
Abstract
Introduction:
It has been reported that long noncoding RNAs (lncRNAs) are crucial regulators in progression of human cancers, including colorectal cancer (CRC). However, the function of lncRNA ARAP1 antisense RNA 1 (ARAP1-AS1) in CRC remains unclear.
Aim:
The aim of this study was to investigate the function and molecular mechanism of lncRNA ARAP1-AS1 in CRC.
Results:
ARAP1-AS1 was highly expressed in CRC tissues and cell lines. ARAP1-AS1 knockdown suppressed cell migration, invasion, and epithelial–mesenchymal transition (EMT). YY1 transcription factor (YY1) enhanced the transcription activity of ARAP1-AS1. The YY1/ARAP1-AS1 axis promoted CRC cell migration and invasion. YY1/ARAP1-AS1 could regulate the Wnt/β-catenin signaling pathway.
Conclusions:
This study revealed that YY1-induced upregulation of ARAP1-AS1 promoted cell migration, invasion, and EMT process in CRC through the Wnt/β-catenin signaling pathway.
Introduction
Colorectal cancer (CRC) ranks third among the most common malignancies, which is the fourth leading cause of cancer-related death. 1,2 There are more than 1.2 million patients who were diagnosed with CRC each year. 3 Surgical resection, chemotherapy, and radiotherapy are the most common therapeutic methods for CRC. Despite therapeutic strategies having been improved, the overall survival rate of CRC patients within 5 years is still low. 4,5 Although molecular mechanisms underlying cancer progression have been studied and reported, the efficient molecular mechanism involved in CRC progression is still largely unknown. Therefore, it is necessary to investigate the novel molecular mechanism associated with the biological processes of CRC.
Long noncoding RNAs (lncRNAs) without the ability to code proteins have been shown to be crucial regulators in the occurrence and development of various tumors. 6 –11 It has been reported that lncRNAs regulate various biological processes at multiple levels. 12 Aberrant expression of lncRNAs has been identified to be an important factor for progression and development of malignancies. 13 –20 The aim of this study is to explore the mechanism of functional lncRNAs in CRC.
lncRNA ARAP1 antisense RNA 1 (ARAP1-AS1) has been reported to be an oncogene in bladder cancer. 21 However, its role in CRC is still unknown. Based on The Cancer Genome Atlas (TCGA) dataset, the authors found that ARAP1-AS1 was highly expressed in colorectal adenocarcinoma samples. Furthermore, the level of ARAP1-AS1 was examined in 82 pairs of CRC tissues and normal tissues. Since the expression of ARAP1-AS1 was correlated with metastasis, the authors performed functional assays to determine the effect of ARAP1-AS1 knockdown on CRC cell migration, invasion, and epithelial–mesenchymal transition (EMT) progress. Additionally, previous studies have shown that upregulation of lncRNAs is usually caused by their upstream transcription factors. 22 –26
In this study, both bioinformatic analysis and mechanism experiments revealed the interaction between transcription factor YY1 and ARAP1-AS1. Additionally, the Wnt/β-catenin signaling pathway was found to be a downstream pathway of ARAP1-AS1. Finally, rescue assays were conducted to demonstrate the function of the YY1/ARAP1-AS1 axis in regulating CRC cell migration and the Wnt/β-catenin signaling pathway. In conclusion, the present study revealed that the YY1/ARAP1-AS1 axis promoted CRC cell migration, invasion, and EMT progress through the Wnt/β-catenin signaling pathway.
Materials and Methods
Tissues samples
All CRC tissues and adjacent nontumor tissues (n = 82) were obtained from patients who received surgical resection at The First Affiliated Hospital of Wenzhou Medical University. After collection, samples were immediately frozen in liquid nitrogen and preserved at −80°C until total RNA isolation. No local or systemic treatments were performed in these patients before surgery. This study had the approval of the Ethics Committee of The First Affiliated Hospital of Wenzhou Medical University. All patients had signed written informed consent before the study.
Cell culture
The normal human colon epithelial cell line, HCoEpiC, and five human CRC cell lines (SW620, SW480, HT29, LoVo, and HCT116) were purchased from American Type Culture Collection (ATCC, Manassas, VA). These cell lines were then incubated in Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO), 100 U/mL penicillin, and 100 U/mL streptomycin (Invitrogen, Carlsbad, CA) in a humidified atmosphere with 5% CO2 at 37°C. The culture medium was replaced every 3 d and cell passage was performed when the cells grew up to 80%–90% confluence.
RNA extraction and quantitative real-time polymerase chain reaction
Total RNA was extracted from tissues or cells using TRIzol reagent (Invitrogen) based on the manufacturer's protocol. NanoDrop2000c (Thermo Scientific, Waltham, MA) was used to detect the quantity and quality of isolated RNAs. Total RNA was reverse transcribed to complementary DNA (cDNA) using the PrimeScript™ RT Master Mix (TaKaRa, Shiga, Japan). cDNA templates were amplified by real-time polymerase chain reaction (RT-PCR) using SYBR Green Master Mix (Applied Biosystems, Foster City, CA). PCR amplification was carried out as follows: 95°C for 5 min, followed by 40 cycles at 95°C for 5 s and 61°C for 30 s. GAPDH served as an internal control. The 2−ΔΔCt method was used to calculate relative expression levels of RNAs. Primers for quantitative real-time polymerase chain reaction (qRT-PCR) are listed in Supplementary Table S1.
Plasmid construction and cell transfection
Human CRC cell lines (SW480 and HT29) were incubated in six-well plates for 24 h before transfection. The short hairpin RNAs against ARAP1-AS1 (termed sh-ARAP1-AS1#1 and sh-ARAP1-AS1#2) and negative control (termed sh-NC), the pcDNA 3.1 vector specific to YY1 (termed YY1) and negative control (termed NC), and the short hairpin RNA against transcription factor YY1 (termed sh-YY1) and negative control (termed sh-NC) were designed and synthesized by the RiboBio Company (Guangzhou, China). Transfections were conducted and finished using Lipofectamine 2000 (Invitrogen). The interference sequences are listed in Supplementary Table S1.
CCK-8 assay
The transfected SW480 and HT29 cell lines were reaped for CCK-8 assay. After diluting to 5 × 103 cells/mL, cells were planted into 96-well plates. Cell proliferation was assessed using a Cell Counting Kit-8 kit (DOJINDO, Tokyo, Japan) following the user guide. Cells of each well were incubated with 10 μL of CCK-8 solution at 37°C for 2 h. At length, a microplate reader (Bio-Rad, Hercules, CA) was utilized to measure absorption at 450 nm.
Transwell assay
Transwell assay was used to measure the migratory and invasive abilities of indicated cells. In brief, cells (1 × 104 cells/well) were plated in 24-well plates that were placed into the Transwell chamber (8-mm pore size; Millipore, Eschborn, Germany). For invasion and migration assays, the upper chamber was precoated with or without 50 mL of Matrigel (BD Biosciences). After transfection and resuspension, 200-mL cell solutions were added into the upper chambers. Meanwhile, in the lower chamber, 500 mL of DMEM solution mixed with 10% FBS was added. Following incubation for 48 h, cells on the upper surface of the chamber were removed with cotton swabs, while cells on the lower surface were fixed with 4% paraformaldehyde (PFA) and stained with 0.1% crystal violet (Beyotime Institute of Biotechnology, Haimen, China) for 15 min. Finally, the cells that had invaded and migrated through the membrane were calculated under a microscope (Nikon, Tokyo, Japan) at a magnification of × 100.
Western blot analysis
Total proteins were lysed with RIPA lysis buffer (Beyotime Institute of Biotechnology). Protein concentrations were measured with the BCA Protein Assay Kit (Beyotime Institute of Biotechnology). Afterward, proteins (20 μg) were separated with sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene difluoride (PVDF) membrane. Subsequently, the membranes were blocked with Tris-buffered saline (TBS) containing 5% fat-free milk for 1 h at room temperature.
Afterward, membranes were incubated with specific primary antibodies: E-cadherin (1:1000 dilution), N-cadherin (1:1000 dilution), YY1 (1:1000 dilution), YY2 (1:1000 dilution), β-catenin (1:1000 dilution), C-myc (1:1000 dilution), cyclin D1 (1:1000 dilution), and GAPDH (1:1000 dilution) at 4°C overnight. All antibodies were bought from Abcam (Eugene, OR). Afterward, the appropriate secondary antibody conjugated with horseradish peroxidase (1:2000 dilution; Santa Cruz Co.) was incubated with the membranes for 4 h at room temperature. The expression level of the protein band was normalized to GAPDH.
Immunofluorescence
SW480 and HT29 cells (1 × 105 cells/well) were seeded on glass coverslips in six-well plates for 24 h and then fixed in 4% PFA at room temperature for 30 min. Subsequently, slides were permeabilized with PBS-T solution (mixed with 0.3% Triton X-100) for 15 min and blocked with PBS-B solution (mixed with 4% BSA) for 20 min. After consecutive incubations with the primary antibodies E-cadherin (1:1000 dilution) and N-cadherin (1:1000 dilution) overnight at 4°C, cells were incubated with the appropriate secondary antibody (1:2000 dilution) for 1 h at room temperature. DAPI (Beyotime Institute of Biotechnology, Shanghai, China) was used to stain nuclei for 30 min at room temperature. Finally, cells were visualized by fluorescence microscopy (Olympus, Tokyo, Japan).
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) assay was performed using the SimpleChIP® Enzymatic Chromatin IP Kit (No. 9003; CST, Boston, MA) in accordance with the manufacturer's introduction. Briefly, SW480 and HT29 cells were fixed with 1% formaldehyde and cross-linked at 37°C for 10 min, and then glycine was added to stop the cross-linking reaction. Subsequently, SW480 and HT29 cells were harvested and sonicated by enzymatic digestion. Afterward, lysates were immunoprecipitated with specific antibodies at 4°C overnight. IgG was used as a negative control (NC). After washing, elution, and decross-linking, the recovered DNA was quantified using qRT-PCR.
Luciferase reporter assay
The ARAP1-AS1 promoter region containing the binding sites (wild type or mutant type) was constructed into the pGL3 vector (Promega, Madison, WI) and cotransfected into SW480 and HT29 cells along with YY1 or NC. Forty-eight hours after incubation, a dual-luciferase reporter assay system (Promega) was used to measure luciferase activity. Renilla luciferase activity was used for normalization.
Statistical analyses
Each experiment was repeated independently three times. All data are presented as mean ± standard deviation, and statistical analyses were performed using statistics 20.0 (SPSS, Chicago, IL). Besides, Student's t-test was used to analyze the difference of two independent groups, while one-way analysis of variance was utilized to analyze differences among multiple groups. A value of p < 0.05 was considered to be statistically significant.
Results
ARAP1-AS1 was overexpressed in CRC tissues and cell lines
Based on the TCGA dataset, ARAP1-AS1 was highly expressed in CRC samples (Fig. 1A). Furthermore, its high expression was further validated in CRC tissues and collected from 82 CRC patients, identified, and compared with adjacent normal tissues (Fig. 1B). Based on the mean expression level of ARAP1-AS1 in 82 CRC tissues, all patients were classified into the ARAP1-AS1 high or low expression group. Then, the authors analyzed the association between the ARAP1-AS1 expression and clinicopathological features of CRC patients. As listed in Table 1, the expression level of ARAP1-AS1 correlated with the TNM stage and lymph node metastasis. In addition, ARAP1-AS1 was expressed higher in CRC tissues with metastasis rather than in those without metastasis (Fig. 1C). Next, the authors examined the expression level of ARAP1-AS1 in both CRC cells and normal colorectal epithelial cell. As a result, CRC cell lines exhibited a relatively higher expression level of ARAP1-AS1 (Fig. 1D).
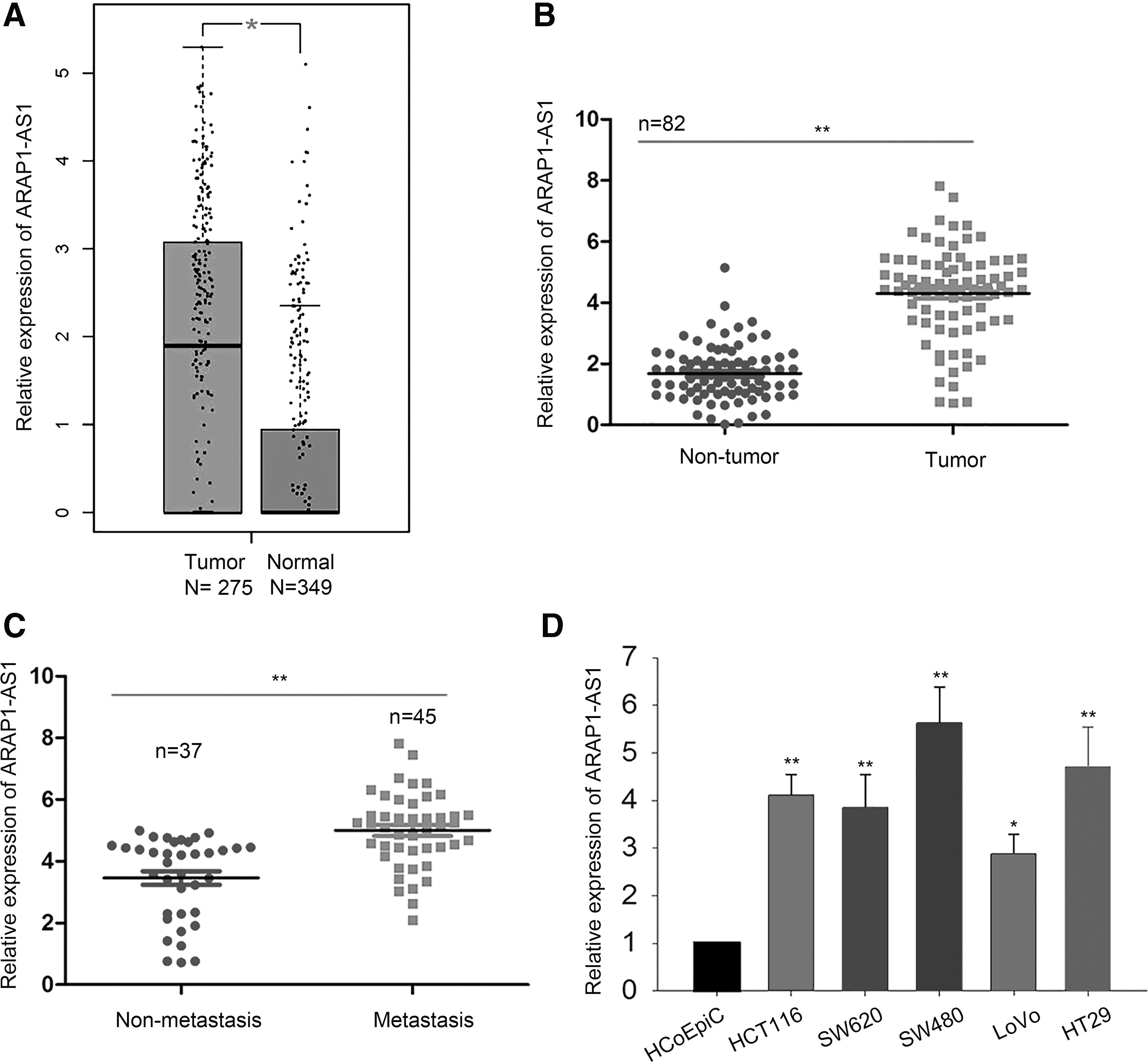
ARAP1-AS1 was overexpressed in CRC tissues and cell lines.
Correlation Between ARAP1 Antisense RNA 1 Expression and Clinical Features of Colorectal Cancer (n = 82)
Low/high by the sample median. Pearson χ 2 test.
p < 0.01 was considered statistically significant.
ARAP1-AS1, ARAP1 antisense RNA 1.
Knockdown of ARAP1-AS1 suppressed cell migration, invasion, and EMT process in CRC
To identify the biological role of ARAP1-AS1 in CRC, functional assays were conducted. According to data of Figure 1, SW480 and HT29 cells exhibited the highest level of ARAP1-AS1 compared with the other three cell lines. Therefore, the authors decreased the expression level of ARAP1-AS1 in SW480 and HT29 cells by transfecting with ARAP1-AS1-specific shRNAs (sh-ARAP1-AS1#1 and sh-ARAP1-AS1#2). qRT-PCR analysis showed that sh-ARAP1-AS1#2 most efficiently decreased the expression level of ARAP1-AS1 in two CRC cells (Supplementary Fig. S1A). Therefore, the authors chose sh-ARAP1-AS1#2 for subsequent experiments.
At first, cell viability was tested in ARAP1-AS1-downregulated CRC cells. It was found that knockdown of ARAP1-AS1 efficiently decreased CRC cell viability (Fig. 2A). Considering the association between ARAP1-AS1 expression and metastasis, the authors detected the effect of ARAP1-AS1 knockdown on CRC migration and invasion. According to results of Transwell assays, both migration and invasion were inhibited by ARAP1-AS1 knockdown (Fig. 2B). Moreover, EMT was detected in indicated CRC cells by measuring expression levels of EMT markers (E-cadherin and N-cadherin). ARAP1-AS1 knockdown increased the level of E-cadherin, but decreased the level of N-cadherin, indicating that ARAP1-AS1 reversed EMT progress (Fig. 2C, D). All these results suggested that ARAP1-AS1 promoted CRC cell migration and invasion.
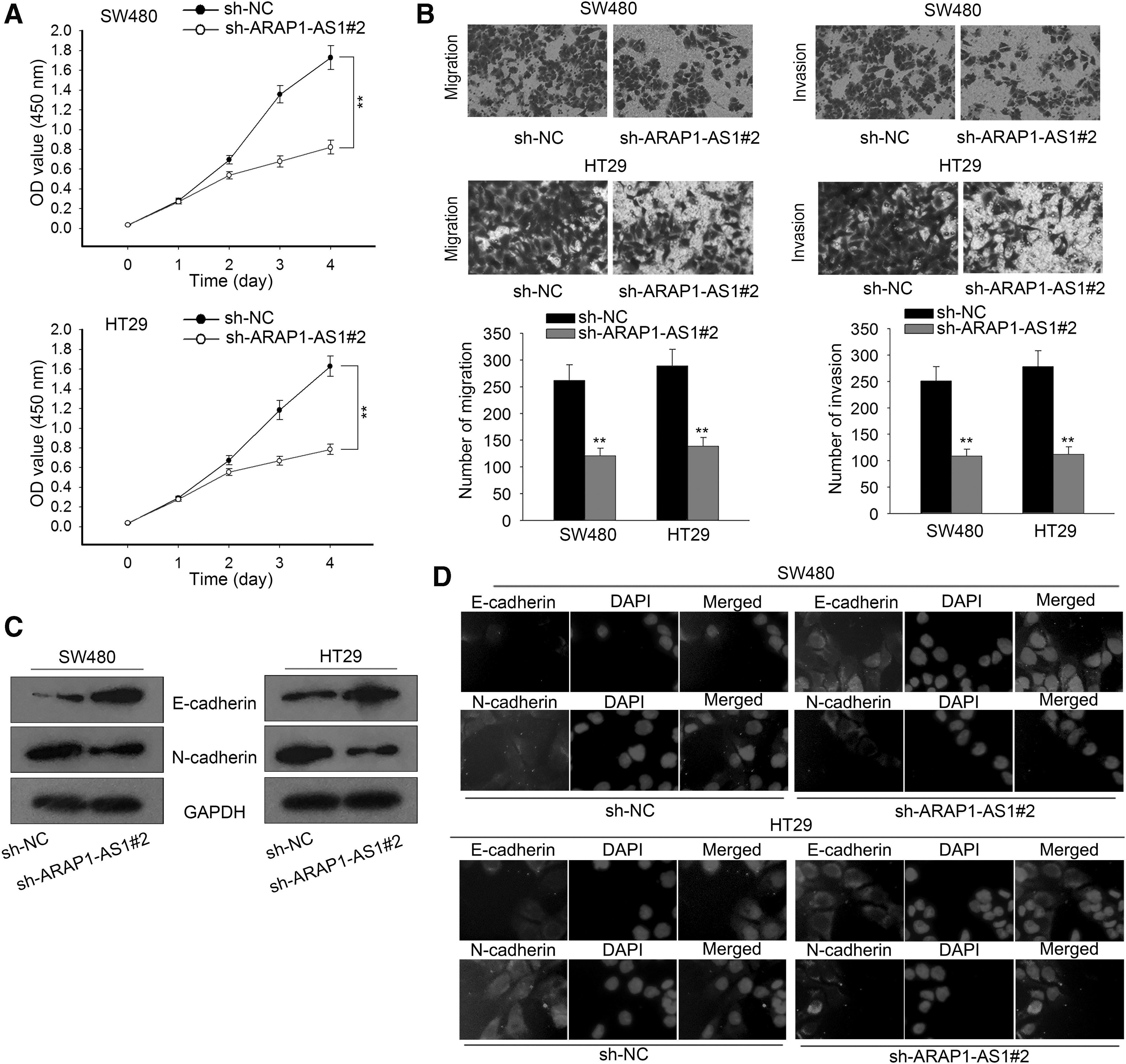
Knockdown of ARAP1-AS1 suppressed cell migration, invasion, and the EMT process in CRC.
YY1 activated the transcription of ARAP1-AS1
Based on the data above, the authors confirmed that upregulation of ARAP1-AS1 contributed to tumor progression in CRC. The authors further explored the molecular mechanism involved in upregulation of ARAP1-AS1 in CRC. Increasing evidence shows that lncRNAs can be activated by their upstream transcription factors. Therefore, the authors tried to explore the transcription activator of ARAP1-AS1 in CRC. According to ChIP-Seq results of UCSC (

YY1 activated the transcription of ARAP1-AS1.
Based on the position, these two binding sites were divided into two parts of the ARAP2-AS1 promoter. ChIP assay showed that part 2 (P2) of the ARAP1-AS1 promoter exhibited a strong binding affinity with YY1 (Fig. 3C). Furthermore, luciferase reporter assay indicated that P2 was responsible for transcription activation of YY1 on the ARAP1-AS1 promoter (Fig. 3D). Similarly, YY1 was upregulated in CRC tissues and cell lines (Fig. 3E). Correlation between the expression of YY1 and that of ARAP1-AS1 was found to be positive in CRC tissues (Supplementary Fig. S1B). To analyze whether YY1 regulated ARAP1-AS1 expression in CRC cells, the authors overexpressed or silenced YY1 in both SW480 and HT29 cells (Fig. 3F). The expression of ARAP1-AS1 was found to be positively regulated by YY1 in CRC cells (Fig. 3G).
Then, the authors also examined the messenger RNA (mRNA) and protein levels of YY2 in CRC cell lines and the normal cell line. Results showed that the expression level of YY2 was not significantly expressed in CRC cell lines compared with the normal cell line (Supplementary Fig. S1C, D). To demonstrate the monospecificity of YY1, the authors examined the protein band of YY1 by using a specific antibody. It was found that the position of YY1, but not YY2, is corresponding to this antibody (Supplementary Fig. S1E). Collectively, the authors confirmed that YY1 enhanced the expression of ARAP1-AS1 by promoting ARAP1-AS1 transcription.
YY1/ARAP1-AS1 axis promoted cell migration and EMT progress
To validate the biological function of the YY1/ARAP1-AS1 axis in CRC progression, rescue assays were conducted in SW480 cells. At first, the authors examined the expression of ARAP1-AS1 in cells cotransfected with sh-ARAP1-AS1 and YY1 expression vector. As show in Figure 4A, cell migration and invasion suppressed by sh-ARAP1-AS1 were partially recovered by YY1. Moreover, overexpression of YY1 reversed the effect of ARAP1-AS1 knockdown on the EMT process (Fig. 4B, C). Therefore, the authors confirmed that YY1 was involved in ARAP1-AS1-mediated CRC cell migration and the EMT process.
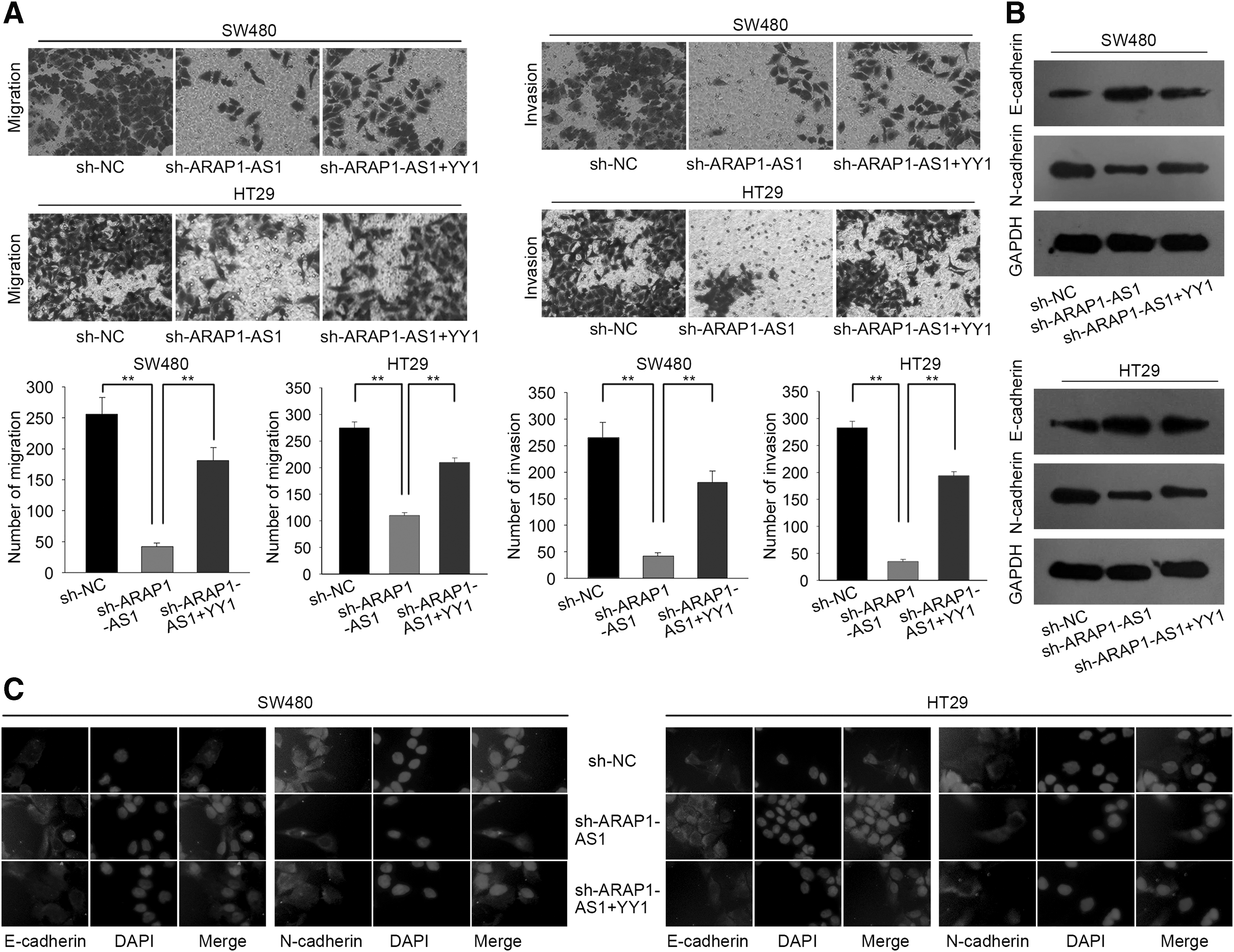
The YY1/ARAP1-AS1 axis promoted cell migration and EMT progress.
YY1/ARAP1-AS1 axis regulated the Wnt/β-catenin signaling pathway in CRC cells
It has been widely reported that the Wnt/β-catenin signaling pathway can be involved in lncRNA-mediated cancer progression. 29,30 In this study, the authors investigated whether the YY1/ARAP1-AS1 axis could regulate the Wnt/β-catenin signaling pathway in CRC cells.
Western blot analysis showed that protein levels of the core factors of the Wnt/β-catenin signaling pathway (β-catenin, c-myc, and cyclin D1) decreased by ARAP1-AS1 knockdown, but increased by YY1 overexpression (Fig. 5A). Moreover, overexpression of YY1 rescued the decreased levels in CRC cells transfected with sh-ARAP1-AS1. Functional assays were further performed to demonstrate the involvement of the Wnt/β-catenin signaling pathway in YY1/ARAP1-AS1-mediated CRC cell migration and invasion. As shown in Figure 5B, cell viability decreased by the knockdown of ARAP1-AS1 or YY1 was partially increased by the treatment with LiCl (activator of the Wnt/β-catenin signaling pathway). Similarly, both migration and invasion that were inhibited by sh-ARAP1-AS1 or sh-YY1 were recovered by introduction of LiCl (Fig. 5C, D). Therefore, the YY1/ARAP1-AS1 axis might promote CRC cell migration through the Wnt/β-catenin signaling pathway.
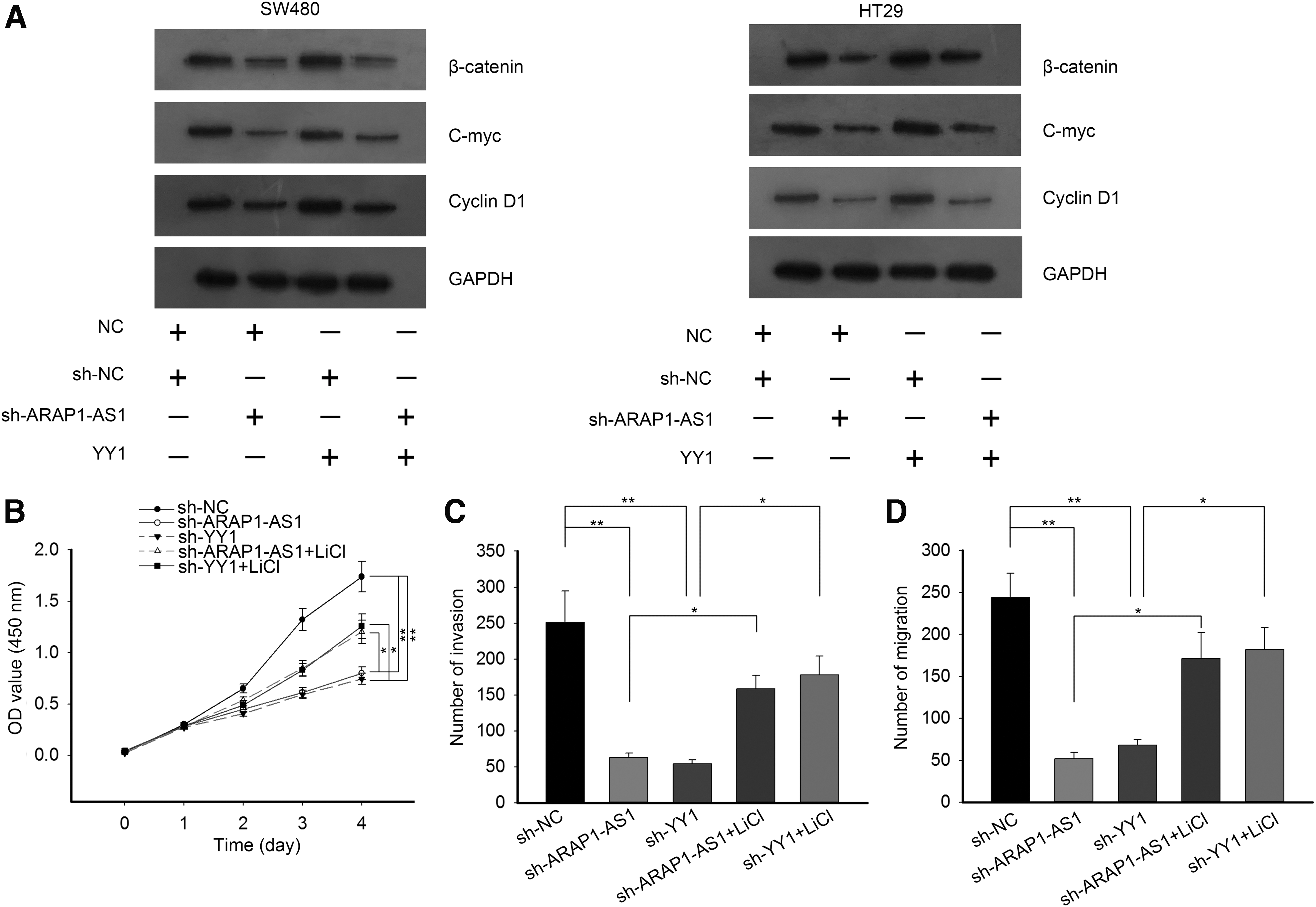
The YY1/ARAP1-AS1 axis regulated the Wnt/β-catenin signaling pathway in CRC cells.
Discussion
With the development of high-throughput techniques, numerous systematic cancer genomic projects have been applied in investigating various molecular pathways in human cancers. 31,32 According to TCGA data, ARAP1-AS1 was upregulated in CRC samples. Therefore, this study focused on the role of ARAP1-AS1 in CRC progression. Consistently, the expression of ARAP1-AS1 was found to be higher in 82 CRC tissues and 5 CRC cell lines compared with normal controls.
Dysregulation of lncRNAs is closely correlated with the abnormal biological behaviors in human cancers. Previous studies have shown that lncRNAs are crucial regulators for cancer cell migration and EMT progress. 33 –37 In the present study, the authors found upregulation of ARAP1-AS1 in CRC tissues and metastatic tissues, indicating the potential participation of ARAP1-AS1 in CRC cell migration and invasion. In this regard, the authors investigated whether ARAP1-AS1 regulated CRC cell migration, invasion, and EMT progress. The authors designed and conducted loss-of-function assays in SW480 and HT29 cell lines that exhibited the highest level of ARAP1-AS1. Results of loss-of-function assays demonstrated that knockdown of ARAP1-AS1 efficiently suppressed cell migration, invasion, and EMT progress, suggesting that ARAP1-AS1 acted as an oncogene in CRC by promoting cell migration, invasion, and EMT progress.
Previous reports have revealed that transcription factors can lead to dysregulation of lncRNAs in human cancers. 38 –40 Based on the bioinformatic analysis, the authors found some potential transcription factors of ARAP1-AS1. Among which, YY1 has been demonstrated to be a transcription activator of lncRNAs. 41 Therefore, the interaction between YY1 and ARAP1-AS1 was analyzed in this study. Based on mechanism investigation, YY1 activated ARAP1-AS1 transcription by binding to its promoter. More importantly, the authors found that the expression of ARAP1-AS1 was positively regulated by YY1 in CRC cells. These data suggested that YY1 is a transcriptional activator of ARAP1-AS1 in CRC. Rescue assays demonstrated the crucial role of YY1 in ARAP1-AS1-mediated CRC cell migration and the EMT process. All these findings indicated that YY1-induced upregulation of ARAP1-AS1 promoted cell migration, invasion, and EMT progress in CRC. In this study, the authors explored a novel YY1/ARAP1-AS1 axis in CRC progression.
The Wnt/β-catenin signaling pathway is closely associated with progression 42 –44 of human cancers, which can be involved in lncRNA-mediated tumor progression. 42 In the present study, the authors investigated potential regulation of the YY1/ARAP1-AS1 axis on the Wnt/β-catenin signaling pathway. The authors found positive regulation of YY1 and ARAP1-AS1 on Wnt/β-catenin signaling, indicating that the YY1/ARAP1-AS1 axis might promote CRC cell migration by activating the Wnt/β-catenin signaling pathway. By using LiCl (activator of the Wnt/β-catenin signaling pathway), rescue assays were carried out. It was found that the function of sh-ARAP1-AS1 or sh-YY1 in CRC cell migration and EMT progress was recovered by addition of LiCl. In conclusion, the study revealed that the YY1/ARAP1-AS1 axis promotes CRC cell migration and EMT progress through the Wnt/β-catenin signaling pathway. These findings may contribute to developing novel therapeutic targets for CRC.
Footnotes
Acknowledgment
The authors thank all the laboratory members.
Funding Information
This work was supported by the Wenzhou Science and Technology Planning Project (Y20120030).
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.