
Abstract
Background:
Calbindin 1 (CALB1), a constituent Ca2+-binding protein, has been reported to prevent apoptotic death in tumor cells. However, the microRNA-mediated regulatory mechanism of CALB1 expression in nonsmall cell lung cancer (NSCLC) has not been reported so far.
Methods and Results:
In this study, CALB1 was found to be overexpressed in NSCLC tissues through the immunohistochemistry assay. Higher CALB1 expression levels were significantly associated with the tumor–node–metastasis (TNM) stage. Moreover, higher expression of CALB1 predicts poor survival in NSCLC patients using the Kaplan–Meier plotter online analysis. In addition, miR-454-3p was predicted to target CALB1 using a software algorithm, validated by the luciferase assay, and analyzed by quantitative polymerase chain reaction and Western blot. The authors further found that miR-454-3p was downregulated in NSCLC tissues and cell lines. Downregulation of CALB1 or upregulation of miR-454-3p significantly suppressed NSCLC cell proliferation and induced cell apoptosis as shown by CCK-8 and flow cytometry analysis, respectively.
Conclusions:
Our results suggest that CALB1 is a direct target of miR-454-3p and downregulation of CALB1 by miR-454-3p suppressed NSCLC cell functions, which may shed light on its potential application in NSCLC therapy.
Introduction
Lung cancer is one of the most frequently diagnosed cancers and leading causes of cancer-related deaths worldwide. 1 It was estimated that there were 234,030 new cases, leading to the death of ∼154,050 people in 2018 in United States. 2 Nonsmall cell lung cancer (NSCLC) represents ∼85% of all cases of lung cancer, which were classified into adenocarcinoma, squamous cell carcinoma, and large cell carcinoma. 3 Most patients have a relatively poor outcome due to limited treatment options, inadequate screening systems, and delayed appearance of clinical symptoms. 4 It is well known that the complex biology of cancer progression is a major obstacle in the treatment of NSCLC. Therefore, deep understanding of the mechanism of cancer growth and development may provide new ways for developing therapeutic agents.
Calbindin 1 (CALB1) is a vitamin-dependent calcium-binding protein with six putative EF hands and is located in human chromosome 8q21.3-q22.1. 5 CALB1, also known as calbindin-D28K, 6 is expressed in the intestine of birds, neurons, and endocrine cells of rats. It is detectable in mouse Purkinje cells and functions as a deciding factor for normal motor behavior, coordination, and sensory processing. 7 As a rapid endogenous calcium-buffering protein, CALB1 occurs in cellular Ca2+ influx and its depletion causes excess exocytosis. 8 Deficiency of CALB1 was also found to be associated with synaptic damage, neurodegeneration, apoptosis, and decrease in neurite growth and neuronal differentiation. 6 Interestingly, a recent study implicates CALB1 as an aberrantly dysregulated gene in several types of malignant cancers. Meng et al. 9 found that CALB1 expression was significantly decreased in endometrial cancer cells with overexpression of MTDH. In osteosarcoma, CALB1 knockdown caused a reduction in cell proliferation and colony formation. 10 A study by Watanabe et al. showed that most of the NSCLCs, which had a significantly high level of CALB1, were at the advanced cancer stage. 11 Notably, CALB1 was correlated with recurrence-free survival in postoperative NSCLC patients. 12
It has been suggested that over one-half of genes encoded in the human genome are regulated by microRNAs (miRNAs), which can bind to the target gene's 3′ untranslated region (UTR) to promote or depress transcripts through partial complementarity. 13,14 Emerging evidence has suggested the implication of miRNAs in progression and development of human cancer through cross talk with their target messenger RNAs (mRNAs). 15 Based on these evidences, the authors speculated that CALB1 might be regulated by certain miRNAs in NSCLC. On the other hand, miR-454-3p is a human cancer-related miRNA that overexpressed in diffuse large B-cell lymphoma, 16 glioma, 17 osteosarcoma, 10 and breast cancer, 18 but underexpressed in renal carcinoma. 19 Until now, it is reported to be associated with glioma prognosis and plays an important role in regulation of osteosarcoma cell growth and survival. 9,17,19 However, its biological function and its target gene are still undefined in NSCLC.
To validate speculation, the authors first determined the expression of CALB1 and miR-454-3p in NSCLC tissues and cell lines. Then, the authors assessed whether CALB1 is associated with clinicopathologic parameters, prognosis, and malignant behaviors of NSCLC. The cross talk between CALB1 and miR-454-3p is explored after screening the potential regulator of CALB1 by using miRNA-target gene databases. Hence, modulation of CALB1 through miR-454-3p may provide a possible therapeutic strategy for NSCLC therapy.
Materials and Methods
NSCLC tissue specimens and cell lines
A total of 40 paraffin-embedded NSCLC samples as well as fresh tumor tissue samples with paired noncancerous tissues were collected from 20 NSCLC patients in the Department of Thoracic Surgery, The First People's Hospital of Wenling (Wenling, China). Histopathological evaluation was independently performed by two pathologists. Some clinical information, including age, sex, and tumor–node–metastasis (TNM) stage, was recorded and summarized in Table 1. The clinical stage of tumors was evaluated according to the 7th edition of the American Joint Committee on Cancer (AJCC) TNM staging system. 20 Before surgery, all participants did not undergo any chemotherapy or radiotherapy and voluntarily signed an informed consent. The present study was approved by the Ethics Committee of The First People's Hospital of Wenling.
Association Between Calbindin 1 Expression and Clinicopathologic Parameters in Patients with Nonsmall Cell Lung Cancer
Bold value indicates statistically significant difference.
p < 0.05.
CALB1, Calbindin 1; TNM, tumor–node–metastasis.
Human NSCLC cell lines (H1299, 95D, A549, and SPC-A1) and the normal immortalized bronchial epithelial cell line (BEAS-2B) were purchased from the American Type Culture Collection (ATCC, Manassas, VA). All the NSCLC cell lines were cultured in RPMI-1640 medium with 10% fetal bovine serum (FBS; Gibco). The BEAS-2B cell line was cultivated in Dulbecco's modified Eagle's medium containing 10% FBS (Gibco). All cell lines were maintained in a humidified atmosphere containing 5% CO2 at 37°C.
Immunohistochemical analysis
Immunohistochemical (IHC) staining was performed using a standard streptavidin–biotin–peroxidase complex method, as described in a previous study. 21 In brief, paraffin-embedded sections (4 μm thick) were made and subsequently deparaffinized with xylene and rehydrated in gradient alcohol. After antigen retrieval and endogenous peroxidase blocking, sections were incubated with anti-CALB1 (1:200; Santa Cruz Biotechnology, Inc.) overnight at 4°C, followed by incubation with horseradish peroxidase (HRP)-conjugated secondary antibody for 30 min at 37°C. The expression of CALB1 was estimated based on the sum of the intensity and extent score of positive tumor cells, according to a previous report. 22 Finally, the expression of CALB1 was classified into two groups, including low (<3) and high (>3), based on the sum of intensity and extent score scaling from 0 to 6.
Kaplan–Meier overall survival analysis
The Kaplan–Meier plotter (
Protein extraction and Western blot analysis
Cells were collected and lysed with RIPA solution containing protease inhibitors (Beyotime, Beijing, China). Total proteins were extracted by centrifugation at 12,000 × g at 4°C for 20 min. The concentration of protein was determined using the BCA protein assay kit (Beyotime). Subsequently, equal amounts of protein were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto PVDF membranes. After blocking with 5% skimmed milk in Tris-buffered saline with Tween-20 for 2 h, the membranes were incubated with primary antibodies against CALB1 (1:500; Santa Cruz Biotechnology, Inc.) or GAPDH (1:5000; Proteintech) overnight at 4°C. Finally, the membranes were washed and incubated with secondary antibody for 2 h at room temperature. Protein bands were detected by enhanced chemiluminescence (ECL; Millipore, Darmstadt, Germany).
Oligonucleotide synthesis and cell transfection
miR-454-3p mimics (5′-UCCCUGAGACCCUUUAACCUGUGA-3′) and the corresponding negative control (miR-NC: 5′-CAGUACUUUUGUGUAGUACAA-3′) were chemically synthesized by GenePharma Co., Ltd. (Shanghai, China). Small interfering RNA for CALB1 (si-CALB1: 5′-CCTGGCGAGTATCATCATT-3′) and its NC (si-NC: 5′-UUCUCCGAACGUGUCACGUTT-3′) were purchased from RiboBio Co., Ltd. (Guangzhou, China). CALB1 silencing was accomplished by transfecting 95D and A549 cells with 10-nM si-CALB1 or si-NC for 48 h. To achieve miR-454-3p overexpression, 95D and A549 cells were transfected with 0.1 μM miR-454-3p mimics or miR-NC for 48 h. All transfection experiments were performed using Lipofectamine 2000 (Invitrogen, Waltham, MA) and repeated at least three times.
CCK-8 assay
After 48 h of transfection, cells were seeded in 96-well plates at a density of 3,000 cells per well and used for detection of proliferation by CCK-8 assay. After culturing for 0, 24, 48, 72, and 96 h, respectively, cells were washed with phosphate-buffered saline twice, followed by incubation in 10% CCK-8 solution (Boster, Wuhan, China) for 2 h at 37°C. The optical density (OD) value of each well was measured at 450 nm under a microplate reader and cell growth curves were plotted according to OD values. Each group was performed in triplicate.
Flow cytometry analysis
Cell apoptosis was detected using the Annexin V-APC/7-AAD double-labeling kit (KeyGEN, Nanjing, China) according to the manufacturer's instructions. Briefly, transfected cells were seeded into 6-cm dishes at a density of 80,000 cells per dish and culture was continued for 48 h. Then, cells were harvested by trypsinization and resuspended in 500 μL of binding buffer, followed by staining with Annexin V-APC/7-AAD for 30 min at room temperature in the dark. Finally, stained cells were analyzed using the FACSCalibur flow cytometer (BD Biosciences). Early apoptotic (Annexin V+/7-AAD−) and late apoptotic (Annexin V+/7-AAD+) cells were statistically analyzed and calculated from three repeated samples.
Prediction of potential miRNAs targeting CALB1
miRNAs targeting the expression of CALB1 were predicted using three miRNA-target gene databases (miRanda, TargetScan, and MicroCosm 5). One of the common predicted miRNAs was selected and identified as the potential miRNA targeting CALB1.
Dual-luciferase report assay
Dual-luciferase report assay was performed to confirm the direct interaction between miR-454-3p and CALB1. In brief, the pmirGLO-CALB1—3′UTR wild type (WT-CALB1 3′UTR) and pmirGLO-CALB1—3′UTR mutant (MUT-CALB1 3′UTR) were purchased from GenePharma Co., Ltd. For the luciferase reporter assay, 293T cells (1.5 × 10 5 ) were seeded in a 12-well plate and cotransfected with 300 ng DNA (WT-CALB1 3′UTR or MUT-CALB1 3′UTR) and 30 nM of either miR-454-3p mimics or miR-NC using transfection reagent Lipofectamine 2000 (Invitrogen), according to the manufacturer's instructions. After 48 h of transfection, cells were collected and luciferase activity was measured by the dual-luciferase reporter assay system (Promega Corporation, Madison, WI) following the manufacturer's instructions.
RNA extraction and quantitative real-time polymerase chain reaction
Total RNAs were extracted from tissues or cultured cells using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. Reverse transcription of miRNA was carried out using a TaqMan MicroRNA RT Kit (Applied Biosystems, Inc., Hangzhou, China). Expression of miR-454-3p was determined by the SYBR Green quantitative polymerase chain reaction kit. The quantitative real-time polymerase chain reaction (qRT-PCR) was performed with cycling conditions as follows: 5 min at 95°C, followed by 40 cycles at 95°C for 30 s and 65°C for 45 s. All of the stem-loop RT primers were purchased from RiboBio Co., Ltd., to detect miR-454-3p. Data were analyzed using the 2−ΔΔCq method. The relative expression of miR-454-3p was normalized to U6 as an endogenous control.
Statistical analyses
All data are expressed as means ± standard deviations of at least three repeated experiments. The chi-square (χ2) test was used to estimate the association between the expression level of CALB1 and clinicopathological factors. Overall survival curves were analyzed with the Kaplan–Meier method and compared using the log-rank test. Data were analyzed with Student's t-test or one-way analysis of variance. A value of p < 0.05 was regarded as statistically significant.
Results
CALB1 was highly expressed in NSCLC tissues
The expression of CALB1 in 40 paraffin-embedded NSCLC samples was evaluated using IHC analysis. Representative photomicrographs of different degrees of CALB1 expression intensity are shown in Figure 1A. As shown in Figure 1B, about 39.5% of paraffin-embedded NSCLC tissues showed weak staining of CALB1 protein, while 42.3% tissues showed moderate CALB1 staining, and 18.2% showed strong staining. The negative rate of CALB1 in adjacent tissues (69.0%) was evidently higher than that in cancerous samples (39.5%). Subsequent statistical analysis indicated that upregulation of CALB1 was positively associated with the advanced TNM stage (p = 0.003). However, CALB1 expression was not correlated with sex and age (p > 0.05, Table 1).
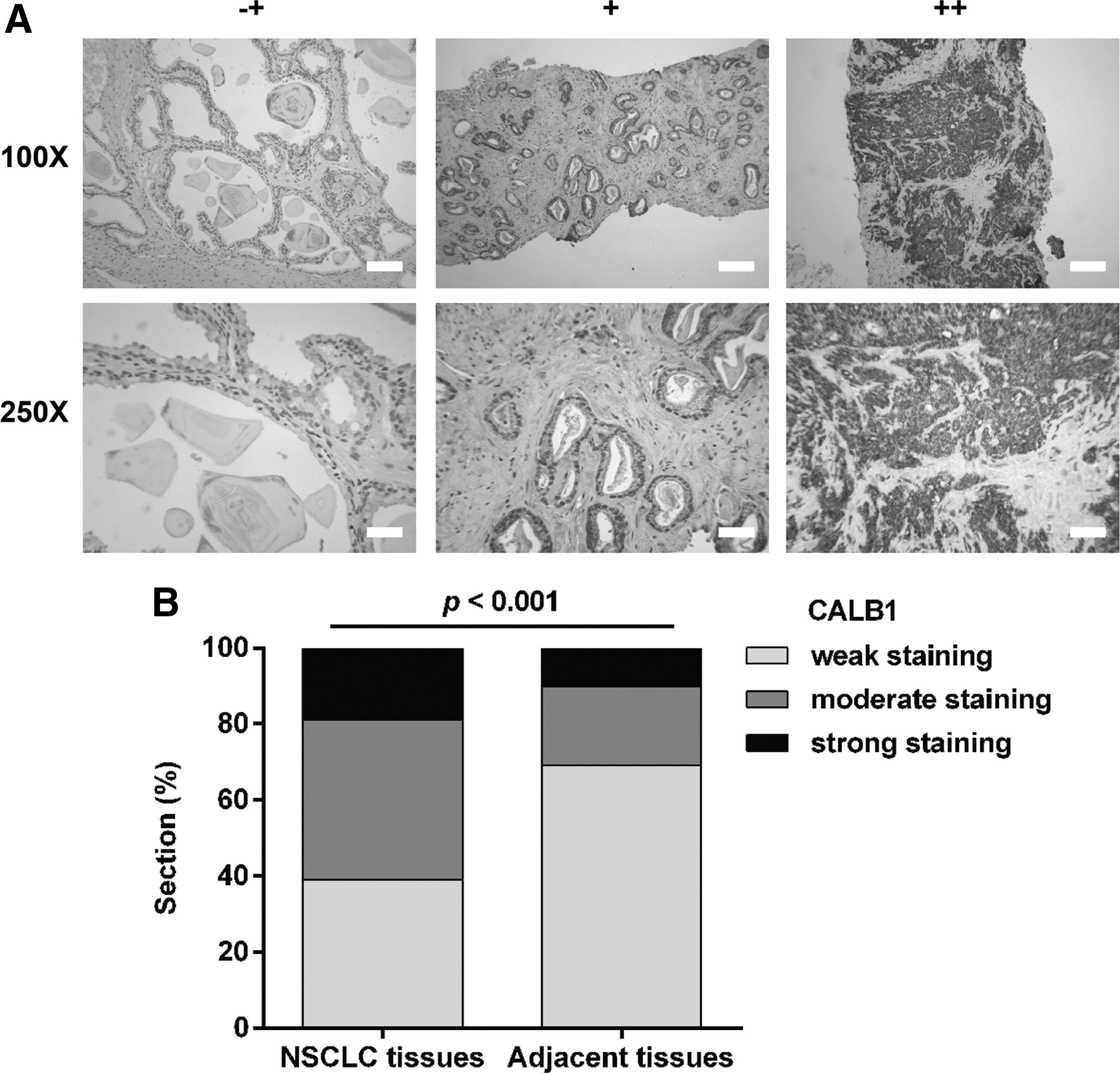
CALB1 expression in NSCLC tissues.
Higher expression of CALB1 predicts poor survival in NSCLC patients
Furthermore, the prognostic value of CALB1 in NSCLC patients was obtained from Kaplan–Meier plotter online software based on a public database (

CALB1 in NSCLC (Kaplan–Meier Plotter). Kaplan–Meier plots showing overall survival in NSCLC. In gray, patients with expression above the median, and in black, patients with expression below the median. The CALB1 gene and corresponding p-values are listed.
Downregulation of CALB1 suppressed cell proliferation and induced apoptosis in NSCLC cell lines
To investigate the biological function of CALB1 in NSCLC in vitro, the authors first explored CALB1 expression in NSCLC cell lines, and results revealed that CALB1 expression was higher in all four NSCLC cell lines compared with the normal, immortalized, bronchial epithelial cell line, BEAS-2B (Fig. 3A). Notably, 95D and A549 cells presented higher CALB1 levels, thus they were selected for transfection with si-CALB1. As shown in Figure 3B, western blot analysis confirmed that the expression of CALB1 was obviously downregulated in both 95D and A549 cells after si-CALB1 transfection. CCK-8 assay was then used to measure 95D and A549 cell proliferation, and results revealed that the knockdown of CALB1 significantly inhibited cell proliferation compared with the control groups (Fig. 3C, D, p < 0.001). Moreover, cell apoptosis assays were performed in the si-CALB1-transfected cells by flow cytometry. As shown in Figure 3E, the early apoptotic (22.5% ± 0.7% vs. 9.5% ± 0.2%) and late apoptotic (11.8% ± 0.3% vs. 3.4% ± 0.3%) percentages of cells in the si-CALB1 group were significantly higher compared with the si-NC group for 95D cells. Similar results were also found for A549 cells.

Downregulation of CALB1 affected cell proliferation and apoptosis in NSCLC cell lines.
CALB1 is a direct target of miR-454-3p
miRNAs, which suppress the expression of CALB1, were predicted using three miRNA-target gene databases (miRanda, TargetScan, and MicroCosm 5). After overlap analysis, miR-454-3p was identified as a biomarker in NSCLC using bioinformatic analysis, as previously reported by Shao et al. 23 Therefore, miR-454-3p was selected to examine whether miR-454-3p could inhibit the expression of CALB1 through binding to the 3′UTR of CALB1 mRNA. The predicted binding sites of miR-454-3p and CALB1 mRNA as well as mutant sequences of the MUT-CALB1 3′UTR plasmid are shown in Figure 4A. Subsequently, expression of miR-454-3p was determined in 20 pairs of fresh tumor tissue samples and noncancerous tissues. The results showed that expression of miR-454-3p was significantly downregulated in NSCLC tissues compared with adjacent tissues (Fig. 4B, p < 0.001). Consistently, qRT-PCR analysis also indicated that all the four NSCLC cell lines presented lower expression of miR-454-3p in comparison with BEAS-2B cells (Fig. 4C, p < 0.001). To further confirm that miR-454-3p directly interacts with the 3′UTR of CALB1 mRNA, dual-luciferase reporter assay was performed in 293T cells. As depicted in Figure 4D, the luciferase activity was significantly decreased in cells cotransfected with miR-454-3p mimics and WT CALB1 3′UTR luciferase reporter plasmid (p < 0.01), which was eliminated by transfection with the MUT CALB1 3′UTR luciferase reporter plasmid. Moreover, the regulatory effect of miR-454-3p on expression of CALB1 was assessed. As shown in Figure 4E, protein levels of CALB1 were significantly downregulated following overexpression of miR-454-3p in 95D and A549 cells. These findings suggested that the expression of CALB1 was negatively mediated by miR-454-3p in NSCLC cells.
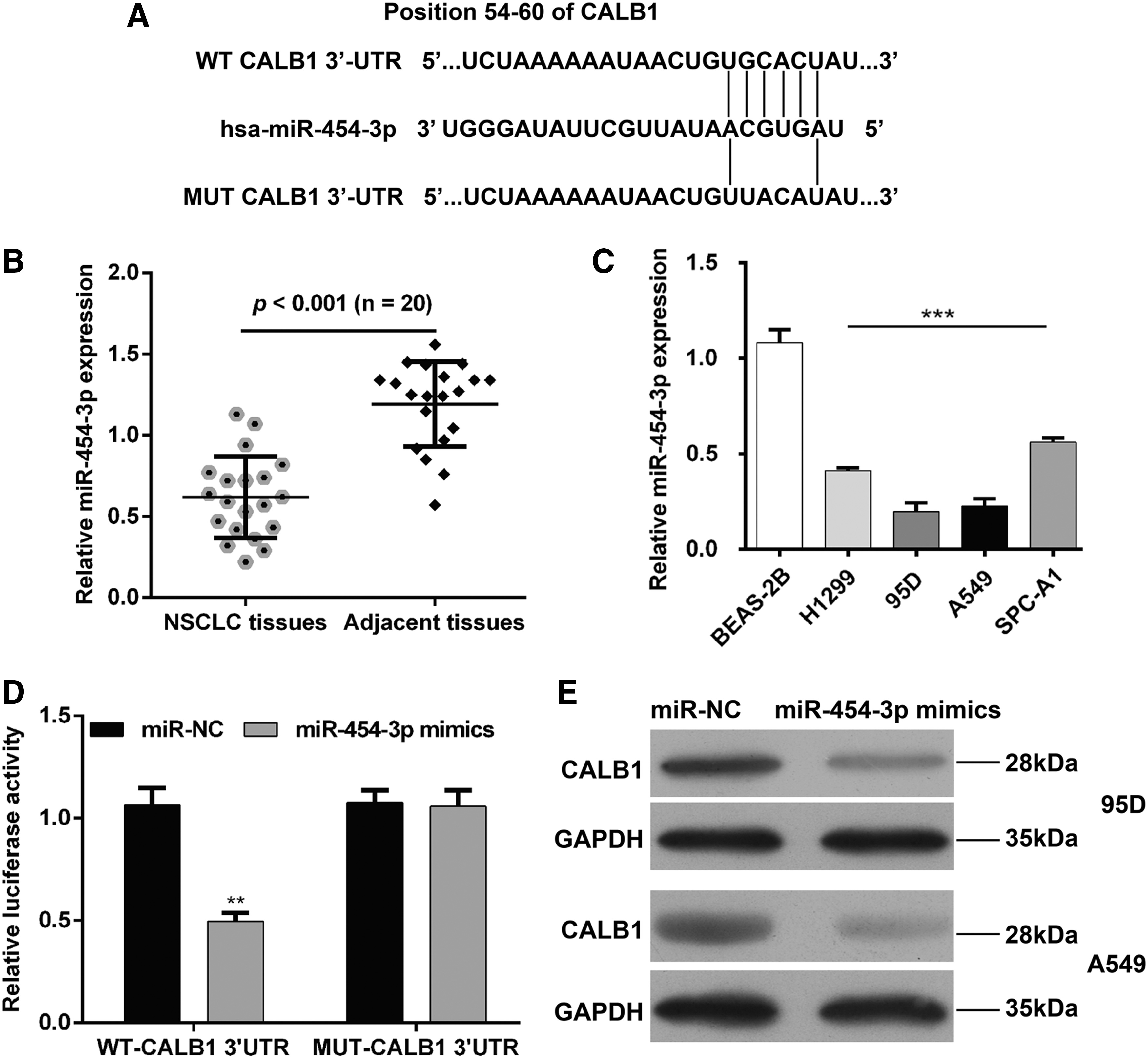
CALB1 is a target gene of miR-454-3p in NSCLC cells.
miR-454-3p overexpression inhibited cell proliferation and promoted apoptosis in NSCLC cell lines
As miR-454-3p expression was significantly downregulated in NSCLC tissues and cell lines, the authors speculated that it might be a tumor suppressor in NSCLC. To verify the hypothesis, gain-of-function assays were performed in NSCLC cells. First, 95D and A549 cells were transfected with miR-454-3p mimics and results showed that miR-454-3p levels were significantly increased compared with the control group, using qRT-PCR analysis (Fig. 5A, p < 0.001). CCK-8 assay showed that miR-454-3p overexpression significantly suppressed cell proliferation in (Fig. 5B, p < 0.001) 95D and (Fig. 5C, p < 0.001) A549 cells. Furthermore, overexpression of miR-454-3p caused a significant increase in early and late apoptotic rates in (Fig. 5D, p < 0.001) 95D and (Fig. 5E, p < 0.01, p < 0.001) A549 cells.
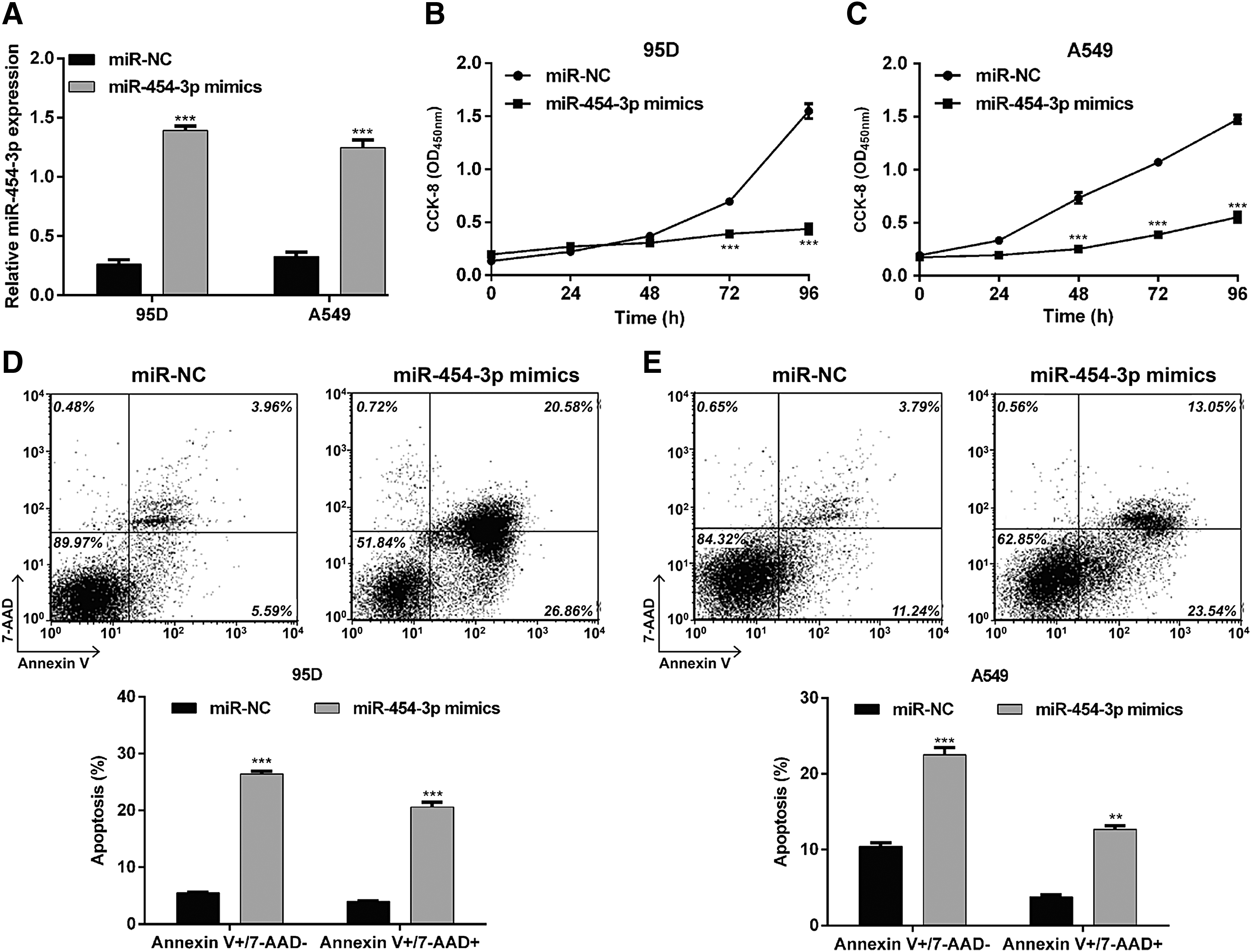
Upregulation of miR-454-3p affected cell proliferation and apoptosis in NSCLC cell lines.
Discussion
Calcium-binding proteins are involved in various physiological processes, including cell signaling, proliferation, apoptosis, differentiation, and memory. 24,25 CALB1 is expressed at higher levels in osteosarcoma cells and plays a key role in malignant biological behaviors. 10 Downregulation of CALB1 at the transcriptional and translational levels was observed in endometrial cancer cells with overexpression of MTDH compared with control cells. 9 However, the expression and physiologic function of CALB1 are unknown in NSCLC. In this present study, higher CALB1 levels were found to be associated with TNM stage and poor survival rate. It is well known that prognosis indicators could divulge information about patients' survival, disease progression, and metastatic spread. From this view, expression and clinical significance of CALB1 may help as a guide to appropriate treatment for NSCLC patients.
Based on the positive relationship between CALB1 expression and TNM stage, the authors hypothesize that CALB1 may act as an oncogene involved in NSCLC growth, which mainly influences proliferation and apoptosis. Up to now, Huang et al. 10 revealed that knockdown of CALB1 caused a reduction in osteosarcoma cell proliferation and an increase in apoptosis. Choi et al. 26 found that enhanced expression of CALB1 is tightly linked to the neuritis number and primary neuritis length, which is caused by uncontrolled cell division. Consistently, results showed that CALB1 induces proliferation and prevents apoptosis in NSCLC cells. The authors thus speculate that CALB1 was responsible for NSCLC cell proliferation, which might be ascribed to delayed cell apoptosis.
Interestingly, accumulating evidence implies that CALB1 has a cytoprotective role in protecting various cell types against apoptosis. 27 For example, CALB1 is reported to have a potential role in preventing MCF-7 cell apoptosis induced by 1,25(OH)2D3. 28 In dopaminergic neurons, CALB1 can activate the PI3-kinase-Akt signaling pathway, leading to inhibition of apoptosis. 29 In osteoblasts and osteocytes, it could suppress glucocorticoid-induced apoptosis. 30 In addition, controlling of Ca2+ signaling through Ca2+ buffers is linked to the process of apoptosis. 31,32 Sergeev 31 revealed that elevation of Ca2+ buffering caused by enhanced expression of CALB1 could block Ca2+-mediated apoptosis in breast cancer cells. The authors thus guess that a similar mechanism may operate in both early and late apoptosis in NSCLC cells with depletion of CALB1, but biological contexts of the downstream signaling mediator still need further investigation.
Furthermore, the authors identified the miRNAs regulating CALB1 using three miRNA-target gene databases. The overlap analysis revealed that CALB1 might be a direct target of miR-454-3p. Further investigation confirmed that miR-454-3p downregulated CALB1 by binding its 3′UTR in NSCLC cells. Actually, miR-454-3p was reported to be a potential prognostic biomarker for human glioma, which plays an important role in reducing cellular radioresistance of renal carcinoma cells. 17,19 Some researchers have demonstrated that STAT3, ATG12, BTG1, and c-Met are direct targets of miR-454-3p. 19,33,34 In this study, CALB1, a member of calcium-binding proteins, is a novel molecular target of miR-454-3p in NSCLC. Of note, enhanced expression of miR-454-3p could imitate the effects of CALB1 knockdown in inhibiting proliferation and promoting apoptosis, indicating that CALB1 is negatively regulated by miR-454-3p and contributes to NSCLC cell growth and survival.
In summary, this study revealed that miR-454-3p negatively regulates CALB1, a gene expressed at high levels in NSCLC tissues and cell lines. CALB1 is associated with the TNM stage and poor prognosis in NSCLC. It is also a potential regulator in cancer cell proliferation and survival. Based on these investigations, targeted CALB1 by miR-454-3p may provide a promising therapeutic target for NSCLC treatment.
Footnotes
Acknowledgment
The study was supported by The First People's Hospital of Wenling.
Disclosure Statement
The authors declare that they have no competing interests.