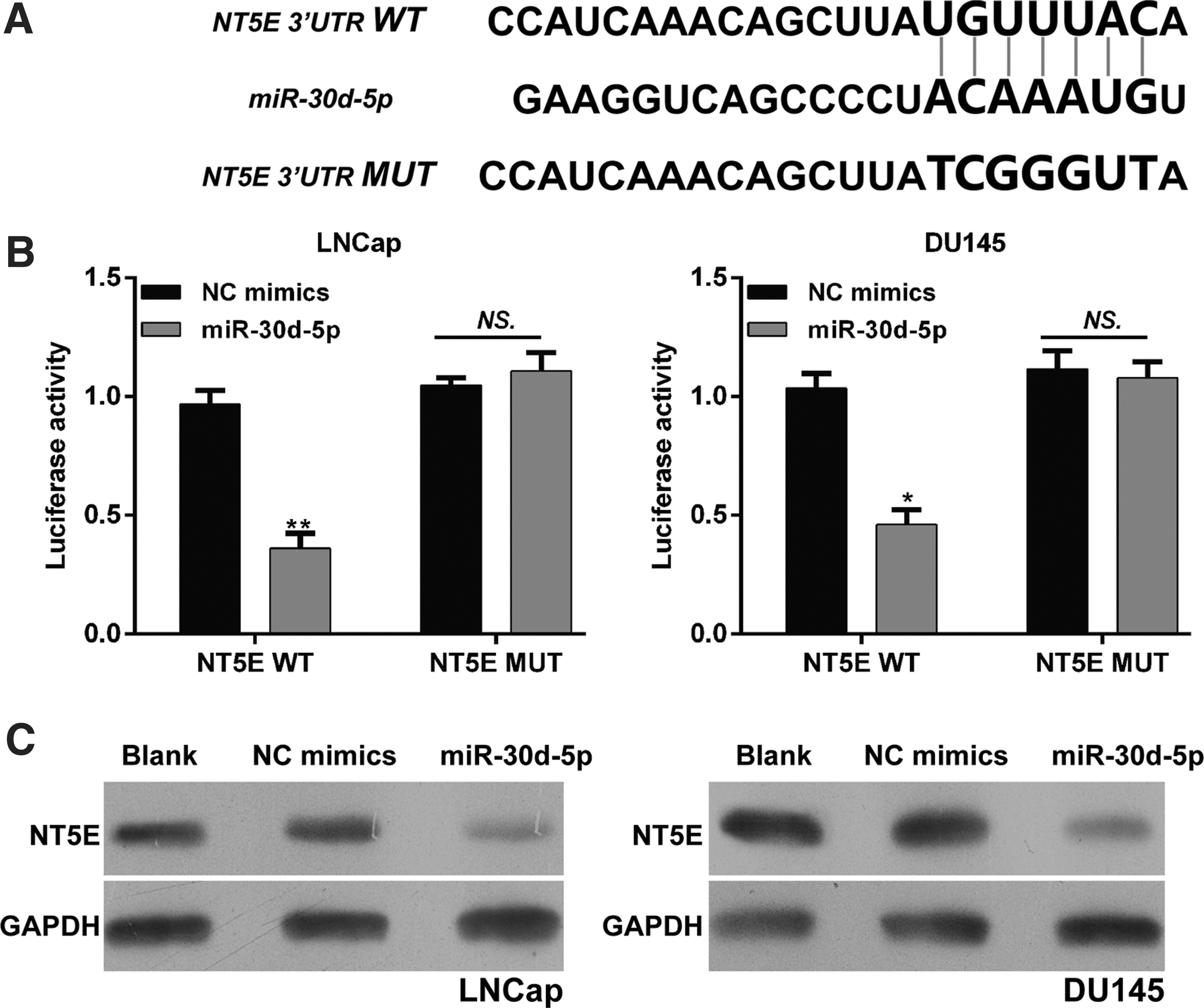
NT5E is a target of miR-30d-5p in PCa cells.
Enhanced expression of NT5E could partially rescue the inhibition effect of miR-30b-5p on PCa cells
The above data suggested that NT5E was upregulated in PCa and inversely correlated with miR-30d-5p expression. Therefore, the authors investigated whether NT5E mediated the inhibitory effect of miR-30d-5p on cell proliferation and migration by performing rescue experiments in DU145 cells. As shown in Figure 6A, Western blot confirmed the upregulated expression of NT5E after co-transfection with miR-30d-5p and NT5E compared with solely miR-30d-5p transfection. In vitro functional assays indicated that overexpression of NT5E partially rescued the impaired cell proliferation (p < 0.001, Fig. 6B) and migration ability (p < 0.05, Fig. 6C) in DU145 cells. These data demonstrated that NT5E indeed partially reversed the effects of miR-30d-5p in PCa cells.
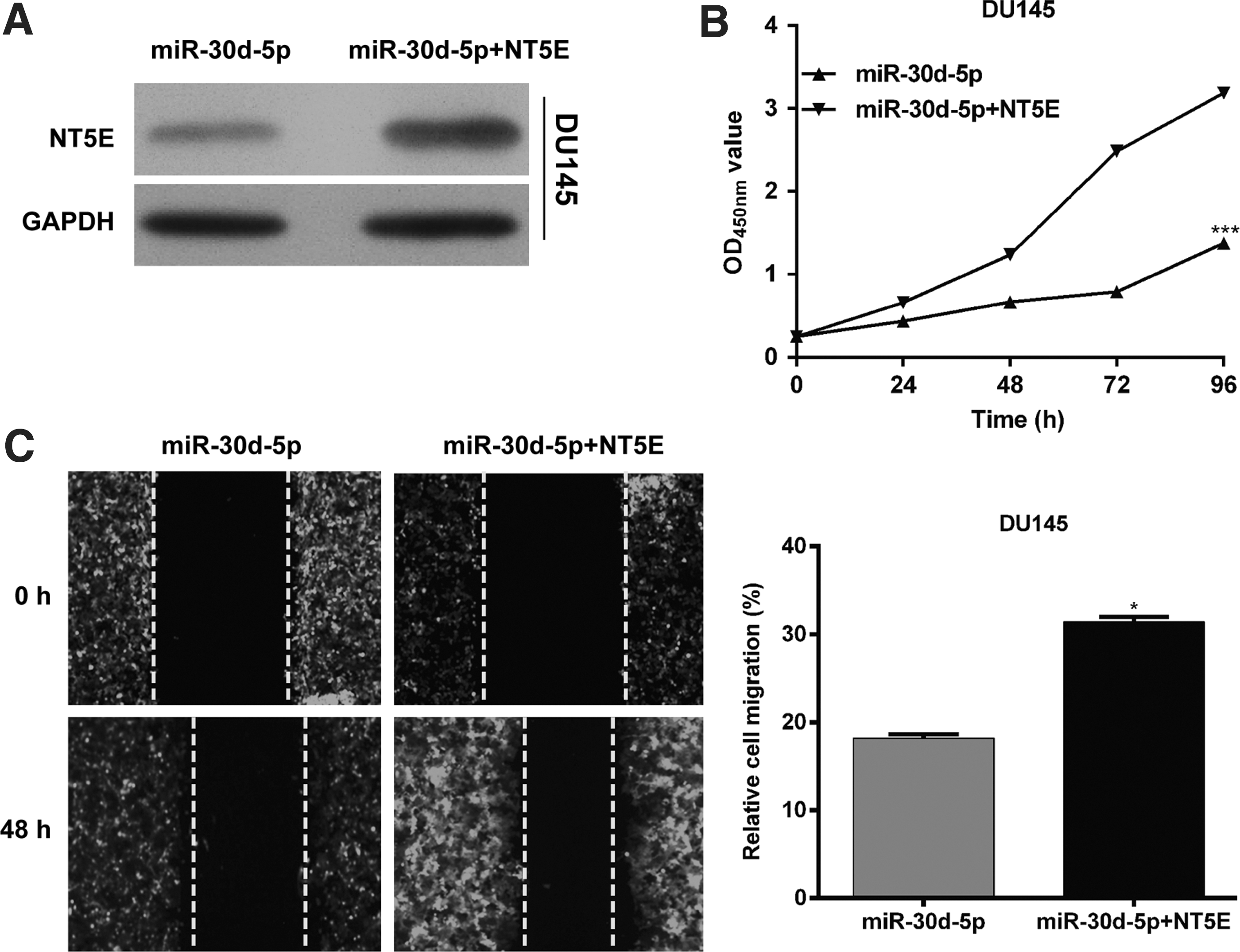
Overexpression of NT5E partially rescues the inhibitory effects of miR-30d-5p on cell proliferation and migration in PCa. DU145 cells were transfected with miR-30d-5p together with plasmid expressing NT5E or empty vector.
Discussion
A growing body of evidence revealed that miRNAs are associated with multiple biological processes and are frequently dysregulated in various human cancers. 20 In this study, reduced miR-30d-5p was found in PCa specimens and cell lines in comparison with adjacent nontumor prostate tissues or noncarcinoma human prostate cell line, RWPE. Ectopic expression of miR-30d-5p lead to decreased cell viability, proliferation, and migration, along with increased apoptosis and G0/G1 phase arrest, whereas NT5E had reversed antitumor effects in vitro. At both the transcriptional and translational levels, the expression of miR-30d-5p was observed to be negatively correlated with NT5E expression.
Chen et al. 13 showed that miR-30d-5p was downregulated in tumor tissues and acted as tumor suppressor by regulating the proliferation, migration, and invasion of NSCLC cells. 13 Therefore, these results are consistent with “carcinomatosis” found in Chen et al., 13 and the loss of miR-30d-5p expression in PCa may provide an unbiased disease biomarker differentiating PCa from health.
With regard to the function of miR-30d-5p, its target gene has also been revealed in NSCLC. Chen et al. 13 identified CCNE2 as the direct target of miR-30d-5p and responsible for the antitumorigenesis mediated by miR-30d-5p in NSCLC. This study identified and characterized a novel gene, NT5E, as a direct target for miR-30d-5p. As a result, it appears that miR-30d-5p in humans might have multiple targets that exert diverse roles under different pathological conditions.
The NT5E gene Ecto-5′-nucleotidase (CD73), a 71 kD dimetric enzyme encoded by NT5E gene could hydrolyze extracellular ADP into phosphorylate and adenosine, which initiates adenosine receptor-dependent signaling through four subtypes, including G protein-coupled adenosine receptors. 21,22 NT5E has putative roles in a number of cellular processes, including proliferation, survival, and adhesion. 23,24 Activation of the adenosine receptor-dependent signaling could contribute to immune escape in malignant cells that express high levels of CD73. 25,26 Besides, the signaling and adhesive properties of CD73 are independent in the immune responses, which are associated with tumorigenesis, angiogenesis, and migration. 27 –29 Zhi et al. 30 showed that CD73 silencing promotes breast cancer cell apoptosis and causes G0/G1 cell cycle arrest. Bonnin et al. 31 revealed that reduced expression of miR-422a accelerates proliferation and cellular adhesion of head and neck squamous cell carcinoma cells by targeting CD73. This study demonstrated that the expression of NT5E was downregulated in LNCaP and DU145 cells with the overexpression of miR-30d-5p. Enhanced expression of NT5E partially reversed the inhibitory effects of miR-30d-5p overexpression. Therefore, the authors conclude that the decrease in endogenous level of miR-30d-5p could promote tumor immune escape and triggers adenosine receptor-dependent signaling and CD73 adhesive property by the upregulation of NT5E in PCa, therefore contributing to tumor cell proliferation and migration.
In summary, this study demonstrated that the expression levels of miR-30d-5p were significantly downregulated in PCa tissues and cell lines. Overexpression of miR-30d-5p significantly inhibited PCa cell proliferation and migration by targeting NT5E. To the best of the authors' knowledge, this is the first study describing that the tumor suppressor miR-30d-5p directly regulated NT5E in PCa cells. All the findings show that miR-30d-5p might be a potential therapeutic target for PCa diagnosis and treatment.
Footnotes
Disclosure Statement
No competing financial interests exist.