
Abstract
Background:
The plant homeodomain finger 6 (PHF6) was originally identified as single gene mutated in Börjeson-Forssman-Lehmann syndrome, which was reported to be a tumor suppressor in T-cell acute lymphoblastic leukemia. However, the biological function of PHF6 in hepatocellular carcinoma (HCC) has been poorly characterized.
Materials and Methods:
In this study, we first determined the mRNA levels of PHF6 in HCC tissues and adjacent normal tissues using quantitative real-time PCR. Then the expression of PHF6 was knocked down in HCC cell lines (HepG2, SMMC-7721, and Bel-7402) by siRNA transfection. A series of functional experiments, including EdU proliferation assay, colony formation assay, and Transwell assay, were performed in HCC cells. Western blot analysis was used to detect the expression of PHF6, E-cadherin, and Vimentin.
Results:
We found that PHF6 was significantly elevated in HCC tissues and positively correlated with TNM stage, differentiation, and lymph node metastasis. Silencing PHF6 significantly inhibited cell proliferation, colony formation, and migration in HCC cells. Furthermore, silencing PHF6 obviously increased E-cadherin and decreased Vimentin expression.
Conclusions:
These findings suggest that PHF6 plays a positive role in the growth of HCC cells, and targeting PHF6 could serve as a promising therapeutic strategy for human HCC.
Introduction
Hepatocellular carcinoma (HCC) ranks the second most common cause of cancer-related mortality worldwide and accounts for 85%–90% of primary liver malignancy. 1,2 Accumulating data indicate that hepatocarcinogenesis is mainly ascribed to chronic hepatitis B, chronic hepatitis C, and other hereditary diseases. 3 Currently, liver resection and transplantation could prolong survival time for early-stage HCC patients, but the prognosis of HCC patients at advanced stage remains unsatisfactory due to its high recurrence and poor prognosis. 4 Currently, molecular-targeted cancer therapy has exhibited clinical significance to the management of HCC, which requires detecting many molecular markers related to tumorigenesis. 5,6 Therefore, it is urgently needed to identify a novel therapeutic target involved in HCC progression.
Plant homeodomain finger protein 6 (PHF6), located on Xq26-q27, was initially reported as a single gene mutated in Börjeson-Forssman-Lehmann syndrome characterized by X-linked intellectual disability. 7 PHF6 encodes a plant homeodomain (PHD) factor containing four nuclear localization signals and two imperfect PHD zinc finger domains, which could regulate gene transcription. 8 PHF6 might play a crucial role in human brain development for its higher expression in fetal and developing embryonic central nervous system. 9 Related studies have shown that inactivating mutations in PHF6 were frequently detected in adult and pediatric patients with T-cell acute lymphoblastic leukemia (T-ALL) or adults with acute myeloid leukemia, 10,11 highlighting its role as a tumor suppressor gene in these malignancies. On the contrary, PHF6 has been described to have a tumor-promoting role in a murine model of BCR–ABL1+ B-cell acute lymphoblastic leukemia (B-ALL). 12,13 Thus, these evidence suggests that PHF6 can act as a tumor suppressor or an oncogene in a lineage-dependent manner. Interestingly, somatic mutation of PHF6 gene has been recently identified in HCC. 14 However, the role of PHF6 and its underlying molecular mechanisms in HCC remain largely unknown.
In this study, we first examined the expression of PHF6 in human HCC tissues and cell lines, and found that it was overexpressed in HCC. Furthermore, we focused on elucidating the cellular functions of PHF6 by performing loss-of-function assays in HCC cells. Our findings may provide insight into the pathogenesis of HCC and identify PHF6 as a potential therapeutic target for patients with HCC.
Materials and Methods
Tissue specimens
In total 30 pairs of HCC tissues and matched adjacent noncancerous liver tissues were obtained from HCC patients at the Nanfang Hospital Affiliated to Southern Medical University. Before surgery, all patients provided written informed consent and did not receive any radiotherapy or chemotherapy treatment. According to the TNM classification system proposed by the 2010 International Union Against Cancer, 30 HCC tissues were classified into I+II (n = 15) and III+IV (n = 15). Pathological diagnosis, including tissue differentiation (9 well, 11 moderate, and 10 poor) and lymph node metastasis (16 No and 14 Yes), was based on the World Health Organization criteria. This study was approved by the Ethics Committee of the Nanfang Hospital Affiliated to Southern Medical University.
Cell culture and transfection
HCC cell lines (human hepatoblastoma HepG2, SMMC-7721, Bel-7402, and Huh-7) and normal liver cell line HL7702 were purchased from the Cell Bank of Chinese Academy of Sciences (Shanghai, China), which were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) with 10% fetal bovine serum (FBS; HyClone) and maintained in an incubator at 37°C with 5% CO2.
The small interfering RNA against PHF6 (siPHF6) was designed and synthesized by GenePharma Company (Shanghai, China). For cell transfection, HepG2 and SMMC-7721 cells were cultured to reach 80% confluence, and transfected with 40 nM of siPHF6 or the corresponding negative control (NC) using the Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The cells without any transfection were used as blank control. After 48 h transfection, the knockdown efficiency was determined, and the following loss-of-function assays were performed.
RNA isolation and quantitative real-time PCR
Total RNA was extracted from tissues or cell lines using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. The cDNA was synthesized by PrimeScript reverse transcriptase reagent kit (Takara, Osaka, Japan). The quantitative real-time PCR (qRT-PCR) was performed on an ABI 7900HT system (Applied Biosystems, Carlsbad, CA) with the SYBR-Green Master Mix (Takara Biotechnology Co., Ltd., Dalian, China) with GAPDH as an internal control. The following primers were used for qRT-PCR detection: PHF6-forward: 5′-CAGCCACCCGAGATTGAGCA-3′, PHF6-reverse: 5′-TAGTAGCGACGGGCGGTGTG-3′;SNAI1-forward: 5′-TCGGAAGCCTAACTACAGCGA-3′, SNAI1-reverse: 5′- AGATGAGCATTGGCAGCGAG -3′, SNAI2-forward: 5′- CGAACTGGACACACATACAGTG-3′, SNAI2-reverse: 5′- CTGAGGATCTCTGGTTGTGGT -3′, β-catenin-forward: 5′- CCTATGCAGGGGTGGTCAAC-3′, β-catenin-reverse: 5′- CGACCTGGAAAACGCCATCA -3′, C-MYC-forward: 5′- GGCTCCTGGCAAAAGGTCA-3′, C-MYC-reverse: 5′- CTGCGTAGTTGTGCTGATGT -3′, CCND1-forward: 5′- GCTGCGAAGTGGAAACCATC-3′, CCND1-reverse: 5′- CCTCCTTCTGCACACATTTGAA -3′GAPDH-forward: 5′-TGGTATCGTGGAAGGACTC-3′, GAPDH-reverse: 5′-AGTAGAGGCAGGGATGATG-3′. Relative expression level of PHF6 was calculated according to the 2−ΔΔCt method.
EdU proliferation assay
Cell Light™ EdU Apollo®488 In Vitro Imaging Kit (Ribobio, Guangzhou, China) was used to determine cell proliferation according to the manufacturer's instruction. In brief, cells were seeded onto six-well plates at a density of 2 × 10 6 cells per well and cultured overnight. Then cells were incubated with 50 μM of EdU labeling medium for 2 h. Then cells were washed with PBS and fixed with 4% paraformaldehyde for 20 min. After washing once with PBS, the cells were stained with 300 μL 1 × Apollo solution for 10 min and washed three times in 0.5% TritonX-100, followed by detecting the percentage of EDU-positive cells by BD AccuriTM C6 flow cytometry (Beckman Coulter, Brea).
Colony formation assay
After 48 h transfection, cells were plated onto a six-well plate at a density of 500 cells per well and cultured for consecutive 2 weeks until colony formation. Then the naturally formed colonies were washed with PHBS and fixed with methanol for 10 min. After staining with 0.1% crystal violet (Sigma-Aldrich Co.) for 30 min, the colonies (>50 cells per colony) were observed and counted under a light microscope.
Transwell migration assay
A migration assay was performed using Transwell chambers (8 μm pore size, Corning Incorporated, NY). Transfected cells (3 × 10 4 ) were resuspended in 300 μL serum-free Dulbecco's Modified Eagle's medium (DMEM), and then seeded onto the upper chamber. The lower chamber was filled with 500 μL DMEM supplemented with 10% FBS as a chemoattractant. After 48 h incubation at 37°C, the cells that had migrated to the lower chamber were fixed with 100% methanol for 10 min at room temperature and stained with 0.5% crystal violet for 15 min. The stained cells were observed, and migrated cell number was counted under a light microscope (magnification, × 200).
Western blot analysis
Total protein samples were extracted from cells using radio immunoprecipitation assay buffer and quantified by the Bradford reagent (Bio-Rad Laboratories, Inc., Hercules, CA). Protein samples were separated through 12% SDS-PAGE and transferred onto polyvinylidene difluoride membranes (Sigma-Aldrich Co.). Then the membranes were blocked with 5% (w/v) skimmed milk in Tris-buffered saline with Tween 20 (TBST) for 1 h, and incubated with primary antibodies against PHF6, E-cadherin, Vimentin, and GAPDH (All from Cell Signaling Technology) overnight at 4°C. Subsequently, the membranes were washed with TBST and incubated with appropriate horseradish peroxidase-coupled secondary antibodies (Abcam, Shanghai, China) for 2°h. Finally, the separated protein signals were detected and visualized using enhanced chemiluminescence (Pierce Technology, Beijing, China).
Statistical analysis
All statistical computations were performed using SPSS software version 17 (IBM Corp., Armonk, NY). Data are presented as mean ± standard deviation (SD). The significance of difference in the expression levels of PHF6 between HCC tissue and paired adjacent tissue or between different HCC tissues was evaluated using the paired t-test. For the in vitro experiments, Student's t test or a one-way analysis of variance was used to assess the differences in the experimental groups. A p value of < 0.05 was considered statistically significant.
Results
PHF6 was upregulated in HCC tissues
Expression of PHF6 at the mRNA level was analyzed in 30 clinical samples of surgically removed HCC tissues and paired adjacent normal tissues using qRT-PCR. As shown in Figure 1A, PHF6 expression was significantly higher in HCC tissues compared with that in adjacent normal tissues (p < 0.01). Moreover, we found higher expression of PHF6 in III/IV stage of TNM classification, as compared with I/II stage of TNM classification (Fig. 1B, p < 0.001). According to the pathological diagnosis, in total 30 HCC tissues were further classified into two groups based on tissue differentiation (9 well, 11 moderate, and 10 poor) and three groups based on lymph node metastasis (16 No and 14 Yes). As shown in Figure 1C, a higher expression level of PHF6 was observed in poorly differentiated group than in the moderate/well differentiation group (p < 0.05). In addition, HCC tissues with lymph node metastasis exhibited significant elevated expression of PHF6 compared with those without lymph node metastasis (Fig. 1D, p < 0.001). Collectively, these data suggest that PHF6 may represent a candidate oncogene in HCC.
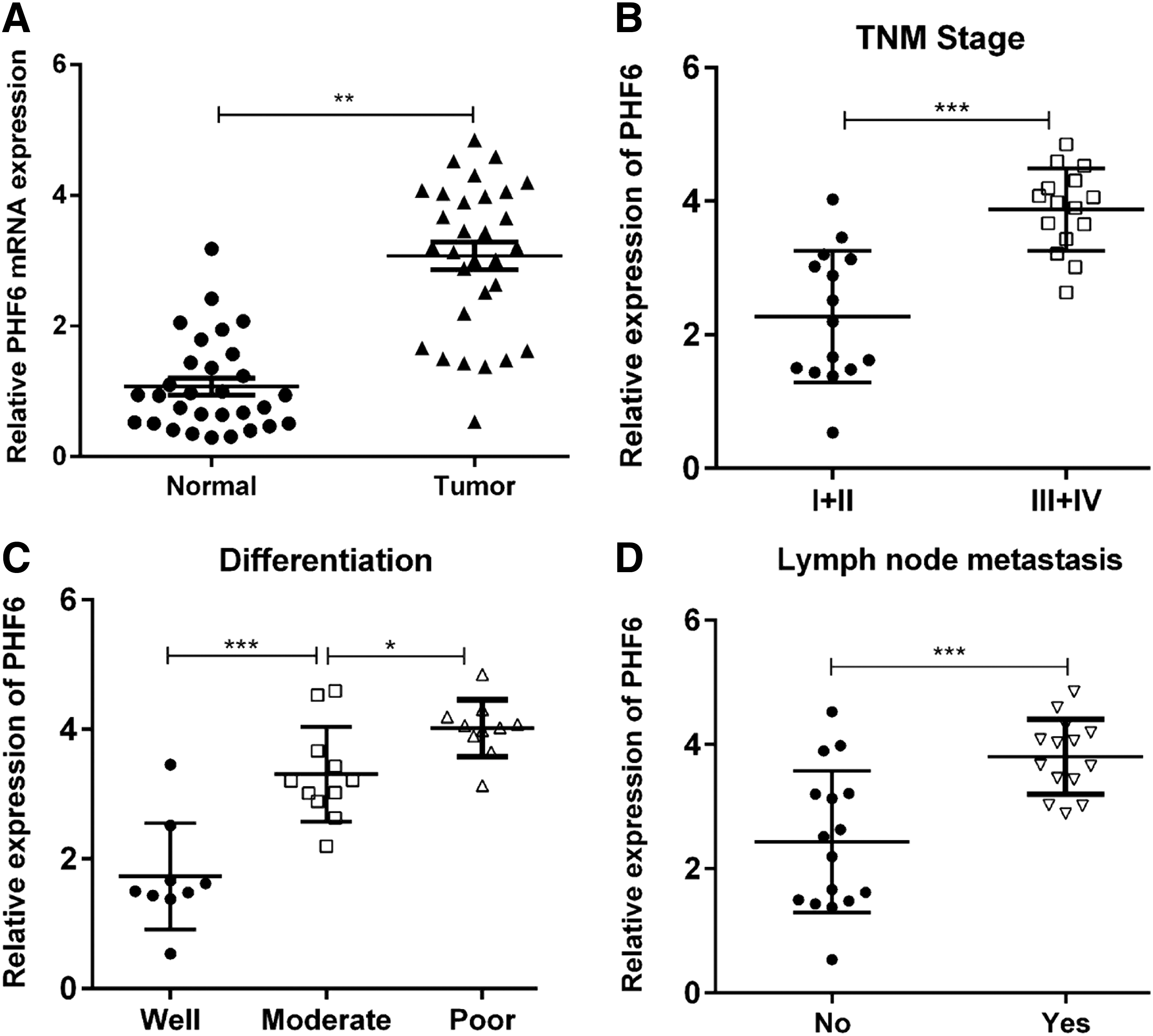
Expression of PHF6 in clinical HCC samples.
Silencing of PHF6 expression suppressed the proliferation and colony formation of human HCC cells
To investigate whether PHF6 plays a role in the biological behavior of HCC cells in vitro, several HCC cell lines were first used to determine the expression of PHF6. As shown in Figure 2A, the expression of PHF6 was obviously upregulated in all the four HCC cell lines, as compared with HL7702 cells. Notably, HepG2 and SMMC-7721 cell lines presented the highest PHF6 expression, thus were selected for the following loss-of-function assays. Using siRNA transfection, we found that the expression of PHF6 was significantly downregulated in HepG2, SMMC-7721, and Bel-7402 cells compared with NC or blank control (Fig. 2B, p < 0.001). Subsequently, the effect of PHF6 knockdown on cell proliferation in HCC cells was evaluated using EdU proliferation assay. As shown in Figure 2C, the percentage of EDU-positive cells was significantly decreased from NC group to siPHF6 group (p < 0.001). Furthermore, the colonies were remarkably reduced in HepG2, SMMC-7721, and Bel-7402 cells after siPHF6 transfection (Fig. 3, p < 0.01). These data suggested that silencing of PHF6 significantly suppressed HCC cell proliferation ability.

PHF6 knockdown suppressed cell proliferation of HCC cells.
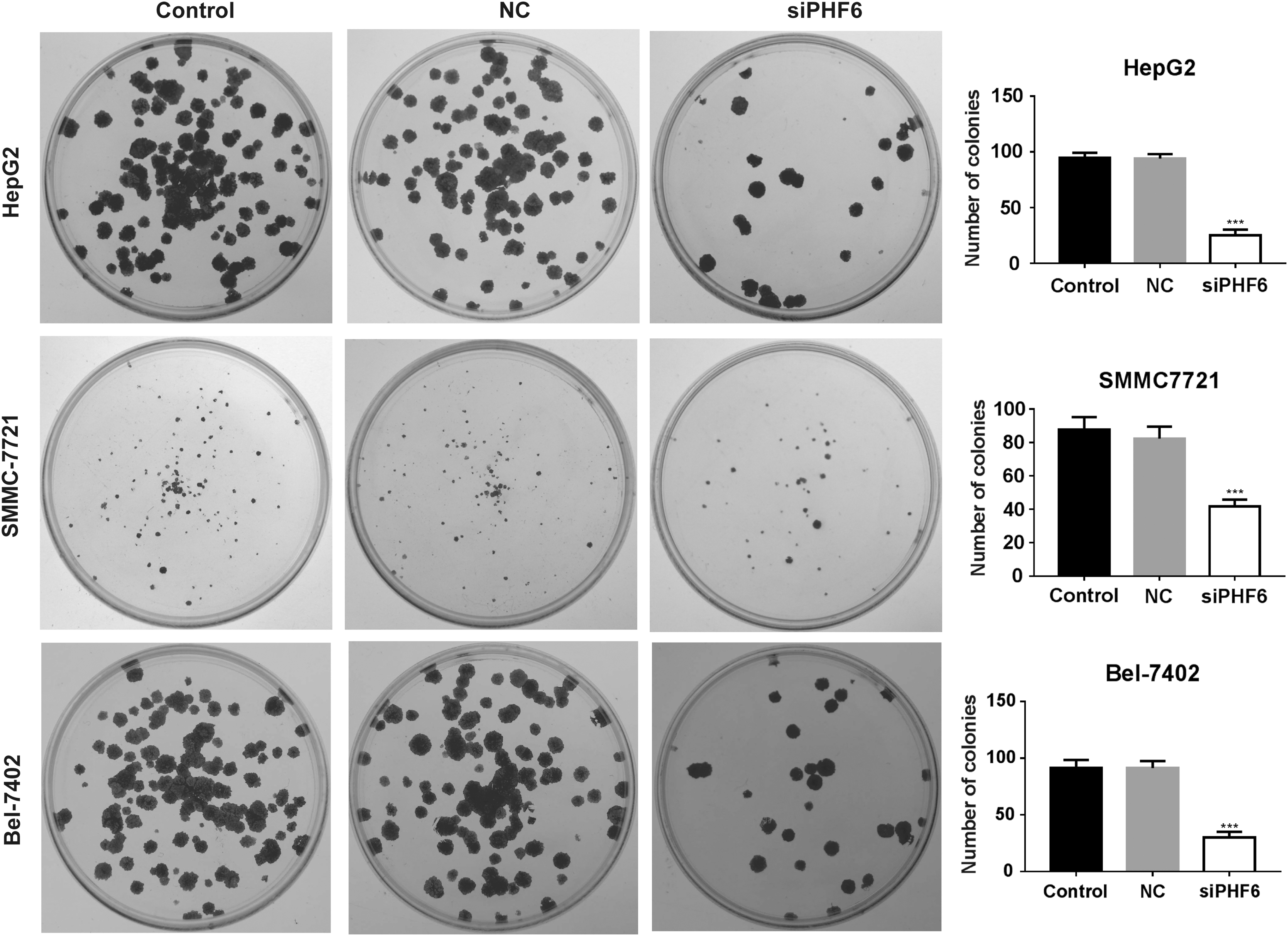
PHF6 knockdown impaired colony formation ability of HCC cells. Representative images of colony formation are shown in the left panel, and statistical quantification is presented in the right panel in HepG2, SMMC-7721, and Bel-7402 cells after PHF6 knockdown. ***p < 0.001.
Silencing of PHF6 expression inhibited the migration of human HCC cells
In addition, we performed a Transwell assay to examine the effect of silencing PHF6 on HCC cell migration. As shown in Figure 4A, PHF6 knockdown significantly reduced the number of migrated cells in HepG2, Bel-7402, and SMMC-7721 cells (p < 0.001). Furthermore, we analyzed the level of EMT-related genes, and found that the expression of SNAI1, SNAI2, β-catenin, C-MYC, and CCND1 was decreased by PHF6 knockdown in HepG2, Bel-7402, and SMMC-7721 cells (Fig. 4C). Then, we analyzed the protein level of E-cadherin and Vimentin, and found that the expression of the tumor metastasis-related protein E-cadherin was significantly increased, and Vimentin level was decreased in HepG2, SMMC-7721, and Bel-7402 cells with PHF6 knockdown, compared with NC or control cells (Fig. 4B). These data suggested that silencing of PHF6 significantly inhibited HCC cell migration ability.
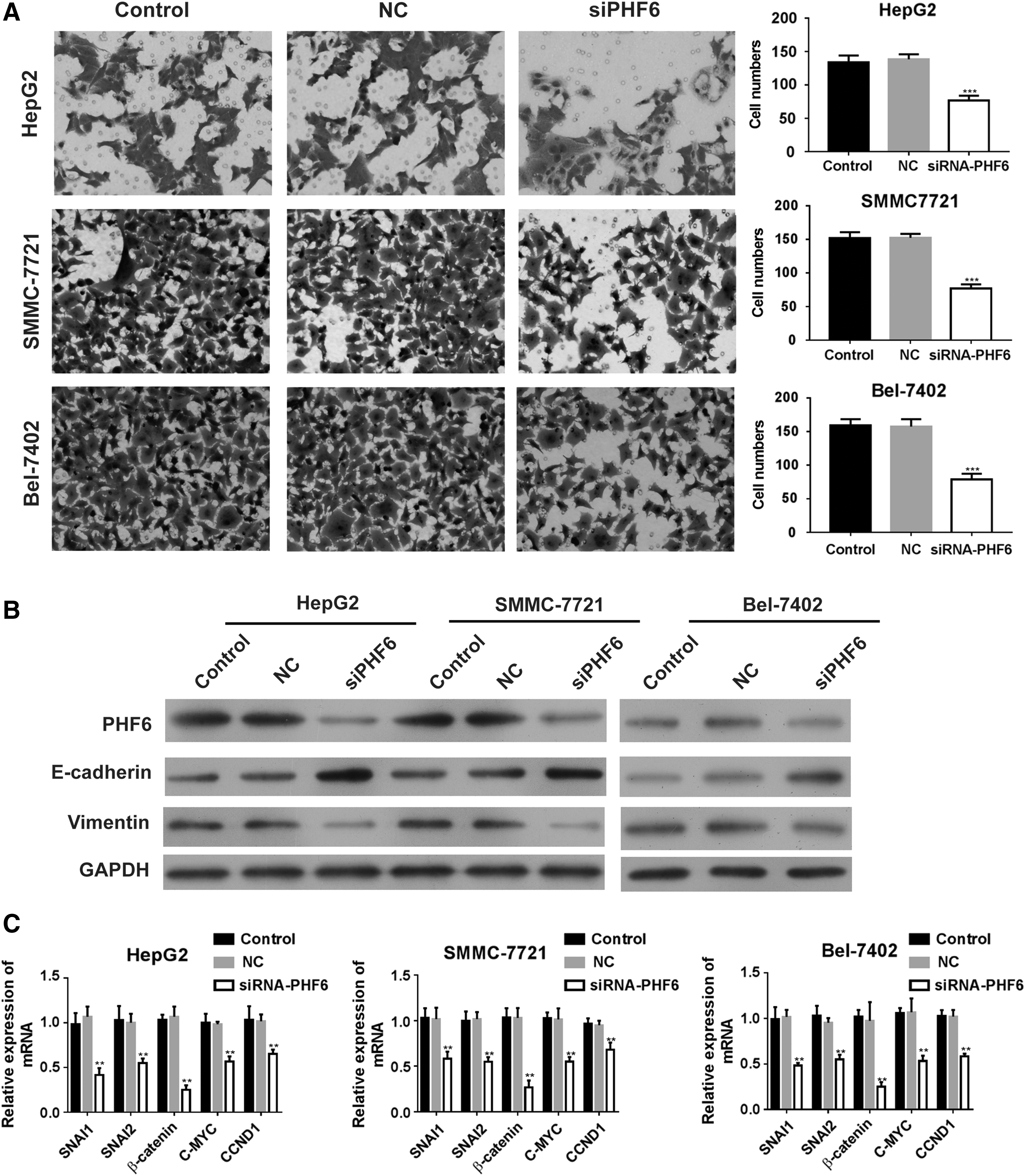
PHF6 knockdown inhibited cell migration ability of HCC cells.
Discussion
Recently, PHF6 has been reported to function as a tumor suppressor or an oncogene in a lineage-dependent manner, but its role in HCC has been poorly characterized. In this study, we found higher expression of PHF6 in HCC tissues, and it was positively correlated with TNM stage, differentiation, and lymph node metastasis, supporting the possibility that PHF6 contributes to the pathogenesis of HCC. Consistent with this notion, PHF6 could drive malignant transformation of developing thymocytes. 15 In addition, PHF6 was remarkably overexpressed in breast cancer and colorectal cancer. 16
To further confirm the role of PHF6 in HCC, we selected two HCC cell lines with higher PHF6 expression to analyze its function by performing loss-of-function assays. Our preliminary results showed that PHF6 knockdown significantly inhibited cell proliferation and colon formation ability in HCC cells. As a nucleolus, ribosomal RNA promoter-associated protein, PHF6 deficiency impairs cell proliferation and causes the accumulation of DNA damage in cells, indicating the key function of PHF6 in regulating rRNA synthesis. 17 Moreover, hairpin-mediated knockdown of PHF6 leads to impaired growth of B-ALL cells in vivo. 18 Thus, we could infer that PHF6 might play a positive role in the cell growth and proliferation of HCC cells. In addition to cell proliferation, we demonstrated that silencing of PHF6 remarkably suppressed cell migration in HCC cells, accompanying with upregulation of E-cadherin and downregulation of Vimentin. We also found that the mRNA level of SNAI1, SNAI2, β-catenin, C-MYC, and CCND1 was decreased by PHF6 knockdown in HepG2, Bel-7402, and SMMC-7721 cells. Epithelial–mesenchymal transition (EMT) is an important process characterized by the loss of cell polarity and cell–cell contact, and is activated in tumor migration and metastasis. 19 As the epithelial marker, E-cadherin is downregulated, while mesenchymal marker Vimentin is upregulated in the activation of EMT process. 20 Therefore, inhibition of EMT may interfere with tumor progression. The results of this study suggested that PHF6 knockdown may inhibit the EMT process by upregulating E-cadherin and upregulating Vimentin. However, the proposed targets in studies were still unclear, and we explore the targets of PHF6 further in the follow-up study.
Taken together, our findings revealed that silencing of PHF6 may significantly inhibit proliferation and migration and EMT in HCC cells, and targeting PHF6 might be a promising strategy for HCC therapy. In the future, further studies are needed to elucidate and understand the potential molecular function and clinical applications of PHF6.
Footnotes
Acknowledgments
This study was supported by The Longyan Science and Technology Plan Project, Number: 2017LY67; The Xiamen Science and Technology Plan Project, Grant/Award Number: 3502Z20164020; Natural Science Foundation of Fujian Province, Grant/Award Number: 2015J01541; The Project Funding for the Training of Young Talents in the Health System of Fujian Province, Grant/Award Number: 2014-ZQNJC-42.
Disclosure Statement
No competing financial interests exist.