
Abstract
Objective:
Long non-coding RNA-ATB (Lnc-ATB) have been reported to promote tumor proliferation and metastasis via regulation of tumor suppressive miRNA-related signals. Patients with endometrial cancer (EC) have advanced stage disease or metastasis have poor prognosis. We here investigated the role of Lnc-ATB in endometrial cancer.
Methods:
Endometrial cancer tissues and normal tissues (n = 35) were collected to determine the expression and clinical significance of Lnc-ATB, and bioinformatics analysis was used to predict the miRNA target. siRNA was used to estimate the function of Lnc-ATB in EC cell lines and in vivo.
Result:
The expression of Lnc-ATB is up-regulated in tumor tissues and EC cell lines. Patients with high expressed Lnc-ATB have high FIGO stage and poor tumor differentiation. The tumor suppressor miR-126 interacted with Lnc-ATB. Down-regulated miR-126 negative correlated with FIGO stage and tumor differentiation. Knockdown of Lnc-ATB in RL95 and HEC1A cell lines increased the miR-126 level and impaired the cell vitality, induced caspase-3-related tumor apoptosis and G1/S arrest. However, abrogation of miR-126 by its inhibitors counteracted Lnc-ATB knockdown-induced tumor inhibition via regulation of miR-126 target gene PIK3R2 and Sox2-related apoptosis and cell cycle pathway. Meanwhile, Lnc-ATB knockdown also suppressed the migration and invasion and inhibited TGF-β-induced epithelial-mesenchymal transition (EMT) phenotype via miR-126. Knockdown of Lnc-ATB in vivo remarkably induced tumor regression via restoration of tumor suppressor miR-126, leading to deceased tumor volume, reduced expression of PCNA and PIK3R2/Sox2 signals and EMT phenotype in tumor tissues.
Conclusion:
These data demonstrate the tumorigenic role of Lnc-ATBs in endometrial cancer via abrogation of tumor suppressor miR-126 signals.
Introduction
Endometrial cancer (EC) is the most common gynecological malignancy of the female genital tract, the annual incidence of which has been estimated to be 10–20 per 100,000 women and also still increased worldwide. 1 Based on clinicopathological and molecular genetic features, EC can be divided into two major categories: type I (estrogen-dependent) and type II (estrogen-nondependent). The most dominant subtype (namely type I) endometrioid endometrial cancer (EEC) accounts for ∼80% of the cases with favorable prognosis. However, patients with advanced stage or non-EEC ECs (namely type II) tend to display more aggressive characteristics, including local recurrence and distal metastases with poor prognosis. 2,3 Therefore, the identification of effective targets for EC tumorigenesis and treatment would be beneficial for improving the survival time of EC patients.
The human transcriptome consists of protein-coding genes and noncoding RNAs (ncRNAs). ncRNA is an RNA molecule with no potential to translate into a protein and evolutionarily conserved among organisms. 4 According to their length, these regulatory ncRNAs can be further divided into short ncRNAs, including small interfering RNAs (siRNAs), microRNAs (miRNAs), PIWI-interacting RNAs, and long noncoding RNAs (lncRNAs). 5,6 The most investigated ncRNAs is miRNAs and LncRNAs, which were found to be involved in transcriptional and post-transcriptional regulation of gene expression in many diseases, especially in cancer. 4,7 The miR-17-92 cluster could code for six different miRNAs often upregulated in tumor tissues and functioned as oncogene through repressing c-Myc, PI3K, and NF-κB pathways in lymphoma, multiple myeloma, pancreatic cancer, and so on. 8 –10 Some miRNAs, including miR-34a, let-7, miR-98, and miR-214, were downregulated in certain cancer types with tumor-suppressive role. 11,12 Similarly, LncRNA-MALAT1, MEG3, H19, HULC, and so on have been found to participate into tumor proliferation, resistance to apoptosis, and cancer therapy through interaction with miRNAs and their target pathways. 13 For example, in EC, the level of H19 expression was also strongly correlated with tumor invasion of the reproductive organs and significantly correlated with neoplastic cell invasion of the myometrium. 14 lncRNA-TUG1 expression in EC tissues was significantly higher than that in adjacent tissues and could promote the progression of EC through inhibiting miR-299 and miR-34a-5p. 15 The role of LncRNA/miRNA signal axis in EC remains to be further investigated.
The LncRNA-activated by TGF-β (Lnc-ATB) was first found to be upregulated in hepatocellular carcinoma (HCC). 16 Lnc-ATB as a mediator of TGF-β signaling upregulated ZEB1 and ZEB2 by competitively binding the miR-200 family and then induced EMT and invasion in HCC patients. Currently, Lnc-ATB have been reported to be involved in many cancers, including colorectal cancer, 17 glioma, 18 osteosarcoma, 19 and gastric cancer, 20 through regulation of cell proliferation, apoptosis, cell cycle, and metastasis. However, the clinical significance and function of Lnc-ATB EC is unclear.
In this study, it was found that the expression of Lnc-ATB is upregulated during the development of EC. Interestingly, Bioinformatics analysis predicted that a classic tumor suppressor miR-126 could bind to Lnc-ATB. Cell growth and metastasis-related PIK3R2/PTEN/PI3K signaling and KRAS/ERK signaling were the target pathways of tumor suppressor miR-126. 21,22 Since EC is highly mutated in regard to PTEN, KRAS, and PI3K pathways, 23 this study hypothesized that Lnc-ATB/miR-126 signals participated into the progression of EC.
Materials and Methods
EC samples
Ethical approval for the study was obtained from the Fifth Hospital of Xi'an Ethics Committee and all participants provided their written consent. To determine the expression of Lnc-ATB/miR-126, EC tissues were collected from women (age median = 58) diagnosed with EC (n = 35, 28 endometrioid carcinoma, 3 serous carcinoma, 3 clear cell carcinoma, and 1 undifferentiated carcinoma) from the Fifth Hospital of Xi'an and the histological types were conducted according to surgical staging International Federation of Gynecology and Obstetrics (FIGO). Normal specimens (n = 35) were collected from women undergoing hysterectomy for nonmalignant conditions. Histological typing of tumor was classified according to the World Health Organization criteria.
RNA isolation and quantitative real-time reverse transcription polymerase chain reaction
To estimate the expression of Lnc-ATB/miR-126 and the target genes of miR-126 in tumor tissues and cells, total RNA was extracted using Trizol reagent (Invitrogen) according to the standard RNA isolation protocol; A260/A280 nm >1.8 was qualified for quantitative analysis. Quantitative real-time reverse transcription polymerase chain reaction was performed, and the expression levels of genes were estimated through 2−ΔΔCt method and were normalized to GAPDH for gene expression.
Cell lines and reagents
EC cell lines RL95, HEC1A, AN3CA, and Ishakawa and the immortalized human endometrial epithelial cell line EM-ER-A were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (Life Technologies), ampicillin, and streptomycin at 37°C and 5% CO2 conditions. The antibodies for PIK3R2, Sox2, Bcl-2, caspase-3, CCND1, CDK4, E-cadherin, and N-cadherin were obtained from Cell Signaling Tech (Denver, MA) and Abcam. The siRNA for Lnc-ATB were purchased from RiboBio (Guangzhou, China). Negative control and oligonucleotide sequences of miR-126 mimics or miR-126 inhibitors were purchased from RiboBio. The renilla luciferase reporter vector psiCHECK2-Lnc-ATB-wild or mut were conducted by GenePharma (Shanghai, China).
Luciferase reporter assay
The RL95 cells were cotransfected containing negative control, psiCHECK2-Lnc-ATB-wild or mut, or miR-126 mimics. Luciferase activity was measured using the Dual-Luciferase Reporter Assay System (Promega). Firefly luciferase acted as a reporter gene for normalized control.
CCK-8 assay
The RL95 and HEC1A cell lines were transfected with siRNA-Lnc-ATB, miR-126, and negative control for indicated time. Then cells were harvested and washed with phosphate buffered saline (PBS) and then cell counting kit-8 (Kumamoto, Japan) mixed with DMEM was used for cell viability assay, and the absorbance was measured at 450 nm by a microplate reader.
Hoechst assay
RL95 and HEC1A cells were transfected with siRNA-Lnc-ATB, miR-126, and negative control for indicated time and were harvested and washed by PBS, and the cells were fixed by PFA. The cells were stained with 0.1 μg/mL Hoechst 33342 (Sigma, St Louis, MO); the changes in nuclear morphology were observed under a fluorescence microscope (Olympus, Tokyo, Japan).
Flow cytometry
To determine the cell cycle, cells were stained with propidium iodide (PI) staining solution (10 μg/mL RNase A and 50 μg/mL PI) at 37°C for 30 min in dark. The cell cycle distribution was also analyzed using a flow cytometry provided with the Cell-Quest software. To determine the cell apoptosis, 2 μL of annexin V mixed with 2 μL of PI (eBioscience) was added into each tube for 30 min according to the manufacturer's instructions and quantified by flow cytometry on a FACSCalibur instrument.
Western blot
At 48 h after treatment, cells were washed with cold PBS and subjected to a lysis buffer. Protein lysates were separated using 8% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis, then electrotransferred onto nitrocellulose filter membranes. The membranes were blocked with a buffer containing 5% nonfat milk in PBS with 0.05% Tween-20 for 2 h and incubated with primary antibody overnight. Then the membranes were incubated with peroxidase-conjugated secondary antibodies (Millipore, Darmstadt, Germany) and developed using an enhanced chemiluminescence detection kit (Pierce, Rockford, IL). β-Actin was used as blank control.
Transwell assay
RL95 and HEC1A cells were transfected with siRNA-Lnc-ATB, miR-126, and negative control for the indicated time. A total of 2 × 10 4 cells were in the upper chamber of a noncoated Transwell insert for the migration assay. For invasion assay, cells in growth medium without serum were seeded into the upper wells of a BD BioCoat Chambers (BD Science). The lower wells contained the same medium with 10% serum. After 24 h of incubation, the cells that had migrated to the lower side of the chamber were fixed with 2.5% glutaraldehyde, stained with 0.1% crystal violet and counted under microscopy.
Immunohistochemistry and immunofluorescence assays
Immunohistochemistry assay was performed to determine the expression of PCNA in tumor tissues and 2-μm-thick formalin-fixed and paraffin-embedded specimen sections were used. Immunofluorescence assays are performed to estimate the E-cadherin and N-cadherin levels in tumor tissues according to previous study described. 24
Tumor model
To investigate the role of Lnc-ATB in EC in vivo. RL95 cells were infected with lentivirus-RNAi-Lnc-ATB (multiplicity of infection = 100), miR-126 mimics or inhibitors or negative control. RL95 cells (2 × 10 6 cells) were subcutaneously coinjected in the rear flank of nude mice (5 per group). V (cm 3 ) = width2 (cm2) × length (cm)/2. Results are presented as the mean tumor size (mm 3 ) for every group at various time points until the end point of the experiment. The animal study was approved by institutional animal research committee of Fifth Hospital of Xi'an and that animals were cared for, following the guidelines for use and care of laboratory animals.
Statistical analyses
The Prism statistical software package (version 5.0; Graphpad Software, Inc.) was used in this study, and the statistical analyses were performed. Unpaired t-tests or Mann–Whitney U tests were used to compare the two groups, and multiple group comparisons were analyzed with one-way analysis of variance. p < 0.05 was considered statistically significant. All experiments were performed at least three times.
Results
Overexpressed Lnc-ATB predicted poor clinical outcomes for patients with EC
First the clinical expression pattern of Lnc-ATB in EC was investigated. This study collected EC tissues and normal tissues (n = 35) and measured the expression of Lnc-ATB. The results of quantitative real time polymerase chain reaction (Q-PCR) indicated that the Lnc-ATB level was significantly elevated in EC tissues when compared with normal tissues (Fig. 1A). Meanwhile, the clinical relevance of Lnc-ATB to clinicopathological characteristics of patients with EC was also analyzed. It was found that the expression of Lnc-ATB positively correlated with FIGO stage and patients with high stage have more expression of Lnc-ATB (Fig. 1B). The tumor differentiation is also associated with Lnc-ATB levels in tumor tissues. The poor differentiation of tumor cell has highly expressed Lnc-ATB (Fig. 1C). But the expression of Lnc-ATB showed to be comparable in patients with different ages (Fig. 1D). These data indicated that highly expressed Lnc-ATB in EC is associated with poor clinicopathological characteristics.
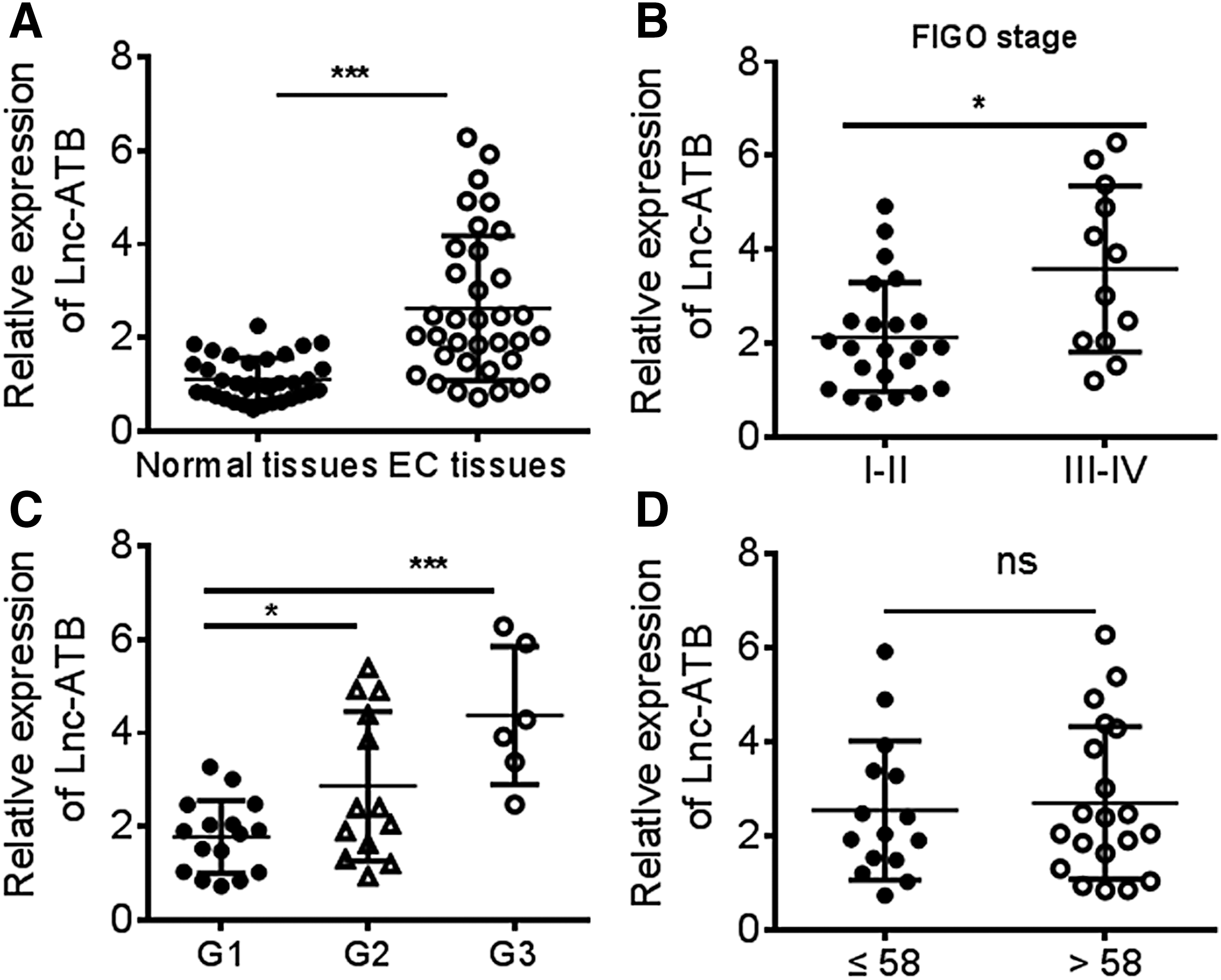
Lnc-ATB is overexpressed in EC patients.
Lnc-ATB interacts with miR-126 and inhibits its expression in EC
Interaction of LncRNA with miRNA is the critical mechanism to exert its function. In this study, bioinformatic analysis revealed putative complementary sequences for miR-126 in human Lnc-ATB and predicted miR-126 binding sites were found (Fig. 2A). In addition, the expression of Lnc-ATB and miR-126 in EC cell lines was determined and found that the expression of Lnc-ATB in EC cell lines RL95, HEC1A, AN3CA, and Ishakawa was higher than that in the immortalized human endometrial epithelial cell line EM-ER-A, but the expression of miR-126 was decreased in EC cells, indicating that miR-126 was negatively associated with Lnc-ATB in EC (Fig. 2A). Therefore, the siRNA assay to knock down the Lnc-ATB was performed, and the results showed that knockdown of Lnc-ATB upregulated the miR-126 level in RL95 and HEC1A cell lines, which had relatively higher expression of Lnc-ATB (Fig. 2B, C). Moreover, to verify the direct binging relationship, we constructed luciferase reporters that contained wild-type (WT) or mutated-type (MUT) miR-126 binding sites. The results showed that miR-126 mimics could significantly impair the luciferase activity of WT Lnc-ATB, but not the mutated-type in RL95 cell line (Fig. 2D). These results demonstrated that Lnc-ATB could function as competitive endogenous RNA (ceRNA) directly sponge to miR-126 and abrogate the availability of miR-126 in EC cells.
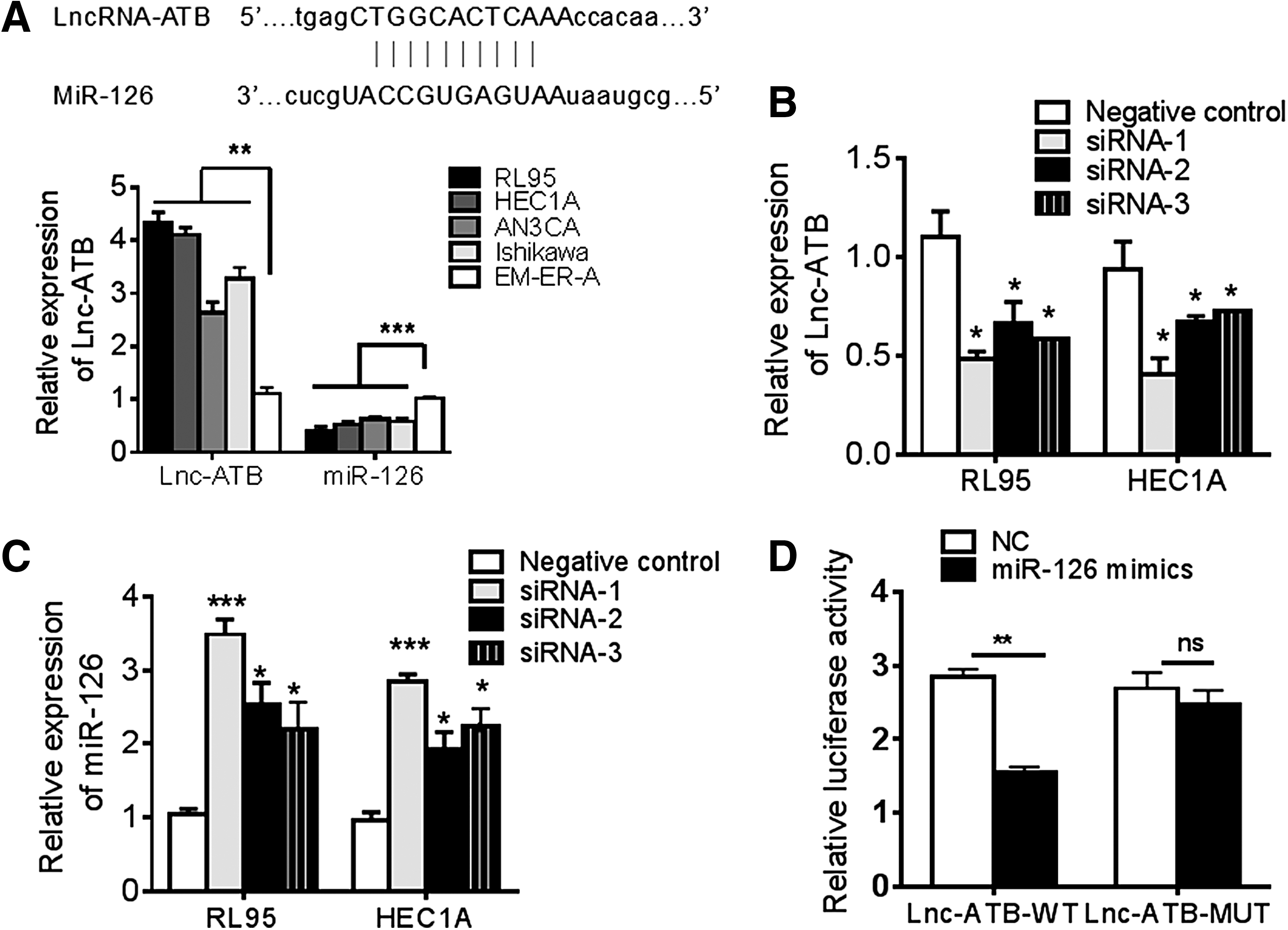
miR-126 is inhibited by Lnc-ATB in EC cells.
Downregulated miR-126 is a tumor suppressor in EC
Considering the tumor-suppressive role of miR-126 in cancer, this study investigated the clinical role of miR-126 in EC. The results of Q-PCR indicated that the miR-126 level was significantly reduced in EC tissues when compared with normal tissues (Fig. 3A). The correlation between miR-126 and Lnc-ATB was analyzed and found that the expression of Lnc-ATB negatively correlated with the miR-126 level in EC tissues (Fig. 3B). Contradictory to the role of Lnc-ATB, downregulated miR-126 predicted high FIGO stage (Fig. 3C) and poor differentiation of tumor cell (Fig. 3D) and the expression of miR-126 showed to be comparable in patients with different ages (Fig. 3E), which suggested the tumor-suppressive role of miR-126 in EC.
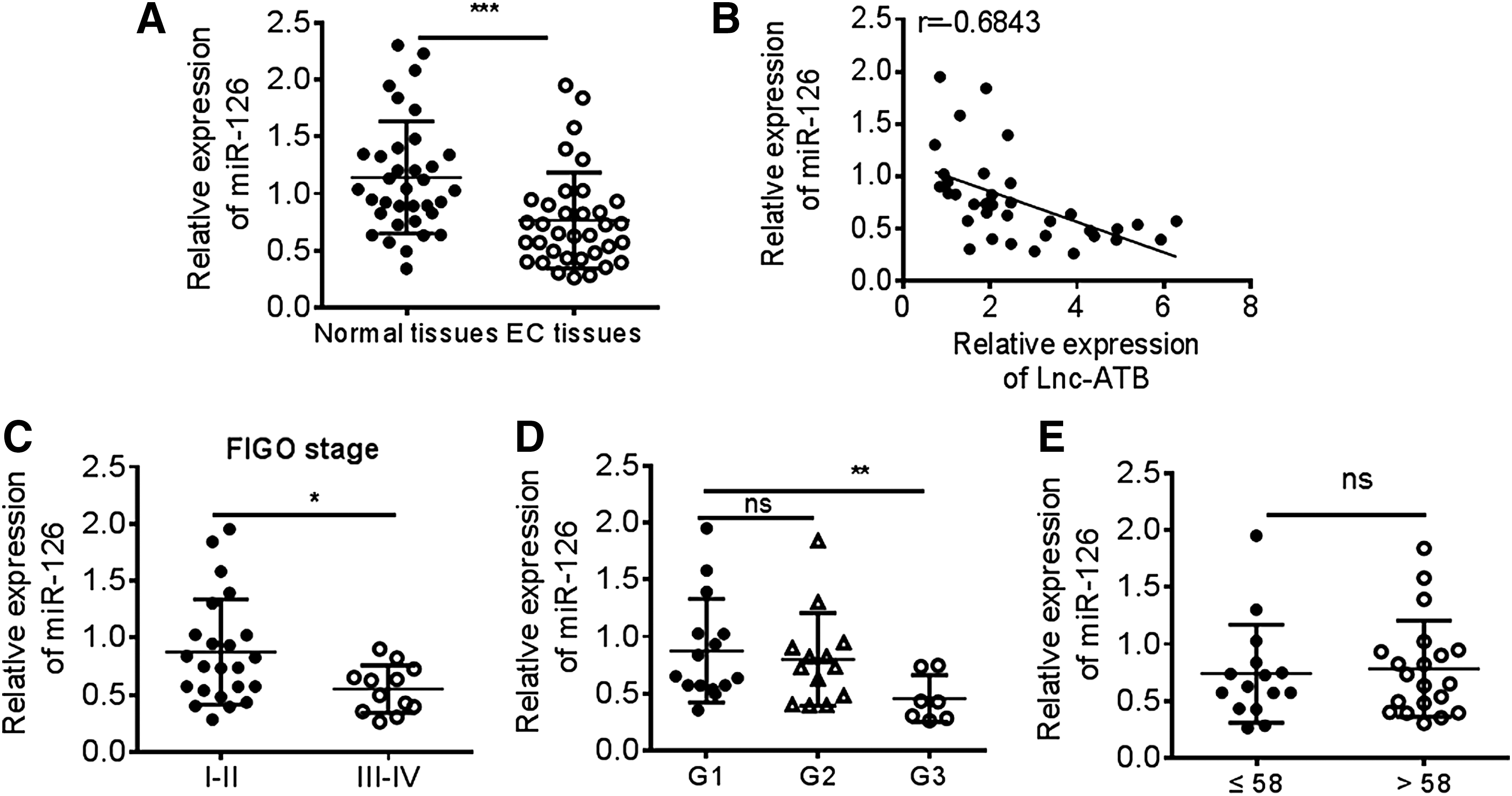
miR-126 is downregulated in EC patients.
Knockdown of Lnc-ATB in EC cells inhibits tumor growth through miR-126 signals
Since Lnc-ATB have been reported to regulate cell proliferation, apoptosis, and metastasis, 25 this study speculated that Lnc-ATB/miR-126 might promote the development of EC cells. The expression of Lnc-ATB in RL95 and HEC1A cell lines were knocked down. The cell vitality, apoptosis, and cell cycle were analyzed. The results showed that Lnc-ATB inhibition reduced the cell vitalities remarkably in two cell lines in 48 and 72 h (Fig. 4A). Thus, the apoptosis in 48 and 72 h were determined and found that knockdown of Lnc-ATB induced the upregulation of apoptosis by Hoechst assay (Fig. 4B) and flow cytometry (Fig. 4C). Meanwhile, the cell cycle was also showed to be arrested in G1/S (Fig. 4D). Because it was found that the tumor suppressor miR-126 was inhibited by Lnc-ATB, the upregulation of miR-126 induced by Lnc-ATB knockdown might contribute to these effects. Thus, abrogation of miR-126 by miR-126 inhibitor in two cell lines could significantly impair Lnc-ATB knockdown-induced cell cytotoxicity, apoptosis, and G1/S arrest (Fig. 4A–D). To clarify this mechanism, the key target genes of miR-126 in RL95 and HEC1A cell lines were investigated. PIK3R2/PTEN/PI3K/AKT signals and Sox2-related signals are critical mediators for cell proliferation, apoptosis, and cell cycle, which could be targeted and regulated by miR-126 (Fig. 4E). The results indicated that knockdown of Lnc-ATB significantly inhibited the expression of PIK3R2 and Sox2 through upregulation of miR-126, which was counteracted by miR-126 inhibitors. Correspondingly, the antiapoptosis factor Bcl-2 was downregulated and the caspase-3 was cleaved, and the expressions of cell cycle-related CCND1 and CDK4 were also reduced by Lnc-ATB knockdown but restored by miR-126, indicating that Lnc-ATB inhibition suppressed tumor growth in EC cell through regulation of miR-126-mediated pathway.
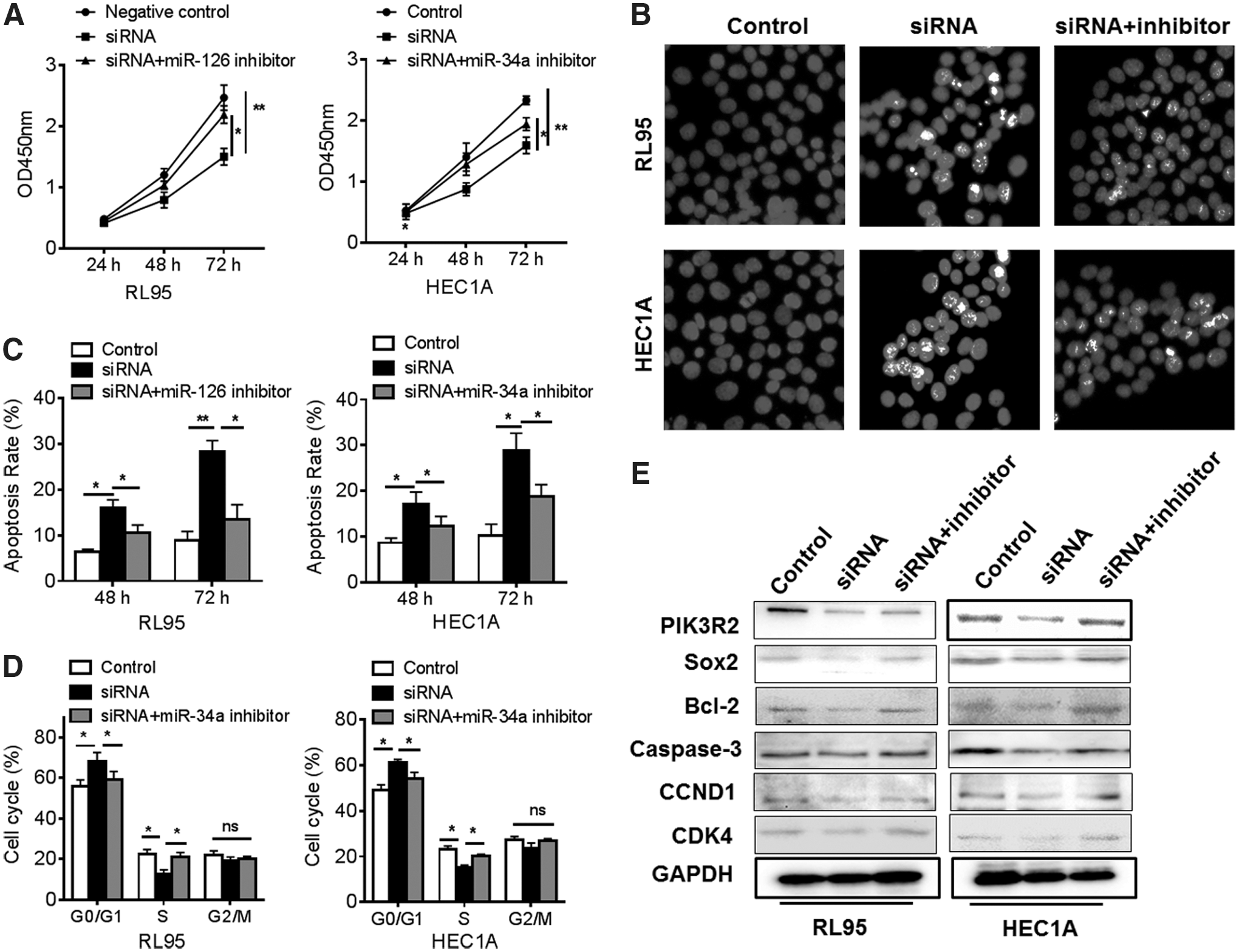
Knockdown of Lnc-ATB inhibits tumor growth through miR-126 in vitro. The RL95 and HEC1A cells were transfected with siRNA-Lnc-ATB, miR-126 inhibitors, and negative control; then
The EMT phenotype is also suppressed by Lnc-ATB inhibition
Similarly, this study also estimated the effects of Lnc-ATB inhibition on the ability of migration and invasion of EC cells. Knockdown of Lnc-ATB impaired the migration and invasion of RL95 and HEC1A cell lines, which was restored by miR-126 inhibitors (Fig. 5A, B). Then, the EMT phenotype was investigated and induced by TGF-β. The results indicated that knockdown of Lnc-ATB suppressed the TGF-β-induced EMT phenotype through upregulating the E-cadherin level and downregulating N-cadherin level, but miR-126 inhibitor could abrogate this process (Fig. 5C, D). Thus, Lnc-ATB also regulated the ability of migration and invasion through miR-126 in EC.

Knockdown of Lnc-ATB impairs EMT phenotype through miR-126 in vitro. The RL95 and HEC1A cells were transfected with siRNA-Lnc-ATB, miR-126 inhibitors, and negative control; then the ability of
Knockdown of Lnc-ATB in vivo efficiently induces tumor regression through miR-126 signals
To determine the function of Lnc-ATB in vivo in EC, the xenograft model of human RL95 cell lines was established. The conditional RL95 cells were transfected with lentivirus vector of siRNA-Lnc-ATB, miR-126 mimics, miR-126 inhibitors, or negative control. The results showed that knockdown of Lnc-ATB and restoration of miR-126 significantly induced tumor regression with reduced tumor volume and weight, but this antitumor effects was impaired by miR-126 inhibitor (Fig. 6A, B) The reduced expression of Lnc-ATB and increased expressed miR-126 were confirmed in tumor tissues (Fig. 6C). Besides, the proliferation index PCNA was estimated and found that knockdown of Lnc-ATB decreased the PCNA level in tumor tissues, which was restored by miR-126 inhibition (Fig. 6D). Mechanistically, Lnc-ATB regulated the tumor suppressor miR-126 during the tumor growth in vivo. miR-126 targeted and inhibited the expression of PIK3R2 and Sox2, and also impaired the EMT phenotype in tumor tissues (Fig. 6E).

Lnc-ATB inhibition in vivo induces tumor regression in vivo. RL95 cells transfected with lentivirus vector of siRNA-Lnc-ATB, miR-126 mimics, miR-126 inhibitor, or negative control were subcutaneously injected in rear flank of nude mice (five per group).
Discussion
Current therapy for EC includes surgery with adjuvant radiation or chemotherapy. Although the overall survival rate has been significantly improved by the development of early detection, 20% of the patients with EC have poor prognosis with a median survival rate of just a year due to the resistance to chemotherapy and disease recurrence. 26 This study found that Lnc-ATB negatively correlated with tumor suppressor miR-126 and predicted a poor clinical outcome for EC patients. Knockdown of Lnc-ATB promoted tumor regression through miR-126-mediated antitumor effects in vitro and in vivo.
Amounting evidences demonstrated that the ncRNAs have been dysregulated during the development of EC. 5 The up- or downregulated ncRNAs could be recognized as efficient diagnostic and prognostic markers for EC patients. Many miRNAs have been dysregulated in EC. miR-30c, miR-103, miR-106a, miR-107, miR-152, miR-181a, miR-185, miR-200, and so on were found to be upregulated and miR-1, miR-133a, miR-504, miR-133b, miR-137, miR-125, and miR-129 were downregulated in EC tissues. 27 Tumor suppressor Let-7 inhibits cancer cell proliferation by inhibiting oncogenes such as RAS and c-MYC. Liu et al. found that 7 members of the let-7 family are downregulated in EC when compared with normal endometrium. 28 Dong et al. found that miR-130 was reduced by mutant p53 and that restoration of its expression was associated with the attenuation of EMT phenotype, which plays a key role in tumor metastasis. 29 This study reported that downregulated miR-126 functioned as tumor suppressor and negatively correlated to FIGO stage and differentiation. Decreased miR-126 in lung cancer cells upregulated its target gene PIK3R2 to activate PTEN/PI3K/AKT signaling pathway for tumor growth. The upregulation of miR-126 in glioma cells also contributed to the aberrant activation of the ERK signaling and inhibits cell proliferation and invasion through targeting KRAS. 22 It is to be noted that type I EC is characterized by PTEN, KRAS, and PIK3CA mutations and type II EC shows HER2 amplification and TP53 mutations. 30 Jayaraman et al. have found that miR-126 was downregulated during the development of EC. 31 miR-126 also inhibited the migration and invasion of EC cells by targeting insulin receptor substrate. 32 Thus, miR-126-mediated signals might be critical for the progression of EC, which partly depend on PIK3R2/PI3K/AKT and Sox2-related apoptosis, proliferation, and metastasis pathway in this study. 33
The role of LncRNAs in EC is limited. Zhao et al. reported that Wnt/beta-catenin signaling pathway significantly induced transcription of MALAT1 by interaction of TCF4 with the binding site of the MALAT1 promoter region in type-1 EC. 34 He et al. reported that the expression of HOTAIR was increased in nearly three quarters of EC, which was associated with poor dedifferentiated tumor cells. 35 In this study, increased expression of Lnc-ATB functioned as oncogene and positively correlated to high FIGO stage and differentiation. LncRNAs can act as miRNA decoys, with the sequestration of miRNAs favoring expression of repressed target messenger RNAs (mRNAs). 36 This study found that Lnc-ATB interacted and inhibited miR-126, leading to impairing the miR-126-mediated inhibition of PIK3R2 and Sox2-related apoptosis and cell cycle pathway. Similarly, Lnc-ATB is upregulated by TGF-β in many cancers and is physically associated with the miR-200 family to upregulate ZEB1 and ZEB2 levels for EMT phenotype. 20 Recently, Lnc-ATB promoted proliferation, migration, and invasion in bladder cancer by suppressing miRNA-126. 37 In esophageal squamous cell carcinoma (ESCC), Lnc-ATB functions as a molecular sponge for miR-200b and Kindlin-2 to promote cell proliferation and migration of ESCC cells and Lnc-ATB predicted poor prognosis for ESCC patients. 38
Taken together, this study found the tumorigenic role of Lnc-ATB during the development of EC in vitro and in vivo. Lnc-ATB inhibited the availability of tumor suppressor miR-126 to impair the function of target pathway of miR-126, including PIK3R2/PI3K/AKT and Sox2-related apoptosis, proliferation, and metastasis pathway in EC.
Footnotes
Acknowledgment
This study was supported by First Hospital of Xi'an.
Disclosure Statement
No competing financial interests exist.