
Abstract
Background:
Circular RNAs (circRNAs) comprise a class of noncoding RNA molecules that play an important role in several normal cellular functions, as well as tumorigenesis in humans. However, the expression patterns and biological functions of circRNAs in ovarian cancer (OC) remain unclear.
Methods:
Therefore, we investigated the expression profiles and biological functions of certain circRNAs in OC tumor tissues. The expression of three circRNAs (VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013) was detected by real-time polymerase chain reaction in OC cell lines, and also in tumor and pericarcinous tissues obtained from 20 patients with OC. The function of VPS13C-has-circ-001567 in SKOV3 and OV-1063 cells was investigated by knockdown of VPS13C-has-circ-001567 and then analyzing any resultant effects on the cell cycle, cell proliferation, apoptosis, and cell invasion ability. E-cadherin and N-cadherin expressions were analyzed by immunofluorescence and western blotting. Finally, the tumorigenicity of OC cells was assessed in nude mice.
Results:
The results showed that VPS13C-has-circ-001567 was expressed at significantly higher levels in OC tumor tissues compared with pericarcinous tissues, and this overexpression was associated with tumor node metastasis stage and lymph node metastasis. We found that knockdown of VPS13C-has-circ-001567 significantly promoted apoptosis and inhibited the proliferation of SKOV3 and OV-1063 cells in vitro. Knockdown of VPS13C-has-circ-001567 led to cell cycle arrest at G1 phase and decreased the percentage of S1 phase cells. Additionally, knockdown of VPS13C-has-circ-001567 decreased the invasion ability of SKOV3 and OV-1063 cells, and also changed the levels of E-cadherin and N-cadherin expressions. Knockdown of VPS13C-has-circ-001567 significantly reduced the tumorigenicity of OC cells.
Conclusions:
Taken together, our results suggest that VPS13C-has-circ-001567 plays a role in the development of OC and might be a prognostic marker and therapeutic target for OC.
Introduction
Ovarian cancer (OC) is one of the most common gynecological cancers, 1,2 with an overall 5-year survival rate of <40%. Surgery followed by chemotherapy using platinum-based compounds such as carboplatin, cisplatin, or oxaliplatin is the standard therapy for OC. 3,4 However, patients with advanced OC have a high mortality rate because their tumors eventually become resistant to drug treatment. Thus, understanding the molecular mechanisms involved in OC pathology and discovering new therapeutic targets are critically important for improving the 5-year survival rate of OC patients.
Circular RNAs (circRNAs) comprise a class of noncoding RNAs, 5,6 which are reported to play an important role in several cellular activities. 7 –9 circRNAs can be generated by backsplicing of exons, introns, or both. 9 circRNAs are abundantly found in the cytoplasm, because their covalently closed continuous loop structure prevents their degradation by RNA exonucleases. 9 It has been proposed that circRNAs may have important regulatory roles. Moreover, recent evidence suggests that circRNAs act as miRNA sponges, 10 and the expression profiles of circRNAs are highly specific for certain tissues and pathological conditions. 11,12 Although the exact mechanism by which circRNAs might regulate genes remains to be clarified, circRNAs can potentially be used as disease biomarkers and novel therapeutic targets.
Recently evidence suggests that circRNAs might participate in the development and progression of cancer. 13 –15 Salzman et al. 11 first identified circRNAs in cancer and noncancer cell lines, as well as in tissue samples obtained from patients with acute lymphoblastic leukemia. 11 Dysregulation of circRNA was found in several human cancers, including bladder cancer, breast cancer, colorectal cancer, hepatocellular carcinoma (HCC), gastric cancer, kidney clear cell carcinoma, and prostate adenocarcinoma. 16 Recent information of how circRNAs might function in carcinogenesis and affect the malignant behavior of tumors has provided new insights into the molecular mechanism of cancer. However, the role played by circRNAs in OC has received little attention.
In this study, we screened three circRNAs and investigated their potential roles in OC, to demonstrate the unique role of those circRNAs in the carcinogenesis process leading to OC, as well as their relationship to the cellular behaviors of OC cells. An improved understanding of circRNAs may enable their use as targets for treating cancer, and lead to novel transcription-based approaches for suppressing or regulating the malignant behavior of cancer cells.
Materials and Methods
Collection of tissue specimens
This study enrolled 20 OC patients who were surgically treated at the People's Hospital of Xiangshui County, Jiangsu Province, China. Specimens of their cancer tissue and pericarcinous tissue (adjacent to the tumor with a distance >3 cm) were collected during surgery. Each patient provided their written informed consent prior to study enrollment, and the study protocol was approved by the Research Ethics Committee of the People's Hospital of Xiangshui County. Immunohistochemical studies of the tumor samples and pericancerous samples were performed by a certified pathologist, and the results were audited by two independent pathologists to confirm a diagnosis of OC. None of the patients had received a blood transfusion, chemotherapy, or radiotherapy before surgery. All tissue samples were immediately frozen and then stored in liquid nitrogen for use in subsequent experiments.
Cell culture
Five human OC cell lines (IOSE80, ES-2, SKOV3, Caov-3, and OV-1063) were purchased from American Type Culture Collection (ATCC, Rockville, MD). The cells were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 mg/mL streptomycin in an incubator with 5% CO2 at 37°C.
A lentivirus vector encoding shRNA targeting has-circRNA-001567 (shRNA) and another lentivirus vector encoding a scrambled sequence shRNA (negative control, NC) were constructed using a vector purchased from Genechem Biotechnology, Inc., Ltd. (Shanghai, China).
SKOV3 and OV-1063 cells were transfected with high titers of shRNA lentivirus (2 × 10 8 TU/mL) or a negative control lentivirus expression vector (all with a MOI of 30) according to manufacturer's instructions. SKOV3 and OV-1063 cells with stable shRNA or NC expression were obtained by continuous treatment with 2 μg/mL puromycin.
RNA extraction and reverse transcription-polymerase chain reaction
Total RNA was extracted from tumor samples, pericarcinous tissue samples, and harvested cells using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA) according to manufacturer's instructions. For mRNA analysis, Oligo (dT) was used as a primer to reverse-transcribe 1.0 μg of total RNA into cDNA using a reverse transcription kit (Qiagen, Hilden, Germany). Three real-time polymerase chain reaction (RT-PCR) analyses were conducted for each sample. A 1:4 dilution of cDNA served as a template, and the PCR products were quantified using a Quantitect SyBr green PCR system (Qiagen) on a Rotorgene 6000 series PCR machine (Qiagen). Data were analyzed using Rotorgene software that was matched with the PCR machine. The following primers were used for VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013, respectively: forward 5′-TATAATTTTGTCTGCTTCATTTA-3′ and reverse 5′-TTAACACAGTCTAAAGTCTCAGAA-3′; forward 5′-AAAGAAAAGATAAATGAAGATATG-3′ and reverse 5′-CCTGTATAAGCTTACGATTCA-3′; forward 5′-ATTACTAAAGGCAAACGGTGAA-3′ and reverse 5′-GCTGGAATGCCTGTCCGT-3′. Relative expression levels were calculated using Rotorgene software. All mRNA quantification data were normalized to GAPDH expression and calculated using the 2ΔΔCt method.
CCK-8 and Edu assays
To investigate the effect of VPS13C-has-circ-001567 knockdown on cell proliferation, cells were cultured for 72 h in a 96-well plate that contained DMEM supplemented with 10% FBS and 100 mg/mL penicillin/streptomycin. Cell proliferation rates were detected with a Cell Counting Kit-8 (CCK-8; Roche) according to manufacturer's instructions. The Edu assay was carried out as previously described. 17
Cell cycle assay
SKOV3 and OV-1063 cells were transfected as previously described. At 48 h after transfection, the cells were collected and washed with cold phosphate buffered saline (PBS), after which they were resuspended in 200 μL PBS and fixed in 800 μL 100% ethanol at 4°C for 24 h. The fixed cells were then stained with PI (100 μg/mL; Sigma) and treated with RNase (50 μg/mL; Sigma) in PBS at 37°C for 20 min in the dark. The cell cycle distribution was analyzed using a flow cytometry provided with the Cell-Quest software.
Cell proliferation and apoptosis assays
SKOV3 and OV-1063 cells were pre-labeled with PI and annexin V-FITC and then examined for apoptosis using an apoptosis detection kit (Invitrogen, Burlington, Canada) according to manufacturer's instructions. The results was analyzed using a flow cytometry provided with the Cell-Quest software (Becton Dickinson, San Jose, CA).
Immunohistochemistry, immunofluorescence, and western blotting
The expression levels of various proteins in OC cells and cancer tissue specimens were evaluated by western blotting, immunohistochemistry, and immunofluorescence as previously described. 18 The primary antibodies used in the studies were anti-E-cadherin (Abcam, Cambridge, MA), anti-N-cadherin (Abcam), and anti-GAPDH (Abcam). GAPDH was used as an internal standard during western blotting. Three different biological samples were independently analyzed by each assay method.
Transwell invasion and tumorigenicity assays
To assess the invasion ability of transfected SKOV3 and OV-1063 cells, aliquots of cells (2.5 × 10 4 ) were seeded into the upper wells of a Transwell plate. The wells had been precoated with 100 μL of growth factor-reduced Matrigel (BD Biosciences) diluted in PBS (1:45). The cells were then incubated in serum-free medium for 24 h. After incubation, the cells in the upper wells were removed and the wells were washed and fixed with 4% paraformaldehyde, after which the cells were stained with 0.1% crystal violet (Sigma Aldrich) and air-dried. The tumorigenicity of OC cells was analyzed by injecting them into nude mice as previously described, 19 and the tumor volumes were recorded. Three separate experiments were performed and the average number of invaded cells was calculated.
Statistical analysis
All statistical analyses were performed using IBM SPSS Statistics for Windows for the Social Sciences, version 19.0 software (IBM Corp., Armonk, NY). Results are presented as mean ± SEM of values obtained from three separate experiments. The statistical significance of standardized expression data was determined by paired or unpaired Student's t-test. Differences with a p-value ≤0.05 were considered statistically significant.
Results
VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013 expression was increased in OC tissues
To determine the role of circRNA in OC, we used RT-PCR to evaluate VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013 expression in the tumor and pericarcinous tissues of patients with OC. Our results showed that the levels of VPS13C-has-circ-001567 (p < 0.01; Fig. 1A), RAD50-has-circ-00718 (p < 0.01; Fig. 1B), and SPECC1-has-circ-000013 (p < 0.05; Fig. 1C) expressions in OC tumor tissues were significantly higher than those in the pericarcinous tissues. Among the three candidate circRNAs investigated, VPS13C-has-circ-001567 was the most highly expressed in tumor tissues. Previous studies have demonstrated that circRNAs play a role in the tumorigenesis of human malignancies. Therefore, we hypothesized that VPS13C-has-circ-001567 may have an important role in OC.
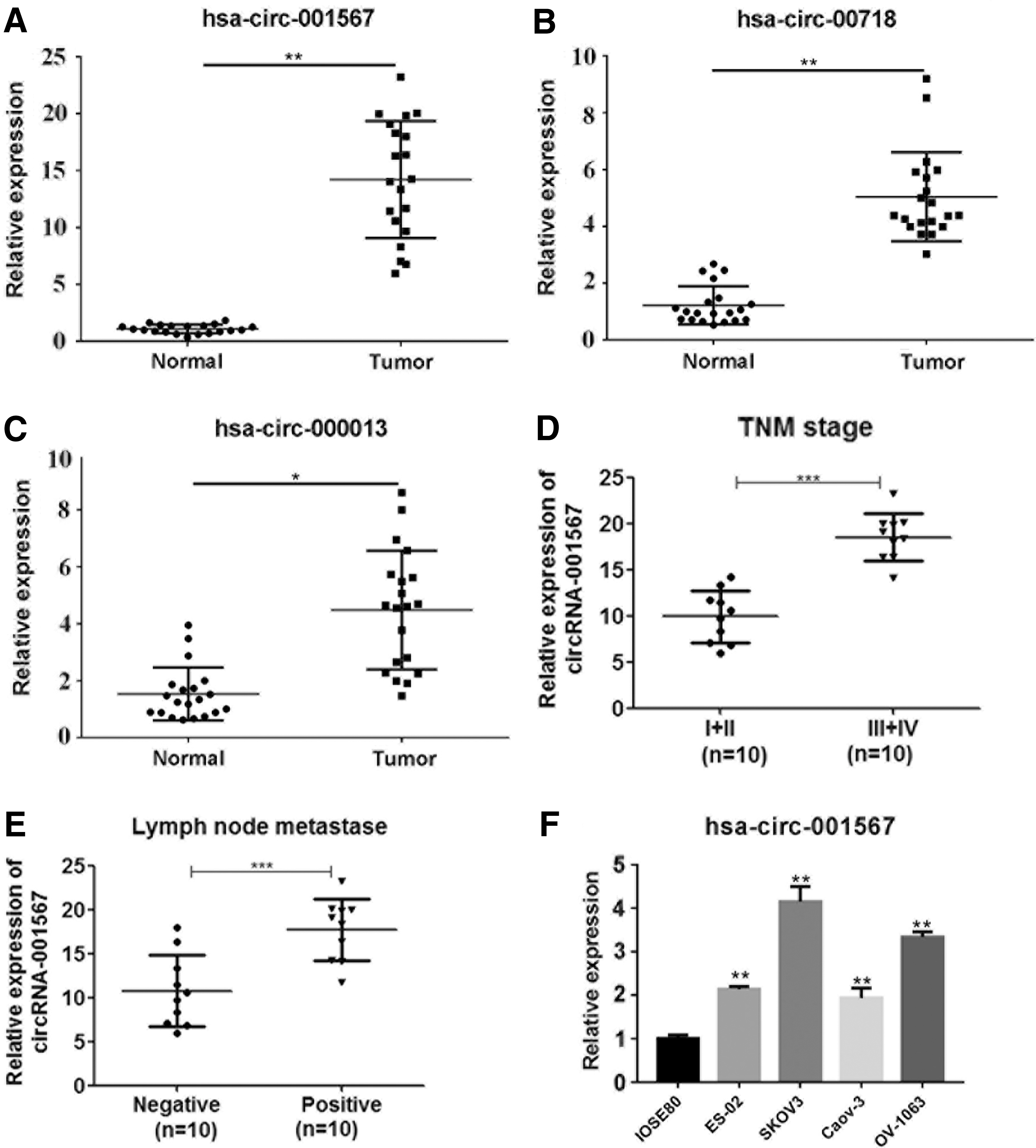
Increased VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013 expressions in OC tissues and cell lines. The relative expression levels of VPS13C-has-circ-001567
We next investigated VPS13C-has-circ-001567 expression in patients with different tumor node metastasis (TNM) stages of OC and found that the levels of VPS13C-has-circ-001567 expression in later-stage OC patients were significantly higher than those in early-stage patients (Fig. 1D). Moreover, patients with lymph node metastasis had higher levels of VPS13C-has-circ-001567 expression than patients without lymph node metastasis (Fig. 1E). To further investigate the role played by VPS13C-has-circ-001567 in the development of OC, we determined the levels of VPS13C-has-circ-001567 expression in five different cell lines (IOSE80, ES-2, SKOV3, Caov-3, and OV-1063) and found significantly higher levels of VPS13C-has-circ-001567 expression in SKOV3 and OV-1063 cells and the lowest levels in IOSE80 cells, which is a normal ovarian cell line (Fig. 1F). This finding was consistent with our results from analyzing patient tissue specimens. Therefore, we used the SKOV3 and OV-1063 cell lines in our subsequent experiments.
Effect of knockdown of VPS13C-has-circ-001567 on the proliferation of SKOV3 and OV-1063 cells
To inhibit the expression of VPS13C-has-circ-001567, we used a lentivirus system to transfect designed shRNA targeting VPS13C-has-circ-001567 into the high VPS13C-has-circ-001567 expression cell lines SKOV3 and OV-1063. We then performed RT-PCR to evaluate the levels of VPS13C-has-circ-001567 expression in SKOV3 and OV-1063 cells at 48 h after transfection. The levels of VPS13C-has-circ-001567 in cells transfected with recombinant lentivirus were significantly decreased at 48 h after transfection in both SKOV3 (Fig. 2A) and OV-1063 (Fig. 2B) cells, compared with levels in cells transfected with scrambled shRNA. CCK-8 assay results showed that knockdown of VPS13C-has-circ-001567 in SKOV3 (Fig. 2C) and OV-1063 (Fig. 2D) cells caused a significant decrease in cell proliferation. To further verify those results, the cell proliferation rates were also analyzed by the Edu assay. We observed that the percentage of Edu-positive cells (M2 peak) was remarkably decreased by knockdown of VPS13C-has-circ-001567 expression in both SKOV3 (Fig. 2E) and OV-1063 cells (Fig. 2F). These results indicated that a lentivirus-mediated knockdown effectively and specifically decreased the levels of VPS13C-has-circ-001567 in SKOV3 and OV-1063 cells, and a stable knockdown of VPS13C-has-circ-001567 caused a reduction of cell proliferation.
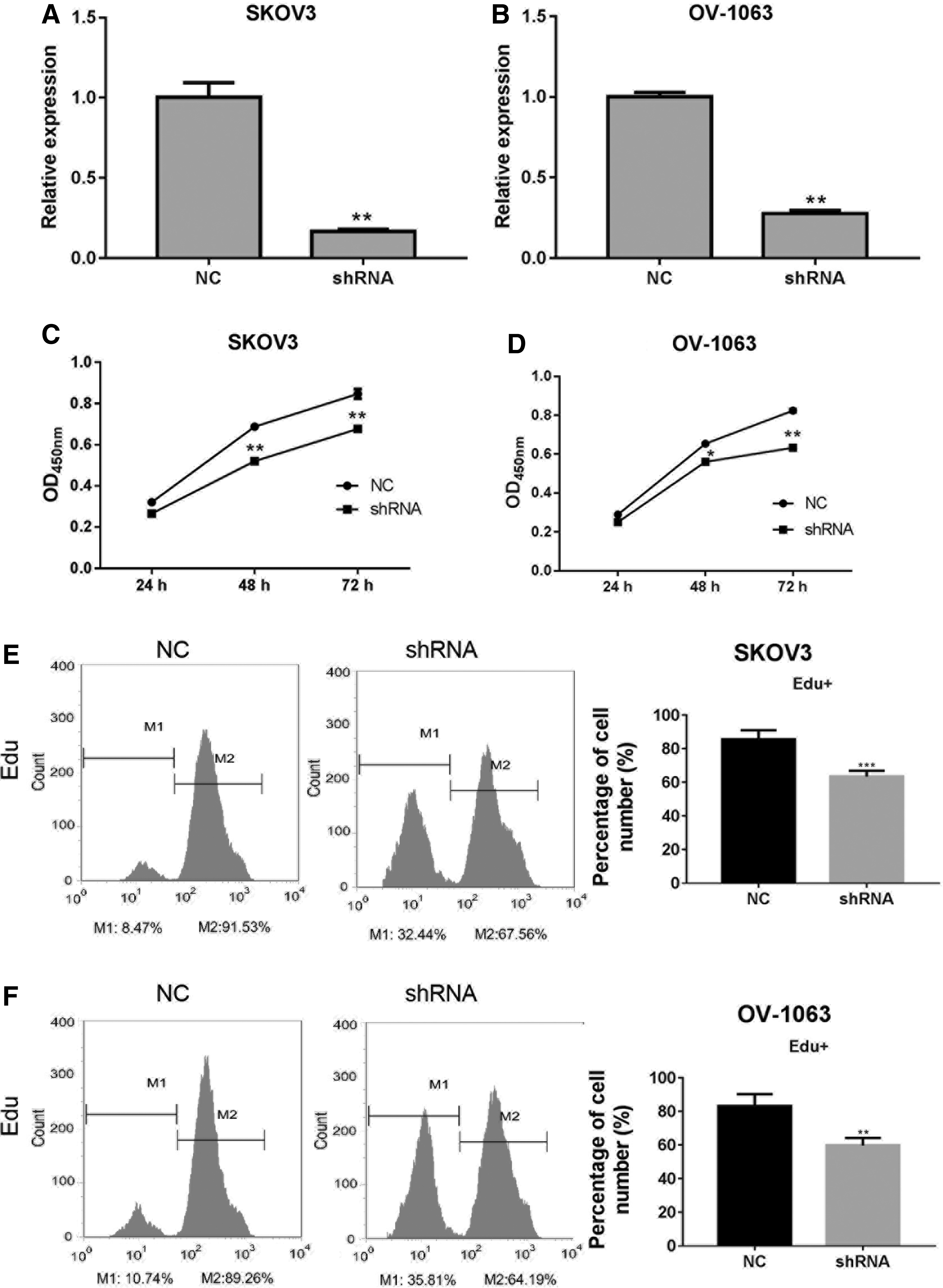
Effect of VPS13C-has-circ-001567 knockdown on SKOV3 and OV-1063 cell proliferation. Relative expression of VPS13C-has-circ-001567 in SKOV3
Effects of VPS13C-has-circ-001567 knockdown on cell apoptosis and cell cycle progression
We next assessed the phenotypic consequences of VPS13C-has-circ-001567 knockdown in SKOV3 and OV-1063 cells. Knockdown of VPS13C-has-circ-001567 induced a significant increase in the percentages of apoptotic SKOV3 (Fig. 3A) and OV-1063 (Fig. 3B) cells. The effect of VPS13C-has-circ-001567 knockdown on the cell cycle was examined through PI staining followed by a FACS analysis. As shown in Figure 3, there were significant increases in the numbers of G1 phase SKOV3 (Fig. 3C) and OV-1063 (Fig. 3D) cells, compared with the number of G1 phase normal control cells. The knockdown cells also showed corresponding decreases in their S phase populations, while the number of G2 phase cells remained unchanged. These results indicated that VPS13C-has-circ-001567 knockdown inhibited the proliferation of SKOV3 and OV-1063 cells by inducing G1 phase arrest and apoptosis.

Effects of VPS13C-has-circ-001567 knockdown on OC cell apoptosis and cell cycle progression.
Effect of VPS13C-has-circ-001567 knockdown on cell migration and invasion
We used the Transwell assay to evaluate the effect of VPS13C-has-circ-001567 knockdown on the migratory and invasive properties of SKOV3 and OV-1063 cells. As expected, knockdown of VPS13C-has-circ-001567 significantly decreased the invasion ability of SKOV3 (Fig. 4A, B) and OV-1063 (Fig. 4C, D) cells, as the numbers of migrated and invaded cells in the knockdown groups of both cell lines were significantly lower than those in the NC groups (p < 0.01). To provide molecular evidence supporting this effect, we examined the expression of epithelial-mesenchymal transition (EMT) biomarkers that play a key role in cell migration and invasion processes. Our immunofluorescence assays revealed that the expression of E-cadherin was greatly increased by knockdown of VPS13C-has-circ-001567 expression in both SKOV3 and OV-1063 cells (Fig. 4E); however, the levels of E-cadherin showed a completely opposite change in cells with already decreased levels of VPS13C-has-circ-001567 (Fig. 4E). Moreover, our western blot analysis revealed changes in E- and N-cadherin protein expression after knockdown of VPS13C-has-circ-001567, which were consistent with the immunofluorescence results (Fig. 4F). These findings suggest that VPS13C-has-circ-001567 is involved in the EMT of OC cells.

Effects of VPS13C-has-circ-001567 knockdown on the migration and invasion abilities of SKOV3 and OV-1063 cells.
Effect of VPS13C-has-circ-001567 knockdown on cell tumorigenicity
To further validate the role of VPS13C-has-circ-001567 expression in OC development, the tumorigenicity of SKOV3 cells was analyzed by injecting them into nude mice. We found that the tumor volumes in mice injected with shRNA-transfected cells were significantly lower than those in mice injected with negative control cells (Fig. 5A, B). Immunohistochemistry results revealed that the levels of E-cadherin in the cancerous tissues of mice injected with VPS13C-has-circ-001567 knockdown cells were greatly elevated compared with those levels in mice injected with negative control cells (Fig. 5C). In contrast, the N-cadherin protein levels in tumors that developed from VPS13C-has-circ-001567 knockdown cancer cells were significantly decreased compared with the negative controls (Fig. 5C). These results show that VPS13C-has-circ-001567 plays an important role in OC tumorigenesis.

Knockdown of VPS13C-has-circ-001567 decreased the tumorigenicity of SKOV3 and OV-1063 cells.
Discussion
In this study, we investigated the potential role of three circRNAs (VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013) in OC. We found significantly increased expression of the three circRNAs in samples of tumor tissue compared with pericarcinous tissues, and VPS13C-has-circ-001567 was the circRNA most highly expressed in the tumor tissues. VPS13C-has-circ-001567 expression was also positively correlated with the OC TNM stage and lymph node metastasis. Our detection of VPS13C-has-circ-001567 expression in both normal ovarian cell lines and OC cell lines confirmed a significant upregulation of VPS13C-has-circ-001567 in OC cell lines SKOV3 and OV-1063, compared with the normal ovarian cell line, IOSE80. Next, we further investigated the effect of VPS13C-has-circ-001567 knockdown on the apoptosis, cell cycle, and proliferation of SKOV3 and OV1063 cells. Our results indicated that knockdown of VPS13C-has-circ-001567 induced reductions in cell proliferation and proportion of S1 cells, and also promoted cell apoptosis and decreased the tumorigenicity of cells in nude mice. These results suggest that the circRNAs VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013 participate in OC development by regulating the biological behaviors of cancer cells.
In this study, we found an upregulation of VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013 in OC cells and tissues. The dysregulation of circRNAs has previously been reported. Dysregulation of has-circ-002059 was reported in patients with gastric cancer 20 and found to be associated with clinical and pathological features such as tumor stage, distant metastases, gender, and age. 21 In patients with KRAS-mutated CRC, a downregulation of certain circRNAs was found in their colorectal tissues, compared with their normal tissues, 22 as well as in KRAS-mutant colon cancer cell lines. 23 circRNAs have also been found to be involved in other cancers such as bladder cancer. For example, circTCF25, circZFR, circPTK2, and circBC048201 were found to be significantly upregulated in bladder tumor tissues compared with adjacent nontumor tissues. 24 Moreover, certain circRNAs, such as has-circ-0005075 and ciRS-7, have been found to be upregulated in HCC tissue. 25 Our study provides new evidence for the role of circRNAs in cancers.
In this study, we investigated the underlying mechanism by which VPS13C-has-circ-001567 influences cancer development and found evidence that increased expression of VPS13C-has-circ-001567 may promote cell proliferation and invasion. circRNAs can affect cancers through various methods, and they serving as miRNA sponges is the most studied method. 26 –28 It also has been reported that circRNAs can transcriptionally or post-transcriptionally regulate gene expression by interacting with miRNAs. 10,14 For example, increasing evidence suggests that cirs-7 regulates miR-7 expression and acts as an oncogene partly through targeting miR-7 in cancer cells. miR-7 can directly regulate the expression of several important genes, including EGFR, IRS-1, PIK3CD, and mTOR. 29 circ-TCF-25 was reported to act as a miRNA sponge, by decreasing the levels of miR-103a-3p and miR-107 in cancerous tissues; this activity is associated with the development of cancer. 30 In our study, increased expression of VPS13C-has-circ-001567 in OC cells promoted cell invasion and proliferation, and a miRNA-dependent regulatory mechanism may be responsible for these effects. Further studies focusing on identifying the target miRNAs of VPS13C-has-circ-001567, RAD50-has-circ-00718, and SPECC1-has-circ-000013 should provide more information.
In summary, our study results provide new evidence that circRNAs are involved in the development of OC and suggest that targeting the circRNA VPS13C-has-circ-001567 may be a novel approach for overcoming therapy-related resistance.
Footnotes
Acknowledgment
This study was supported by the People's Hospital of Xiangshui County.
Disclosure Statement
No competing financial interests exist.