
Abstract
Objective:
Apoptosis plays an important role in both carcinogenesis and cancer treatment. Drugs or treatment strategies that can restore the apoptotic signaling pathways have the potential to eliminate cancer. Caspase 8 (CASP8) plays a vital role in the propagation of an enzymatic cascade that results in cell apoptosis.
Methods and Results:
In this study, the authors investigated the inhibitory effects of a HIV Gag virus-like particles (VLPs) that are incorporated with an active CASP8 (Gag-CASP8-VLPs) on the growth of breast cancer. Their data have shown that Gag-CASP8-VLPs, pseudotyped by the stomatitis virus G protein (VSV-G), can efficiently enter and deliver active CASP8 into breast cancer cells, leading to massive cell apoptosis and death. Interestingly, an injection of Gag-CASP8-VLPs in the tumor tissues of a 4T1 mouse breast cancer model can effectively inhibit tumor growth, and the earlier the Gag-CASP8-VLPs is administered, the more profoundly the tumor growth is inhibited.
Conclusions:
Overall, Gag-CASP8-VLPs can deliver CASP8 into breast cancer cells, induce cell apoptosis, and inhibit tumor growth.
Introduction
Breast cancer is the most common cancer worldwide and is the fifth most common cause of cancer-related death in women. 1 Over the past two decades, in addition to cytotoxic chemotherapy, the medical treatment of breast cancer has evolved to a more targeted approach that has greatly improved patient outcomes. However, metastatic breast cancer remains incurable, with an estimated 5-year overall survival rate of only 23%. 2 An important and unexpected finding for breast cancer treatment is the emergence of therapy-resistant cancer cells. 3 –5 Therefore, the development of a novel potential treatment strategy is still needed for further improvements in clinical outcomes and survival rates for breast cancer patients.
Apoptosis is a naturally occurring process by which a cell is directed to programmed cell death. It is a critical mechanism that allows multicellular organisms to maintain tissue integrity and function and to eliminate damaged or unwanted cells. Dysregulation of apoptosis is a key process in cancer development and progression. 6 In other words, the ability of cancer cells to avoid apoptosis and continue to proliferate is one of the fundamental steps in cancer development. 7 It has become clear that some oncogenic mutations disrupt apoptosis, leading to tumor initiation, progression, or metastasis. On the contrary, the silencing of several tumor suppressor genes may lead to an increased risk of tumor formation and progression. Some studies have suggested that caspase 8 (CASP8) and maspin expression are reduced in breast cancer cells (MCF-7, MB231, SKBR3, and HCC1937) due to promoter methylation. 8 Thus, apoptosis is a biological link between cancer genetics and cancer therapy, and the genes and proteins controlling apoptosis can be potential drug targets. CASP8 plays a vital role in the propagation of an enzymatic cascade that results in cell apoptosis. It was shown that the Adv-CASP8-mediated gene transfer strongly induced apoptosis in U251 human glioma cells and suppressed the in vivo growth of U251 xenografts. 9 This study also demonstrated that the introduction of CASP8 via an Adv-CASP8 vector augmented apoptosis and inhibited peritoneal dissemination of human gastric carcinoma cells. CASP8, therefore, shows potential for breast cancer therapy.
Virus-like particles (VLPs) are self-assembling, nonreplicating, nonpathogenic, and genomeless particles. Notably, VLPs have similar sizes and conformations to those of intact infectious viruses. VLPs are attractive candidates for vaccine strategies due to their safety and potency to stimulate cellular and humoral immune responses. 10 –13 Moreover, VLPs can also be used as efficient vehicles to deliver foreign genes and small molecules. 14 –16 Recently, a bacteriophage P22 VLP containing cytochrome P450 (CYP) enzymes with photosensitizer and targeting moiety at the surface has been developed to combinatory treatment of breast cancer. 17 –19 HIV Gag (group-specific antigen) can efficiently form VLPs even when expressed in the absence of other viral proteins. 20 A previous study suggested that the Gag-VLP pseudotyped with vesicular stomatitis virus G glycoprotein (VSV-G), which mediates efficient entry into a broad spectrum of cells, might enhance intracytoplasmic delivery of VLPs components and therefore improve their immunogenicity. 21 In the present study, the authors have generated a Gag-CASP8 fusion protein that is able to form mature VLPs in the presence of HIV protease, and these VLPs contain significant amounts of active CASP8. In addition, this study shows that after being pseudotyped with VSV-G, these CASP8-VLPs are able to efficiently enter the breast cancer cells and induce apoptosis. Furthermore, in a 4T1 mouse breast cancer model, tumor growth is significantly inhibited when the tumor formation site within mice is injected with Gag-CASP8-VLPs.
Materials and Methods
Cells, antibodies and chemicals
Human embryonic kidney 293T cells, mouse breast tumor cell line 4T1 cells, and human breast cancer cell line MCF-7 were cultured in Dulbecco's modified Eagle's medium (DMEM), supplemented with 10% fetal bovine serum and 1% penicillin and streptomycin. DNA transfection into 293T cells was performed with a PEI method. 22
CASP8 mouse monoclonal antibody was purchased from Cell Signaling Technology (Danvers, MA). Rabbit anti-HIV P24 antibody was obtained from the NIH AIDS Research and Reference Reagent Program. Z-VAD-FMK (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoromethylketone) and acridine orange/ethidium bromide (AO/EB) were purchased from Enzo Life Sciences, Inc. (Farmingdale, NY) and Life Technologies, Inc. (Burlington, ON, Canada), respectively.
Plasmids
A human CASP8-expressing plasmid was kindly provided by Dr. S. M. Srinivasula (Thomas Jefferson University). A pSVCMVGag-CASP8 fusion protein-expressing plasmid was constructed by replacing the HIV Pol gene with a PCR-amplified full-length human CASP8 cDNA in a SVCMV-Gag-Pol plasmid, as described previously. 23 An HIV-1 protease cleavage site (SQNY/PIV) was inserted between Gag and CASP8 (Fig. 1A). The helper packaging construct pCMV-Δ8.2 and the vesicular stomatitis virus G glycoprotein (VSV-G) expresser were previously described. 24 HIV-1 RT/IN/Env tri-defect proviral plasmid (ΔRI/E−/Gluc+) used for generating a stable cell line was described previously. 23
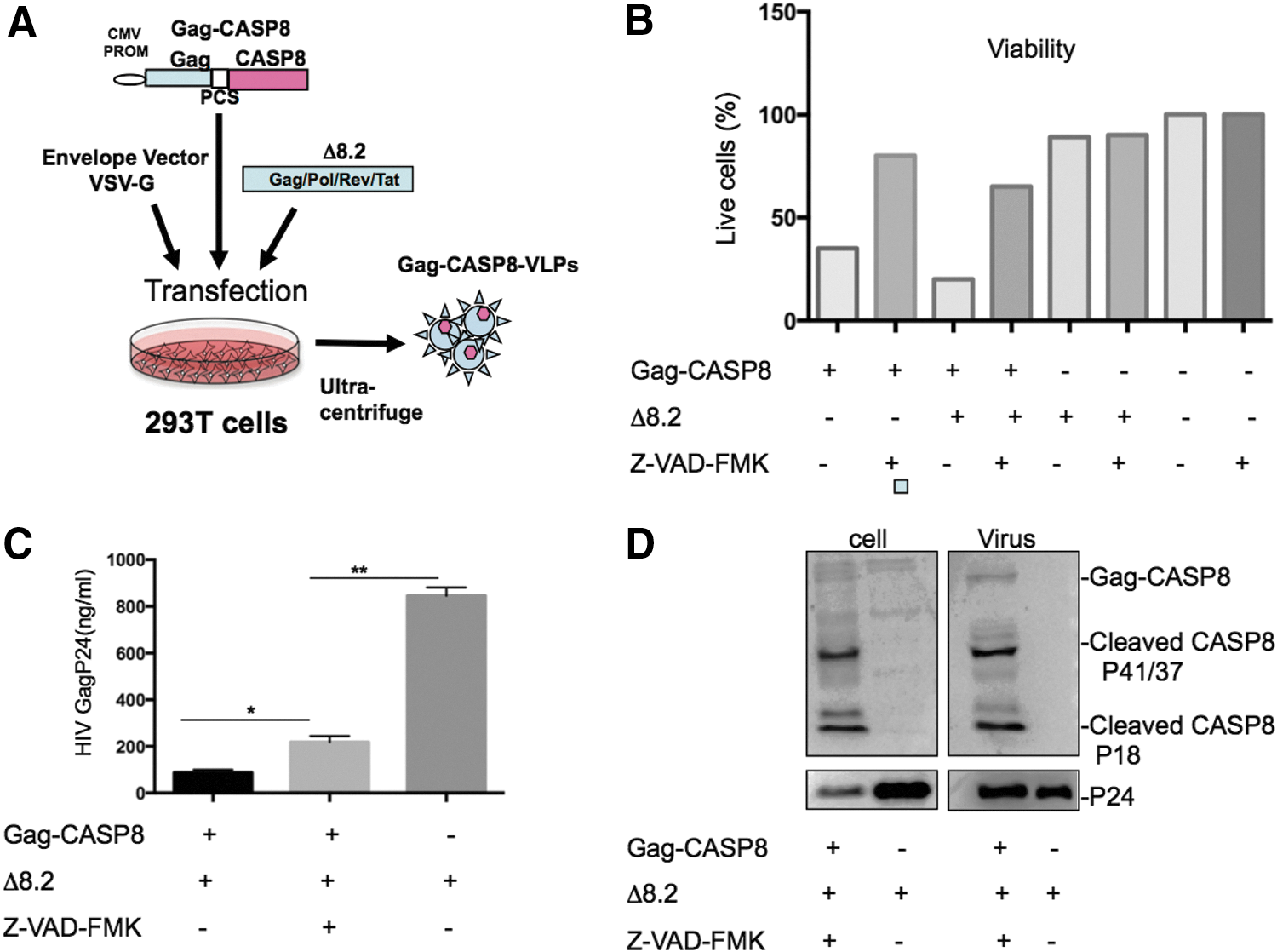
Transfection and VLP production
VSV-G-pseudotyped Gag-CASP8 or Gag-VLPs were produced by transfection 293T cells with the appropriate pSVCMV-Gag-CASP8, pCMV-Δ8.2, and VSV-G plasmids. To inhibit apoptosis induced by CASP8 in VLPs-producing cells, Z-VAD-FMK was added into culture medium to a 20 μg/mL final concentration 5 h after transfection. At 48 h posttransfection, VLPs were concentrated from the supernatant by ultracentrifugation with a Beckman Ti70 rotor at 32,000 rpm for 2 h, and virus titers were quantified by using HIV-1 p24 Antigen Capture Kit obtained from the AIDS Vaccine Program of the Frederick Cancer Research and Development Center.
Transduction
The 4T1 or MCF-7 cells were plated at 4 × 10 3 cells per well in a 96-well plate for 24 h before transduction. Equal amounts of VSV-G-pseudotyped Gag-VLPs or Gag-CASP8-VLPs (adjusted by the p24 level) or serial twofold dilutions of VLPs were incubated with target cells in DMEM. Between 12 and 48 h posttransduction, the apoptotic cells or the percentage of survived cells were detected.
Establishment of 4T1 cell line that stably expresses Gaussia luciferase
HIV-1 ΔRI/E−/Gluc+ were produced, as previously described, 23 and used to infect 0.5 × 10 6 of 4T1 cells. After 4 d, the GLuc activity of supernatant from infected cells was tested, and the cells were used in further studies.
AO/EB staining
In brief, the equal amounts of VSV-G-pseudotyped Gag-VLPs or Gag-CASP8-VLPs (adjusted by the p24 level) were used to treat 4T1 cells seeded in 96-well plate. After 12–24 h, EB/AO dye mixture (10 μL) was added to each well, and cells were viewed under an Axiovert 200 fluorescence microscope (Carl Zeiss, Jena, Germany).
Cell proliferation assay (Cell Counting Kit-8)
The 4T1 or MCF-7 cells in a 96-well plate were untreated or treated with Gag-VLPs, Gag-CASP8-VLPs for 24–48 h. Cell Counting Kit-8 (CCK-8) solution was added to each well and incubated for 2 h. Finally, the absorbance of the sample taken from each well was measured using a microplate reader at 450 nm. Cell survival was calculated as the percentage of the optical density (OD) of treated well relative to that of untreated well.
Western blot
Twenty-four hours posttransfection, the 293T cells were harvested, lysed in RIPA lysis buffer (140 mM NaCl, 1.0% NP-40, 0.5% sodium deoxycholate, 0.05% SDS, pH 7.2), and directly loaded onto the 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Meanwhile, the above-described purified Gag-VLPs and Gag-CASP8-VLPs were also lysed with 1% Triton X-100 and loaded onto 12% SDS-PAGE. CASP8 and the viral protein Gag-p24 were detected with anti-CASP8 and anti-p24 antibodies.
Immunohistochemistry
MCF-7 cells were grown on glass cover slips (12 mm2) in a 24-well plate for 12 h. Then, Gag-VLPs or Gag-CASP8-VLPs were added to the well and incubated for 2–24 h. Immunohistochemistry was carried out according to the manufacturer's instructions. In brief, the slides were incubated overnight with mouse anti-CASP8 antibody at a 1:200 dilution, followed by the corresponding second peroxidase-conjugated antibody and stained by 3,3-N-diaminobenzidine tetrahydrochloride (DAB). Then, cells on the cover slip were viewed under a microscope.
Animal experiments
Six-week-old female BALB/C mice were purchased from the Third Military Medical University (Chongqi, China). The animal protocol was approved by the Zunyi Medical College Animal Care and Use Committee. After acclimatization over 7 d, each mouse was injected subcutaneously with 1 × 10 6 4T1 cells at the right second mammary fat pads (day 0). Protocol 1: At 10 d, when the tumor size was ∼70 mm 3 , the mice were divided into different treatment groups as follows. Group I: Gag-CASP8-VLPs (6 mice); Group II Gag-VLPs (6 mice); and Group III: phosphate-buffered saline (PBS; 6 mice). The mice received three injections of 100 μL of VLPs (100 ng p24) or PBS only to the center of the tumor mass at days 10, 13, and 16 during the entire experimental period. The tumor volume and body weight were measured twice per week, and mice were sacrificed at day 22. Protocol 2: Group I: Gag-CASP8-a, mice received a Gag-CASP8-VLP treatment at the day of injection of 4T1 cells (day 0) at the same body place, followed by another two treatments at days 3 and 6; Group II: Gag-CASP8-b, mice received a Gag-CASP8 injection at the center of the tumor mass at days 3, 6, and 12; Group III: Gag; or Group IV: mice received an injection of Gag-VLPs or PBS at the center of the tumor mass at days 3, 6, and 12—four mice for each group. The experimental period lasted 30 d. The tumors were collected for histological and immunohistochemical analyses.
Hematoxylin and eosin staining and immunofluorescence
The tumor tissues were placed in 10% neutral buffered formalin solution (Sigma-Aldrich, St. Louis, MO) before being processed and paraffinized. Then, the samples were sectioned and stained with hematoxylin and eosin solution (Merck, Germany). For immunofluorescence, the tissue sections were incubated overnight with mouse anti-CASP8 antibody (1:200) at 4°C followed by incubated with Cy3-labeled anti-mouse IgG for 30 min in the dark. Following DAPI counterstaining, the tissue section was examined and photographed with an Axiovert 200 microscope (Carl Zeiss).
Statistical analysis
The data were expressed as the mean ± standard deviation. The significance of the data was determined by one-way ANOVA. p-Values of less than 0.05 were considered statistically significant.
Results
Generation of VSV-G-pseudotyped Gag-VLPs that are incorporated with an active CASP8
To deliver an active proapoptotic protein into cancer cells, the authors first developed a CASP8-VLP delivery system shown in Figure 1A. It is well known that the HIV-1 structural protein Pr55Gag is sufficient to form HIV-1-based VLPs. 20 They, therefore, generated a fusion protein construct, termed pSVCMV-Gag-CASP8, by fusing full-length CASP8 to the C-terminus of HIV-1 Pr55 (Fig. 1A). An HIV-1 protease cleavage site (SQNY/PIV) was inserted between Gag and CASP8 to release the CASP8 protein in the VLPs through specific cleavage mediated by HIV-1 protease. To produce Gag-CASP8-VLPs, 293T cells were cotransfected with pSVCMV-Gag-CASP8 and packaging pCMV-Δ8.2 plasmid, which can provide sufficient amounts of HIV-1 protease for the cleavage of Pr55Gag precursor and release of CASP8. To enhance the carcinoma cell-targeting efficiency of Gag VLPs, a vesicular stomatitis virus G glycoprotein (VSV-G) plasmid was also included in the transfection mixture (Fig. 1A).
To test whether the expression of Gag-CASP8 could induce apoptosis in the VLP-producing cells and to test how to inhibit its toxicity to produce sufficient Gag-CASP8-VLPs, they transfected 293T cells with pSVCMV-Gag-CASP8 alone or cotransfected with pCMV-Δ8.2 and cultured cells in the presence or absence of caspase inhibitor Z-VAD-FMK (carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoromethylketone) (Fig. 1B). After transfection, cell viability was observed under a microscope and assessed by trypan blue exclusion. After 24 h of transfection and in the absence of Z-VAD-FMK, massive cell death was observed, and the surviving cell fractions expressing Gag-CASP8 alone and coexpressing pCMV-Δ8.2 were only ∼35% and 20%, respectively (Fig. 1B, bars 1 and 3). However, in the presence of caspase inhibitor Z-VAD-FMK, the survival rate of cotransfected cells reached ∼70%–85% (Fig. 1B, bars 2 and 4).
In addition, the authors measured the levels of VLPs in the supernatants from each transfected cell culture. In agreement with the above observation, the Gag-CASP8-VLP production level in the absence of Z-VAD-FMK was very low due to massive cell death. By using Z-VAD-FMK to inhibit cell apoptosis, a significant amount of Gag-CASP8-VLPs could be rescued (Fig. 1C).
The authors then characterized the expression of Gag-CASP8 in pSVCMV-Gag-CASP8 and pCMV-Δ8.2-transfected 293T cells and its incorporation into Gag-VLPs by western blot with the specific anti-CASP8 antibody. As shown in Figure 1D, while low levels of Gag-CASP8 were detected, abundant levels of cleaved CASP8 p41/37 and p18 were observed in both pSVCMV-Gag-CASP8-transfected cells and purified CASP8-VLPs, demonstrating that CASP8 was expressed and readily activated in 293T cells 24 h after transfection. Caspase inhibitor Z-VAD-FMK did not interfere with the expression and cleavage of CASP8. Moreover, the HIV Gag precursor in the VLPs was also efficiently processed, as an abundance of cleaved capsid protein (p24) was detected in both Gag-CASP8- and pCMV-Δ8.2-transfected cells.
VSV-G-pseudotyped Gag-CASP8-VLPs can efficiently deliver CASP8 into breast cancer cells and induce cell apoptosis
To investigate the effect of Gag-CASP8-VLPs on cell survival, the mouse breast tumor cell line 4T1 was treated with VSV-G-pseudotyped Gag-VLPs or Gag-CASP8-VLPs in different doses. The results showed that a large number of Gag-CASP8-VLP-treated cells were killed (Fig. 2A, lower panel (c and d)). Twenty-four hours treatment of cells with Gag-CASP8-VLPs at the p24 doses of 5, 10, or 20 ng induced ∼55%, 71%, and 87% cell death, respectively (Fig. 2A, upper panel). All Gag-CASP8-VLP-treated 4T1 cells completely died 48 or 72 h after transfection (data not shown). The above observation was further confirmed by using 4T1 cells that stably expressed secreted Gaussia luciferase (Gluc) protein. When the untreated and Gag-VLP-treated cells grew with time, they released more Gluc into the medium, and the levels of Gluc activity in culture medium gradually increased (Fig. 2B). However, after being treated with Gag-CASP8-VLP, the Gluc activity in the 4T1 cell culture dramatically declined with time, demonstrating that CASP8-VLP indeed blocked cell growth (Fig. 2B). It is worth noting that the Gag-VLP treatment also resulted in modest cell death (∼22%), compared with the untreated cells (Fig. 2A, B). In addition, AO/EB staining was used to detect apoptosis in VLP-treated 4T1 cells. While no significant apoptosis was detected in mock-treated cells (Fig. 2C-a), a large amount of early-stage apoptotic cells (yellow-green) was found in Gag-CASP8-VLP-treated cells (Fig. 2C-c). With longer treatment, late apoptotic and necrotic cells (orange-red nuclear staining) were also clearly seen (Fig. 2C-d).
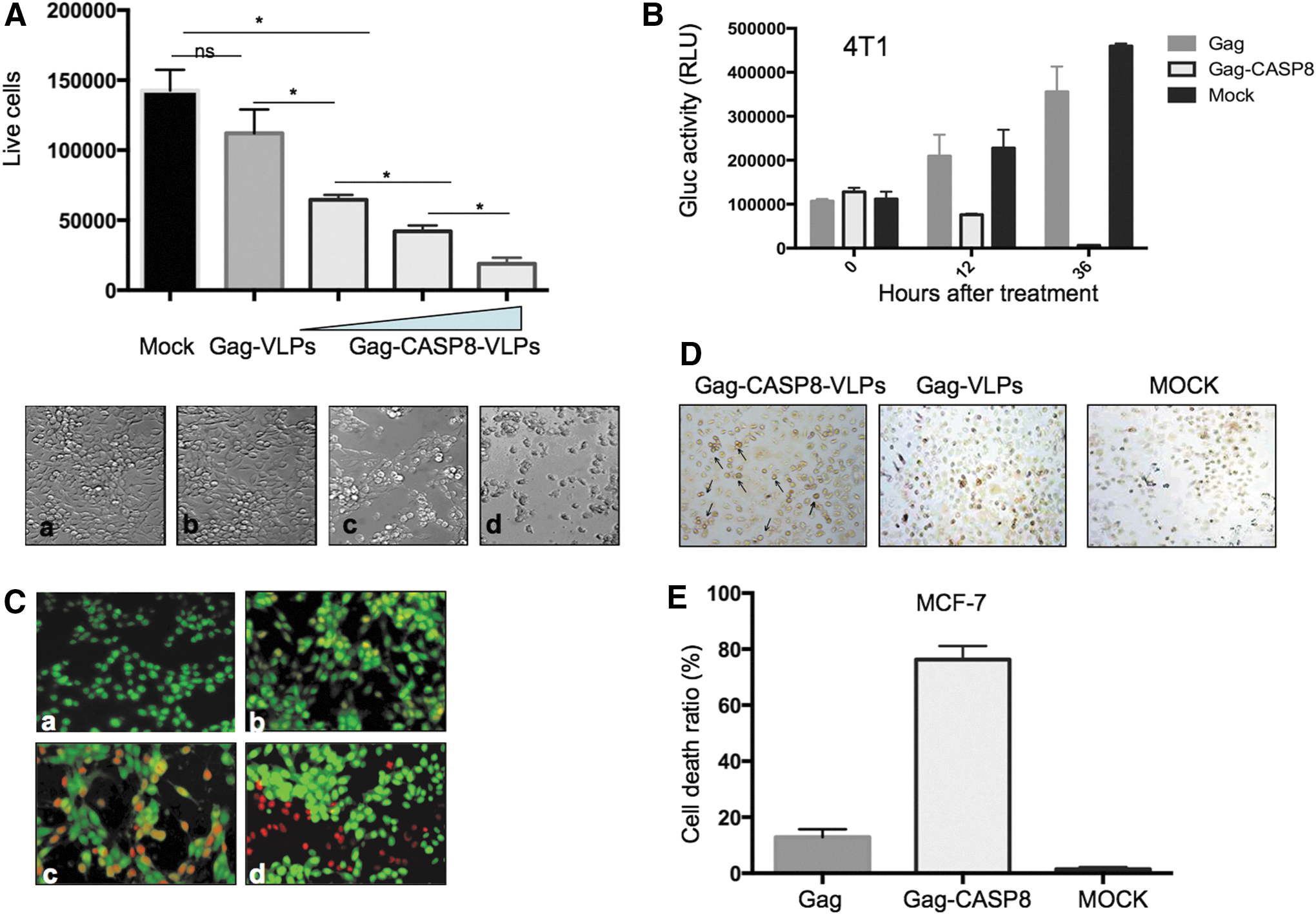
Virus-like particles deliver CASP8 into breast cancer cells and induce cell apoptosis.
Then, the authors tested whether Gag-CASP8-VLP was able to target human cancer cell, MCF-7 cell line. Immunohistochemistry study showed that after 2 h incubation with Gag-CASP8-VLP, a significant positive signal was found on the cell membranes of Gag-CASP8-VLP-treated cells (Fig. 2D-c), indicating that Gag-CASP8-VLPs have targeted MCF-7 cells (Fig. 2D-c), After 24 h of treatment, the viability of MCF-7 cells was determined by means of the Cell Counting Kit-8 (CCK-8) assay. Similar to 4T1 cells, the Gag-CASP8-VLP treatment induced ∼76% MCF-7 cell death, while Gag-VLPs only resulted in ∼12% cell death. Overall, these results indicate that Gag-CASP8-VLP can effectively enter and kill breast cancer cells.
In vivo delivery of VSV-G-pseudotyped Gag-CASP8 VLPs significantly inhibits the growth of tumors in the 4T1 mouse breast cancer model
The potential anticancer effect of Gag-CASP8 VLPs in mice was determined with breast cancer derived from 4T1 cells. The summary of the experiments is depicted in Figure 3A and described at materials and methods. In brief, the mice received three injections of Gag-CASP8-VLPs, Gag-VLPs (100 ng p24 in 100 μL PBS), or PBS to the center of the tumor mass at days 10, 13, and 16. To investigate the efficiency of CASP8 delivery in vivo, 2 mice from each group were sacrificed in 12 h after the first injection of VLPs, and the tumor sections were subjected to immunofluorescence analysis by using anti-CASP8 antibody (Fig. 3B). In Gag-CASP8 VLPs group, a strong CASP8 signal was detected in the tissue, especially in the cytoplasm. As expected, a very weak CASP8 signal was found in Gag-VLP and PBS-treated cells (Fig. 3B). These data suggest that Gag-CASP8-VLP injection can efficiently deliver exogenous CASP8 into tumor cells.

In vivo Gag-CASP8-VLP treatment inhibits mouse breast cancer progress.
All other mice were sacrificed at day 22. The primary mammary tumors were dissected and weighed at the time of sacrifice. As shown in Figure 3C, compared with the PBS injection group, Gag-CASP8-VLP injection at days 10, 13, and 16 rapidly inhibited tumor growth. At day 22, the volumes and weights of Gag-CASP8-VLP-treated tumors were only 30% of those of the PBS-treated tumors (Fig. 3D, E). No significant difference was detected between Gag-VLP and PBS treatment. The side effects, such as the loss of body weight and changes in gait, were not observed in Gag-CASP8-VLP- or Gag-VLP-treated mice during the observation period.
Early administration of VSV-G-pseudotyped CASP8-VLPs displays stronger inhibitory effect on tumor growth
The authors then tested the inhibitory effect of VSV-G-pseudotyped Gag-CASP8-VLPs on tumor growth by injecting VLPs at the earliest time. As shown in Figure 4A, the Gag-CASP8-a group received injections at days 0, 3, and 6, while the Gag-CASP8-b, Gag-VLPs, and PBS were given injections at days 3, 6 and 12. After 30 d, the tumor growth evaluation and the histological and histomorphometrical examinations were performed. The results revealed that the administration of VSV-G-pseudotyped Gag-CASP8-VLPs on the day of 4T1 cell injection (CASP8-a) reduced the volumes and weights of the tumors to 5% of those in the control groups (Gag-VLPs and PBS), while in the Gag-CASP8-b group, the tumor size was reduced only to 35% of that of the control (Fig. 4B, C). These results suggest that the earlier the Gag-CASP8-VLPs are injected, the more profound the achieved effects will be.

In vivo early treatment with Gag-CASP8 efficiently inhibits the growth of mouse breast cancer.
Discussion
In this study, the authors have developed apoptotic protein CASP8-loaded lentivirus-like particles (Gag-CASP8-VLPs) that are able to assemble through an HIV Gag-CASP8 fusion protein. By being pseudotyped with VSV-G glycoprotein, these Gag-CASP8-VLPs can efficiently enter breast cancer cells and kill the cells through inducing apoptosis. Furthermore, in a 4T1 mouse breast cancer model, they have demonstrated that the injection of Gag-CASP8-VLPs in tumor tissues can significantly inhibit tumor growth, and the earlier the Gag-CASP8-VLPs were administered, the more profound the inhibition of the tumor growth was observed.
Apoptosis is triggered through two main signaling mechanisms: the “intrinsic” and “extrinsic” pathways. 25,26 In both pathways, signaling results in the activation of a family of Cys (Cysteine) proteases or caspases. Activation of CASP8, 1 of 11 human caspases, is central to the process of apoptosis. 27 –29 Similar to all other caspases, CASP8 is synthesized as an inactive single polypeptide chain zymogen procaspase and is activated by proteolytic cleavage, through either autoactivation after recruitment into a multimeric complex (Fas-associated death domain) or trans-cleavage by other caspases. 30 Active CASP8 then activates downstream caspases 3 and 7, committing the cell to apoptosis. Therefore, CASP8 is a potent proapoptotic molecule that is present in the cell. In this study, a CASP8-loaded lentivirus-based VLP system was generated to directly deliver active CASP8 into tumor cells. Since an HIV-1 protease cleavage site (SQNY/PIV) was inserted between Gag and CASP8 in the Gag-CASP8 expression plasmid, CASP8 protein was efficiently released from the Gag-CASP8 fusion protein through the HIV protease cleavage during the assembly of CASP8-VLPs (Fig. 1D). Interestingly, the authors observed that most of the CASP8 protein in VLPs was in its active form (p41/37, p18) and could readily induce cell apoptosis (Fig. 2A, B). Previous studies demonstrated that in addition to Fas-l stimulation, the activation of CASP8 might also occur via a variety of different mechanisms. In certain physiological conditions, abundant expression of CASP8 precursor proteins may also result in self-oligomerization and in subsequent self-cleavage and activation in the absence of any external apoptotic signal. 31 Interestingly, in the Gag-CASP8 fusion protein-transfected 293T cells, the overexpression of Gag-CASP8 fusion protein also led to CASP8 activation, which induced apoptosis of CASP8-VLP-producing 293T cells, which reduced the production levels of CASP8-VLPs (Fig. 1C). Hence, the authors added a caspase inhibitor Z-VAD-FMK to CASP8-VLP-producing 293T cells and showed that its presence could significantly inhibit apoptosis in Gag-CASP8-transfected 293T cells and significantly increase the production of CASP8-VLPs (Fig. 1C). However, more studies are still required for condition optimization to reduce the apoptosis of CASP8-VLP-producing 293T cells and subsequently increase the production levels of Gag-CASP8-VLPs.
To mediate efficient entry into cancer cells, the authors pseudotyped the Gag-CASP8-VLPs with VSV-G, which is frequently used in gene transfer with lentiviral vectors. In contrast to general lentiviral vector systems, Gag-CASP8-VLPs do not contain any viral genomic materials. Therefore, this eliminates the potential risk of viral genome integration into the host chromosome. This is particularly important since it is possible that this anticancer agent will be used in vivo if the anticancer efficiency and safety are proved in the animal models. At this point, their investigation indeed showed that Gag-CASP8-VLPs efficiently induced tumor cell death and inhibited tumor formation in a mouse model. All these observations provide evidence of the feasibility of a novel antitumor strategy.
Since the cell entry of Gag-CASP8-VLPs depends on the VSV-G glycoprotein present on the VLPs, it is expected that these VLPs will target both tumor cells and normal cells. Even though no obvious side effects were observed in mice injected with Gag-CASP8-VLPs, it is necessary to further optimize this Gag-CASP8-VLP-targeting system. One critical aspect of this strategy is to direct CASP8-VLPs to specifically or preferentially target cancer cells. Previous studies have revealed several unique receptor proteins or ligands that are over expressed in various tumor cells, such as human epidermal growth factor receptor 2 (HER2), interleukin 11 receptor, alpha subunit (IL-11Ra), 78 kDa glucose-regulated protein (GRP78), ephrin type-A receptor 5 (EphA5), and folate and estrogen receptor. 32 –34 Among them, HER2 is one of the most well-characterized tumor-associated antigens for cancer therapy and plays an important role in the growth and progression of breast cancer. Overexpression of HER2 in various cancers and the availability of its extracellular region make it a clinically useful target for the development of tumor antigen-specific agents. Previous reports have shown that the placement of a single-chain antibody directed against the tumor-associated antigen HER2 on the surface of enveloped vaccinia virus is a step toward selective tumor cell targeting. 34 Encouragingly, some recent studies have shown that biocatalytic bacteriophage P22 VLPs can be chemically covered with PEG-containing folic acid or an estradiol based targeting ligand. Above studies also demonstrated that those VLPs containing targeting moiety at the surface were recognized and internalized into tumor cells, including ER+ breast cancer cells. 18,19 Therefore, their system can provide an opportunity for linking these tumor-specific targeting molecules to the surfaces of VLPs and consequently drive active CASP8 specifically or preferentially into cancer cells to achieve specific anticancer activity.
In conclusion, the authors have developed and demonstrated a Gag-CASP8-VLP that is able to assemble through an HIV Gag-CASP8 fusion protein and efficiently enter breast cancer cells and induce apoptosis and cell death in vitro. In addition, the administration of Gag-CASP8-VLPs to tumor tissues in a 4T1 mouse breast cancer model effectively inhibited tumor growth, and the earlier the Gag-CASP8-VLPs were injected, the more profound the inhibition of tumor growth was observed. Overall, this study provides evidence of the feasibility of a novel antitumor strategy. Further optimization of the cancer cell-targeting specificity of this antitumor system may provide a potent anticancer therapy.
Footnotes
Acknowledgments
The authors thank Dr. S. M. Srinivasula (Thomas Jefferson University) for kindly providing CASP8-expressing plasmid. X-J.Y. is a recipient of the Manitoba Research Chair Award. This work was financially supported by Manitoba Research Chair Grant to X-j.Y, and grant from National Natural Science Foundation of China NNSFC 81460466 to Z-J.A.
Disclosure Statement
No competing financial interests exist.