
Abstract
Background:
Earlier we assumed that small molecules selectively accumulated in cancer cells might have a role in a defense system capable of killing cancer cells. We reported earlier that an experimentally selected mixture of substances present in the serum (“active mixture,” AM) shows a selective toxic effect in vitro and in vivo on various cancer cells. In this study we investigated additional compounds found in the serum to further expand our knowledge of this defense system.
Materials and Methods:
The cell proliferation was detected by WST-1 assay. The mRNA level of the examined genes was measured by quantitative real-time polymerase chain reaction.
Results:
We identified 34 additional compounds (
Conclusions:
The newly identified compounds significantly enhance the anticancer effect of AM. The components of AM and NM together may form part of a defense system capable of killing cancer cells and are worthy of further investigation.
Introduction
The antitumor defense system of higher order species uses mechanisms sensing and repairing DNA damage, processes regulating the cell cycle, molecules playing a role in the regulation of apoptosis, and the antitumor effect of the immune system. 1 During our earlier work on the basis of epidemiological and experimental data we assumed that beside the immunological and nonimmunological surveillance an additional defense mechanism might provide protection against tumor development.
Through our research we have focused on small molecular weight compounds (amino acids, monosaccharides, nucleobases, etc.) that are present in the circulatory system and are differentially taken up by tumor and normal cells. 2 It has been shown that in addition to glucose many molecules (amino acids, vitamins) are accumulated in cancer cells. 3 –6 The elevated uptake of these substances by cancer cells is utilized in positron emission tomography, 7 and based on the accumulation of amino acids or vitamins potential targeting strategies have been described. 8,9 According to our hypothesis, some of the accumulated substances beside their role in metabolism might participate in a defense system capable of killing cancer cells.
We have experimentally identified small molecular weight compounds present in the serum whose mixture (“active mixture,” [AM]) showed a selective toxic effect in vitro and in vivo on different cancer cell lines.
10,11
We have analyzed mainly different
The aim of this study was to find additional compounds present in the serum that can enhance the effect of AM. We analyzed another 130 molecules that were not included in the previous studies and from these we identified 34 additional substances (
Furthermore, we demonstrate that the mixture of the newly identified substances (“new mixture,” [NM]) applied in combination with AM has a significantly greater cell growth inhibitory effect on different cancer cells, and more efficiently activates genes involved in apoptosis than either NM or AM applied alone. In addition, we provide evidence that various combinations of the compounds of NM and AM always produce an enhanced antitumor activity compared with the effect of AM.
Materials and Methods
Cell culture
HeLa (human cervix adenocarcinoma), 4T1 (mouse mammary carcinoma) cell lines were obtained from the American Type Culture Collection through LGC Standards GmbH, Germany. MCF-7 (human breast adenocarcinoma), PC-3 (human prostate adenocarcinoma), Caco-2 (human colorectal adenocarcinoma, male), and HepG-2 (human hepatocellular carcinoma, male) cell lines were obtained from The European Collection of Authenticated Cell Cultures through Sigma-Aldrich. Cells were expanded and early passage stocks were stored under liquid nitrogen. All stocks were tested for mycoplasma with the Mycoplasma Detection Kit both before cryopreservation and after thawing. Cancer cells were cultured in minimal essential medium (Sigma-Aldrich, Budapest, Hungary) supplemented with 10% (v/v) fetal calf serum, 100 U/mL penicillin, and 0.1 mg/mL streptomycin. Human renal epithelial (HRE cells pooled from donors with different sex) cells were obtained from Lonza and cultured in renal epithelial cell basal medium (Lonza, Szeged, Hungary) supplemented with human epidermal growth factor, hydrocortisone, epinephrine, insulin, triiodothyronine, transferrin, GA-1000, and 0.5% fetal bovine serum. Cells were incubated at 37°C in a humidified atmosphere at 5% CO2.
Active mixture
The selection of the components of AM has been described previously.
10,11
The composition of 100% AM is as follows: 4 mM
New mixture
The composition of the 100% NM, identified in this article, is as follows: 2 mM
WST-1 assay
The cells were seeded at a density of 4 × 10 3 cells/well on 96-well plates, and then the cells were treated with the test compounds for 48 h in triplicates. 5-fluorouracil (5-FU) (Sigma-Aldrich) was used as a positive control of the inhibition of cell proliferation in the concentration range of 5–60 μM. The number of viable cells was measured with WST-1 cell proliferation reagent 15 according to the manufacturer's instructions (Roche, Budapest, Hungary). At the end of the treatment, cells were washed with phosphate-buffered saline, and then 90 μL phenol-red free RPMI medium (Sigma-Aldrich) and 10 μL WST-1 reagent were added to the cells. The cells were incubated for 30 min at 37°C, and then the optical density of each well was measured at 450 nm using an EL800 microplate reader. The results were expressed as the percentage of the untreated control. Experiments were repeated three times. The half maximal inhibitory concentration (IC50) was determined by fitting the dose–response curve using the OriginPro 8.6 software.
Quantitative real-time polymerase chain reaction
Total RNA was isolated with PureLink RNA Mini Kit (Thermo Fisher Scientific, Budapest, Hungary) and was treated with DNase I (Sigma-Aldrich). cDNA was prepared with High-Capacity RNA-to-cDNA Kit (Thermo Fisher Scientific). Polymerase chain reaction (PCR) primers used for real-time quantitative amplification of the human housekeeping genes B2M, GAPDH, HPRT1, RPL32, and PPIA were described previously. 16,17 PCR primers for human APAF1, BAD, BAK1, BAX, BCL2, BCL2L1, BCL2L11, BIRC2, BIRC3, CASP3, CDKN1A, CDKN2A, IKBKG, NFKBIA, NFKB1, NFKB2, REL, RELA, and RELB were also described previously. 14 All other PCR primers were designed by Primer Express Software, primer sequences are listed in Supplementary Table S1. The expression levels of BBC3, PMAIP1, RPL32 were measured with TaqMan gene expression assays (BBC3:Hs00248075_mL, PMAIP1: Hs00560402_mL, RPL32: Hs00851655_gL from Thermo Fisher Scientific). RPL32 was used for normalization. PCR reactions were run in triplicates using PowerUp SYBR Green Master Mix (Thermo Fisher Scientific) or TaqMan gene expression master mix II, no uracil n-glycosylase (Thermo Fisher Scientific) on an ABI StepOne Real-Time PCR System (Applied Biosystems, Budapest, Hungary). The relative quantity values were calculated by dividing the normalized target gene expression measured in the treated samples by that of the untreated control samples. 18
Statistical analysis
Statistical analysis was performed with IBM SPSS Statistics 22. Normal distribution of the data was examined by Shapiro–Wilk test, homogeneity of variances was assessed with Levene test, and then one-way analysis of variance followed by Bonferroni post hoc test was performed. Values of p < 0.05 were considered statistically significant.
Data availability
The datasets generated during this study are available in the Open Science Framework repository,
Results
Identification of molecules capable to enhance the cell growth inhibitory effect of AM
To find additional compounds capable to increase the effect of AM we investigated an additional 130 compounds present in the serum. To be able to detect the potentiating effect of various compounds found in the serum, AM was used in a concentration that killed ∼50%–60% of cancer cells. HeLa cells were treated with 50% AM alone or in combination with the newly examined 130 individual compounds. The newly examined compounds were applied in per se nontoxic concentrations determined in preliminary experiments. The cells were treated for 48 h, and then the cell proliferation was measured by WST-1 assay. After 48 h of treatment, we selected those compounds for further evaluation that were able to significantly enhance the effect of AM. Of the 130 examined compounds 34 was able to significantly potentiate (p < 0.001) the cancer cell killing effect of AM (Fig. 1). Treatment with 50% AM reduced the number of living cells to 46.25% ± 1.36% of the untreated control. Each of the newly identified 34 compounds significantly enhanced this effect, although the extent of the enhancement was different among the compounds: the number of living cells was reduced to 22.41%–40.36% of the untreated control. The 34 newly identified compounds are the following:
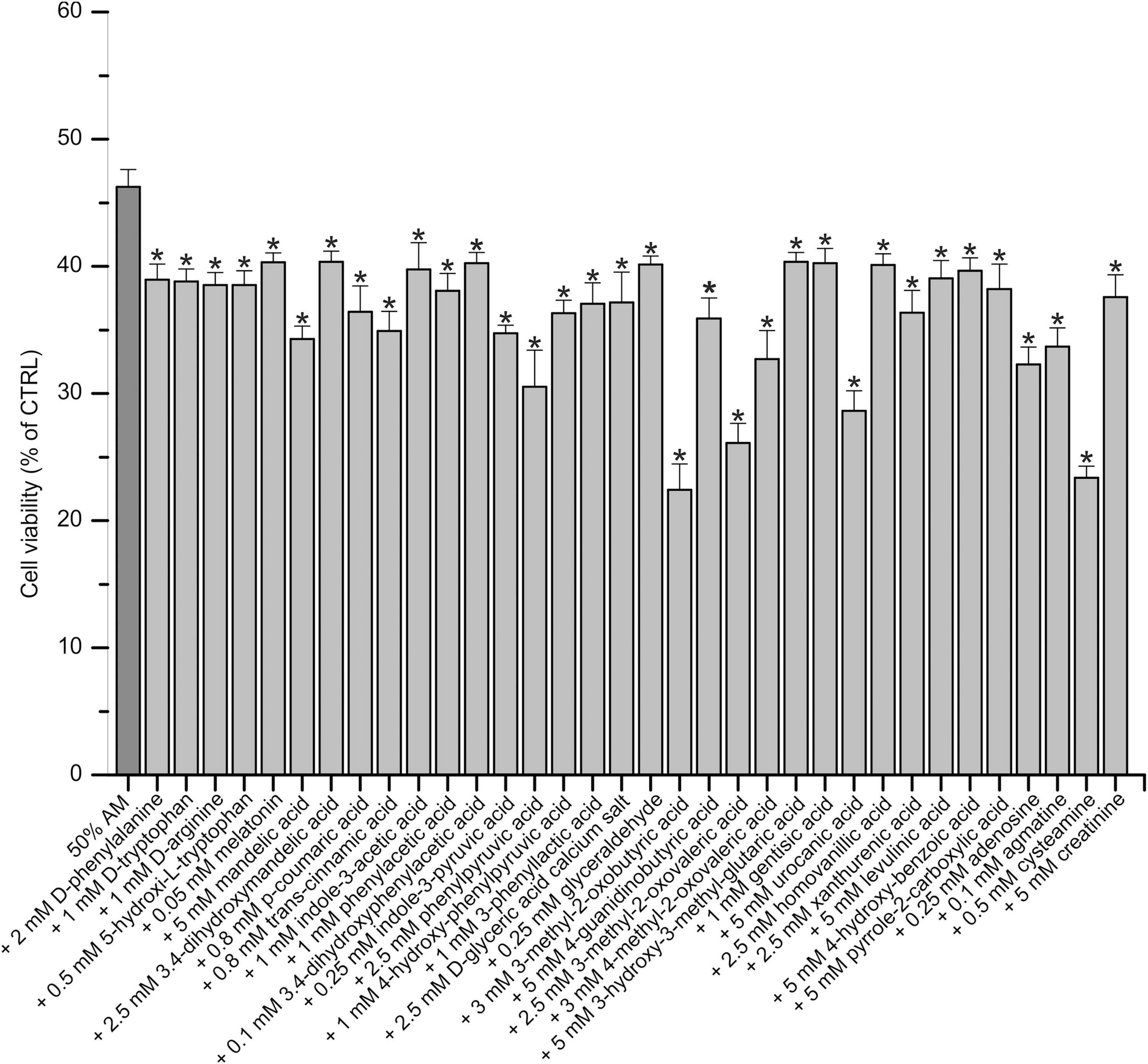
The newly identified 34 compounds enhance the cell growth inhibitory effect of AM. HeLa cells were treated with 50% AM alone or with the individual combination of 34 compounds and 50% AM for 48 h. The newly examined compounds were applied in per se nontoxic concentrations. Results are expressed as percentage of untreated control cells. The bars represent the average ± standard deviation of three independent experiments. *p < 0.001 versus 50% AM (ANOVA, Bonferroni test). AM, active mixture; ANOVA, analysis of variance.
Comparison of the effect of NM and AM on cancer cells
Next we investigated the effect of the mixture comprising the newly identified 34 compounds, called NM, compared with AM on HeLa (Fig. 2A), PC-3 (Fig. 2B), MCF-7 (Fig. 2C), Caco-2 (Fig. 2D), HepG-2 (Fig. 2E), 4T1 (Fig. 2F) cancer cells and on HRE normal cells (Fig. 2G). For the optimal detection of differences in efficacy, we used dilution series of the different mixtures (10%, 15%, 20%, 25%, and 30%). Figure 2 shows that NM has a significantly greater cell growth inhibitory effect compared with the treatment with AM (p < 0.001 vs. corresponding dilution of AM) on the different cancer cells. The combination of AM and NM was also tested. We found that the combined treatment (AM+NM) was able to more efficiently inhibit the cancer cell growth compared with AM or NM (p < 0.001 vs. corresponding dilution of NM and vs. corresponding dilution of AM), and the 30% AM+NM killed all the cells after 48 h of incubation. These results demonstrate that AM, NM, and the combined mixture reduced the number of cancer cells in a dose-dependent manner, but showed differences in their potency. However, the proliferation of HRE cells (Fig. 2G) was not decreased by any of the mixtures, rather they slightly increased the cell number. As a positive control of the inhibition of cell proliferation we used 5-FU in the concentration range of 5–60 μM. Figure 2H shows that 60 μM of 5-FU produced approximately the same degree of inhibition as 15% AM+NM (5-FU: 38.44%–48.2% of the untreated control vs. 15% AM+NM: 34.49%–44.37% of the untreated control) on the tested human cancer cell lines (HeLa, PC-3, MCF-7, Caco-2, and HepG-2), whereas the murine 4T1 cell line was more sensitive to 5-FU (20 μM killed all the cells), and 20% of AM+NM was required to produce approximately the same inhibition as we have seen on the human cell lines (44.44% of the untreated control).
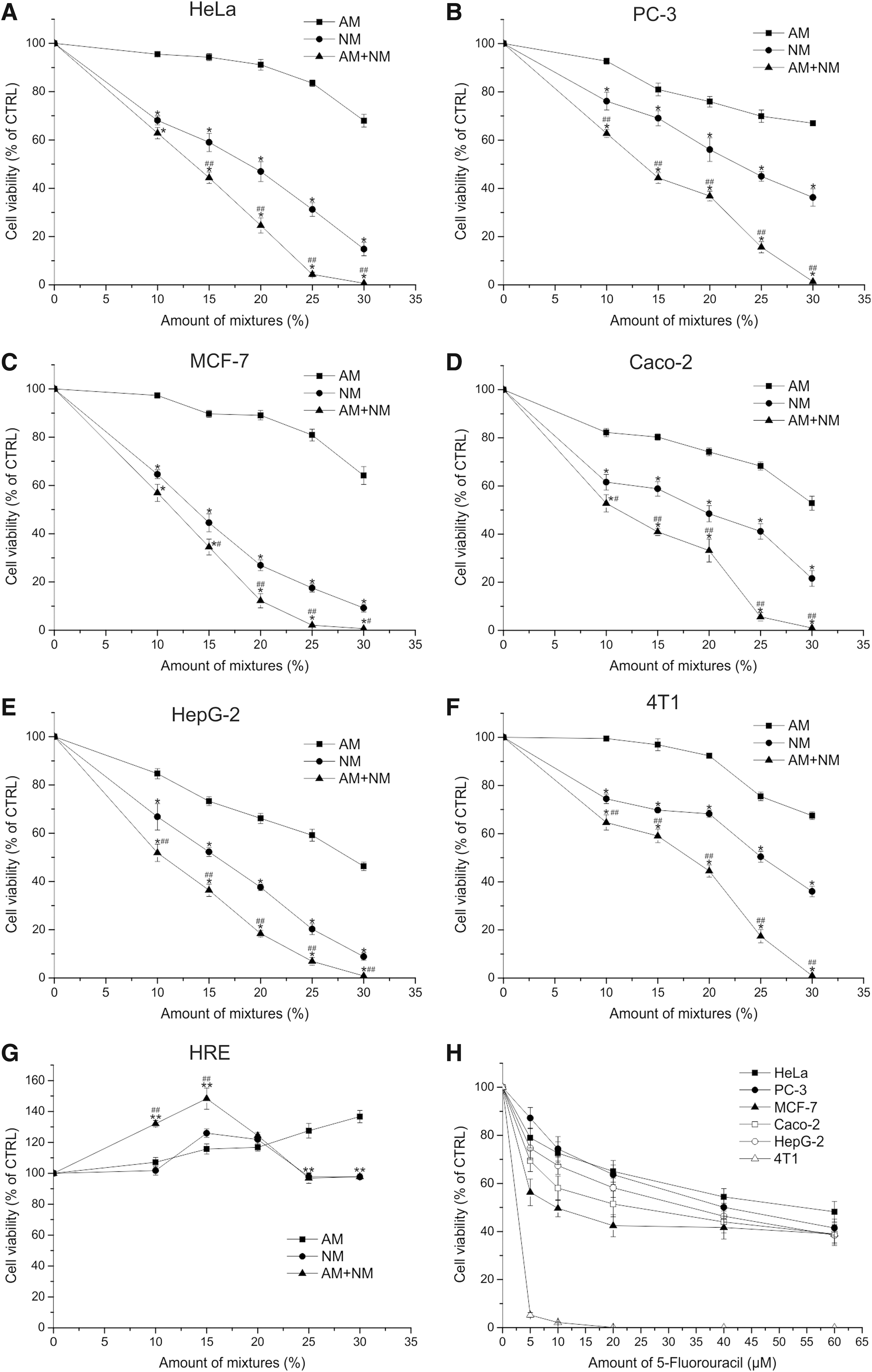
Comparison of the effect of NM and AM on the growth of cancer and normal cells. HeLa
AM and NM influence the expression of genes involved in apoptosis
Next we measured the expression of certain pro- and antiapoptotic genes, as well as genes regulating the cell cycle by quantitative PCR after the treatment of HeLa cells with 30% AM, 20% NM, or the combination of 30% AM and 20% NM (Fig. 3). Treatment with 30% AM had a slight or no effect on the expression of genes investigated compared with the control. Combined treatment with 30% AM and 20% NM significantly increased the expression of the proapoptotic BH3-only protein encoding BIM (3.23-fold, p < 0.001), PUMA (4.40-fold, p < 0.001), NOXA (8.17-fold, p < 0.001), and the antiapoptotic member of BCL-2 family encoding BCL-2 (1.59-fold, p < 0.05) compared with the untreated control. The expression level of the proapoptotic death receptor genes, DR4 (TNFRSF10A) (1.97-fold, p < 0.001) and DR5 (TNFRSF10B) (6.82-fold, p < 0.001) were increased, together with the antiapoptotic protein encoding cIAP (1.85-fold, p < 0.05) and cIAP2 (1.30-fold, p < 0.05) transcript levels. Among the nuclear factor κB (NF-kB) signaling-related genes the expression level of NFKB1 (1.90-fold, p < 0.001), NFKB2 (2.33-fold, p < 0.001), IKBKG (1.80-fold, p < 0.05), RELA (1.94-fold, p < 0.05), and RELB (3.72-fold, p < 0.001) were increased. The level of CDKN1A was increased to 4.82-fold (p < 0.001). Twenty percent NM mixture applied alone also induced changes in the expression of pro- and antiapoptotic genes but to a lesser extent than AM+NM, levels of NOXA (2.47-fold, p < 0.05), cIAP2 (2.29-fold, p < 0.001), TNFRSF10A (1.49-fold, p < 0.05), NFKB2 (1.65-fold, p < 0.05), and RELB (3.24-fold, p < 0.001) were increased.
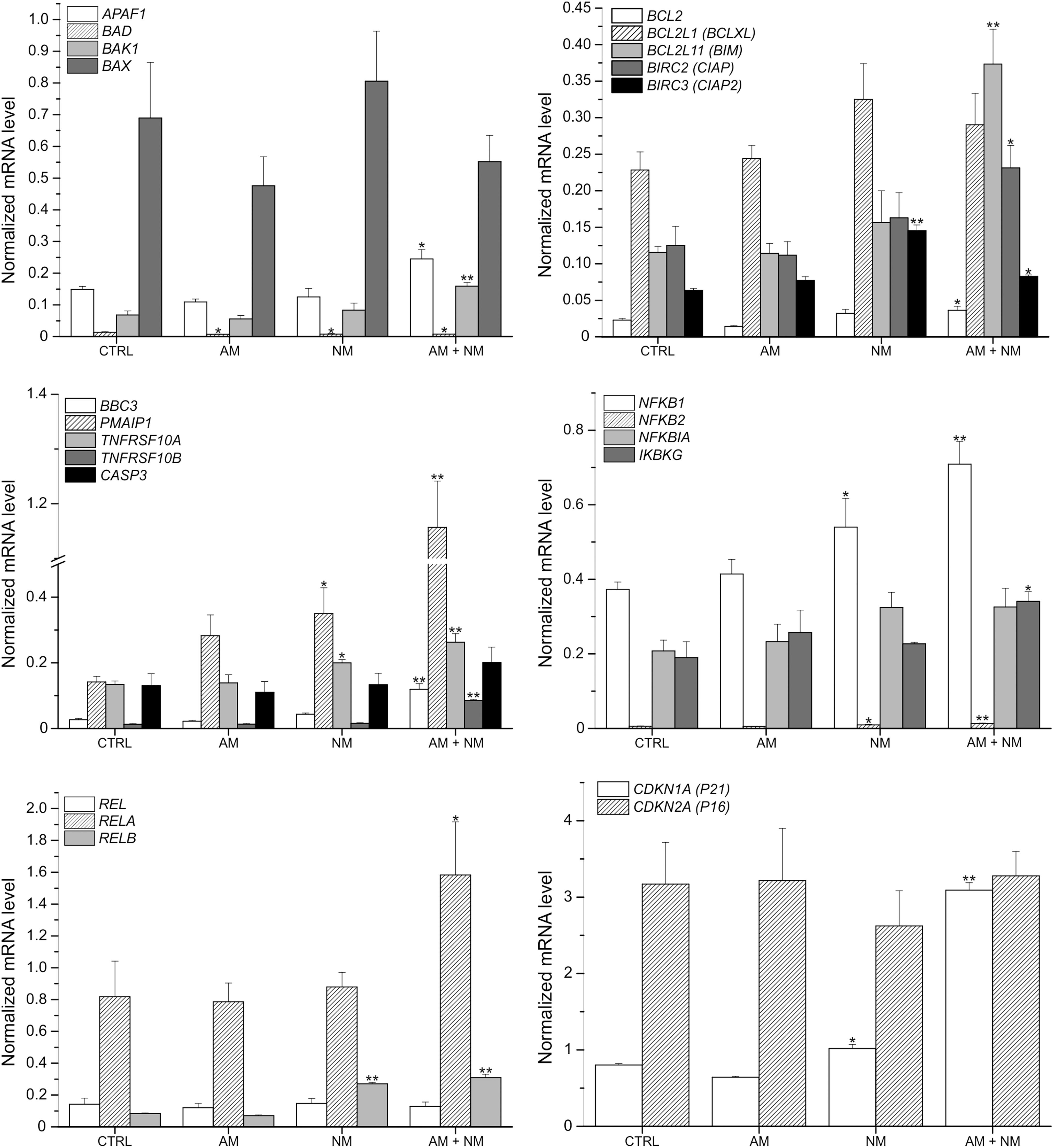
NM and the combination of AM and NM influence the expression of genes involved in apoptosis and cell cycle regulation. HeLa cells were treated with 20% AM, 30% NM, or the combination of 20% AM and 30% NM for 24 h. mRNA levels were detected with quantitative real-time polymerase chain reaction. The bars represent the average ± standard deviation of three independent experiments. **p < 0.001; *p < 0.05 versus control (ANOVA, Bonferroni test). CTRL, untreated control.
Comparison of the tumor cell killing effect of various mixtures of the newly identified substances with mixtures of compounds identified earlier
To further investigate the potentiating effect of the newly identified 34 compounds on the compounds identified earlier, we compared 15 component mixtures comprising 5 compounds selected from NM (NM5.1, NM5.2, NM5.3) and 10 compounds selected from AM (AM5.1+AM5.2, AM5.2+AM5.3, AM5.1+AM5.3) with a 15 component mixture comprising compounds solely from AM (AM5.1+AM5.2+AM5.3) (Fig. 4).

Fifteen component mixtures that comprise 10 earlier identified compounds and 5 newly identified compounds have higher cell growth inhibitory effect on HeLa cells than mixtures containing exclusively components of AM. The cells were treated with the dilution series of the indicated combinations of AM5.1, AM5.2, AM5.3, NM5.1, NM5.2, and NM5.3 for 48 h.
For the optimal detection of differences in efficacy, we used dilution series of the different mixtures (20%, 40%, 60%, 80%, 100%). Treatment with 60% AM5.1+AM5.2+AM5.3 reduced the number of living cells to 31.93% ± 1.59%–37.91% ± 2.43% of the untreated control. When AM5.1 or AM5.2 or AM5.3 was replaced by a 60% mixture containing 5 compounds selected from the newly identified 34 compounds, the tumor cell killing effect was significantly enhanced, the number of living cells was reduced to 8.06% ± 1.26% (p < 0.001)–27.45% ± 2.09% (p < 0.001) of the untreated control. IC50 values of the 15 component mixtures comprising compounds solely from AM were between 49.27% and 53.55%. IC50 values of the mixtures containing 5 compounds from NM and 10 compounds from AM were between 40.93% and 46.40%.
Next, we evaluated the effectiveness of 15 component mixtures that contain 10 compounds from NM (NM5.1+NM5.2, NM5.2+NM5.3, NM5.1+NM5.3) and 5 compounds from AM (AM5.1, AM5.2, AM5.3) compared with the mixture of AM5.1+AM5.2+AM5.3 (Fig. 5). Treatment with 60% AM5.1+AM5.2+AM5.3 reduced the number of living cells to 33.49% ± 2.14%–36.39% ± 1.98% of the untreated control. When either two of AM5.1, AM5.2, or AM5.3 mixtures was replaced by two 60% mixtures comprising five compounds selected from NM (NM5.1+NM5.2, NM5.2+NM5.3, NM5.1+NM5.3), the cell growth inhibitory effect was even more pronounced. The number of living cells was reduced to 3.82% ± 0.85%–25.45% ± 2.72% to the untreated control. IC50 values of the 15 component mixtures comprising compounds solely from AM were between 50.21% and 52.29%. IC50 values of the mixtures containing 10 compounds from NM and 5 compounds from AM were between 27.69% and 42.70%.
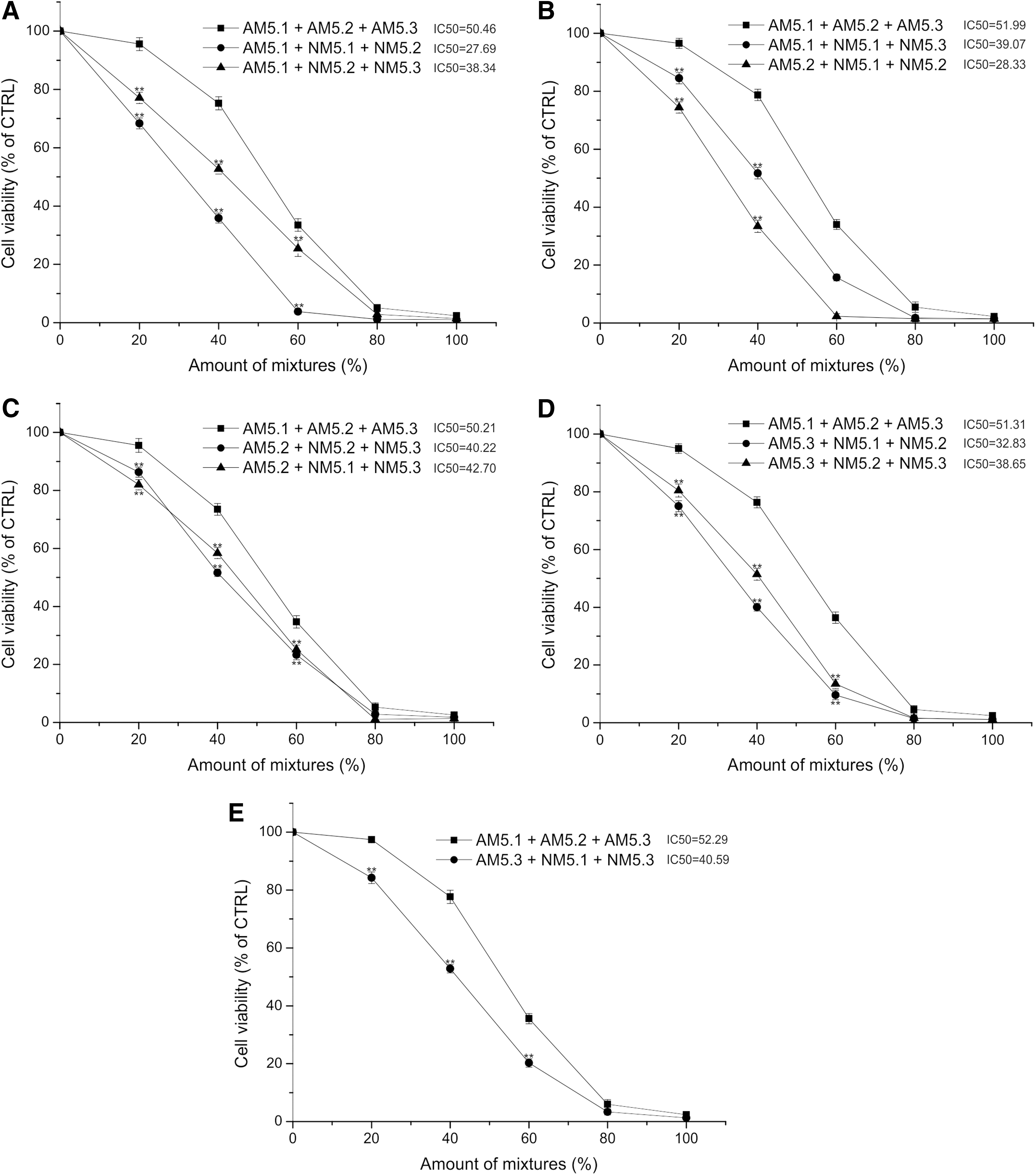
Fifteen component mixtures that comprise 5 earlier identified compounds and 10 newly identified compounds have higher cell growth inhibitory effect on HeLa cells than mixtures containing exclusively components of AM. The cells were treated with the dilution series of the indicated combinations of AM5.1, AM5.2, AM5.3, NM5.1, NM5.2, and NM5.3 for 48 h.
These results show that combinations of the newly identified compounds with the earlier identified compounds always produced an enhanced cancer cell killing effect compared with the single application of mixtures comprised solely of compounds identified earlier (AM).
Discussion
Previously we demonstrated that a mixture of experimentally selected substances (amino acids, vitamins, nucleobases, etc.) found in the serum called AM induced apoptosis of cancer cells both in vitro and in vivo and had a significant tumor inhibitory effect in vivo. 12,14 On the basis of our assumption that some of the accumulated substances might be part of an antitumor defense system, 13 in this study we tested another 130 compounds found in the serum and we identified 34 additional substances that are capable to kill cancer cells and synergistically enhance each other's and the earlier identified compound's effect. We applied the tested compounds in a per se nontoxic concentration; thus, it can be stated that the 34 newly identified compounds potentiated in a synergistic manner the tumor cell killing effect of AM on HeLa cells.
To further investigate the synergistic effect of the newly identified substances, we performed additional cell viability assays with randomly selected compounds of NM and/or AM. We have tested other combinations as well, which always produced the same results (data not given). These results raise the possibility that any given combination of the newly identified compounds with the earlier identified compounds would produce an enhanced tumor cell killing effect compared with the single application of any given combination of the earlier identified compounds.
The components of NM are mainly
Besides, there are reports that demonstrate that agmatine, metabolite of
To further evaluate the anticancer activity of the newly identified substances, the effect of the mixture of the new substances (NMs) was investigated on various cancer cells. We applied the different substances in a per se nontoxic concentration and showed that NM and the combined mixture of AM and NM have a significant growth-inhibiting effect on HeLa human cervix adenocarcinoma, MCF-7 human breast adenocarcinoma, PC-3 human prostate adenocarcinoma, Caco-2 human colorectal adenocarcinoma, HepG-2 human hepatocellular carcinoma, and 4T1 mouse mammary carcinoma compared with the mixture of AM. Treatment with any given mixtures caused no toxic effect on HRE normal cells. This is in agreement with our hypothesis that the mixture of substances we have selected has a specific inhibitory effect on cancer cells. Our findings that NM has anticancer activity on a broad spectrum of cancer cells with different histological origin in our view indicates that the inhibitory effect of NM could represent a general defense mechanism.
Furthermore, treatment with the combination of AM and NM elicited induction of PUMA, NOXA, BIM, and CDKN1A in HeLa cells, which points toward the induction of apoptosis through the mitochondrial pathway and probably leading to cell cycle arrest, is also in agreement with our previous result. 14 The expression level of two death receptors, DR4 and DR5, were also upregulated. As death receptors are important components of the extrinsic apoptotic pathway, 28 increased expression of DR4 and DR5 raises the possibility that this pathway also contributes to the cell growth inhibition caused by AM+NM. In this study, the treatment with AM was not able to activate the examined genes. This is most likely because of the fact that we applied a lower dose of AM compared with our previous reports to notice the synergistic effect of NM. However, treatment solely with NM could induce changes in gene expression but could not reach the same level as produced by the combined treatment.
Given the multicomponent nature of AM and NM, the anticancer activity of them is probably mediated by the interplay of several different mechanisms. In our earlier mechanistic studies of AM, which in part is composed of essential amino acids, we described the mitochondrial pathway of apoptosis induction and the upregulation of genes contributing to apoptosis induction and cell cycle arrest (PMAIP1, BBC3, CDKN1A). 14 In addition to our results it has been reported by Bonfili et al. that mixtures of essential amino acids inhibit proteasomal activity, induce autophagy, and apoptosis in a cancer-specific manner. 29 Of the components of NM, cinnamic acid has been reported to induce cytoskeletal damage and caspase activation in melanoma cells. 20 In addition, it has been shown to inhibit histone deacetylases and increase the expression of certain proapoptotic genes in colon cancer and pancreatic cancer cells. 21 p-coumaric acid is known to increase the production of reactive oxygen species and induce apoptosis through the mitochondrial pathway in colon cancer cells. 23 Agmatine has been shown to interfere with the polyamine metabolism by inhibition of ornithine decarboxylase in colon cancer cells. 25 There are no data about the possible mechanisms involved in the anticancer activity of every one of the components of AM or NM, and we would like to stress out that this study focused on the experimental selection of new substances (components of NM) and not on identifying all possible mechanisms involved. The various mechanisms that might contribute to the anticancer activity of AM+NM mixture are summarized in Figure 6; however, we would like to emphasize that there might be additional signaling pathways that could be activated because of the multicomponent nature of these mixtures.

Possible mechanisms contributing to the anticancer effect of AM+NM.
Cancer cells have altered metabolism to support their growth and survival. 30 The increased consumption of glucose accompanied by increased aerobic glycolysis is one example of that. Aerobic glycolysis and other cancer-associated changes in metabolism can alter the intracellular metabolite levels. 3,31 Many oncogenic mutations lead to the accumulation of certain metabolites with oncogenic potential. 31 For example, high level of fumarate and succinate, caused by the loss of fumarate hydratase and succinate dehydrogenase, can positively affect tumorigenesis by competitive inhibition of the of α-ketoglutarate-dependent dioxygenases that have a role in epigenetic regulation. 32 The elevated glycolytic flux can increase the level of glycolytic intermediates, such as methylglyoxal (MG) produced by the spontaneous decomposition of glyceraldehyde-3-phosphate and dihydroxyacetone phosphate. 33 MG may have oncogenic roles 31 ; however, there are emerging evidences that excess level of MG can inhibit various cancer cells. 34,35 Thus, changes in metabolite levels could have different consequences, some of them may inhibit cancer growth or others are required for tumorigenesis. 31
Thus, our overloading strategy of defined substances that may alter the metabolite level in a way that is deleterious for cancer cells might be a potential therapeutic approach.
Conclusions
We demonstrated that the newly identified compounds are able to significantly enhance the antitumor effect of AM. The combined mixture of AM and NM inhibits the growth of various cancer cells in vitro more effectively than the individual mixtures per se, without any toxic effect on normal cells.
The high number of compounds and the fact that they enhance each other's effect provide the opportunity to design mixtures that might be more effective against specific types of cancer.
Footnotes
Authors' Contributions
Conceptualization, G.K., D.S., and T.C.; Methodology, G.K., D.S., and T.C.; Formal analysis, G.K., D.S. and T.C.; Investigation, G.K., D.S., and T.C.; Writing–Original Draft, D.S., G.K., and T.C.; Supervision, G.K. and T.C.; Funding Acquisition, G.K. and T.C. The corresponding author declares that all coauthors have reviewed and approved the article.
Acknowledgment
This work has been supported in part by a Ministry for National Economy, Hungary grant GINOP-2.1.1.-15-2015-00046.
Disclosure Statement
G.K. owns a 15% share in Culevit Ltd.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.