
Abstract
Cancer Biotherapy and Radiopharmaceuticals
officially retracts the paper entitled, “miR-132 Regulates Adriamycin Resistance in Colorectal Cancer Cells Through Targeting Extracellular Signal-Regulated Kinase 1,” by Yong Liu and Mei Zhang (Cancer Biother Radiopharm. 2019;34(6):398–404; doi: 10.1089/cbr.2018.2749) due to the discovery that the paper was submitted from a paper mill. This is a violation of the journal's standard protocols and is considered an infraction against the rigorous standards of scientific publishing.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves, and does not tolerate any misconduct.
Introduction
Colorectal cancer (CRC) is a common clinical malignant tumor of the digestive tract. 1 The incidence rates of CRC declined annually by ∼2%–3%, although the trend may have stabilized in women during the most recent data years from 2005 through 2014. 2 Reductions in CRC incidence before 2000 are attributed equally to changes in risk factors and the use of screening, which allows for the removal of premalignant lesions. 3 CRC shows the characteristics of occult onset, easy metastasis, and poor prognosis. Therefore, exploring the mechanism of the pathogenesis of CRC is important for helping early diagnosis, implementing individualized treatment, improving treatment effect, and elevating survival and prognosis.
Extracellular signal-regulated kinase 1 (ERK1) is an important protein in the ERK/mitogen-activated protein kinase (MAPK) signaling pathway. Increased ERK1 expression or functional activity plays a crucial role in the occurrence, progression, metastasis, and drug resistance of various tumors, such as CRC, 4 prostate cancer, 5 and breast cancer. 6 Numerous studies revealed that ERK1 expression and activity enhancement are related to CRC occurrence, progression, invasion, metastasis, and drug resistance, suggesting that abnormal elevation of ERK1 is a cancer-promoting factor in CRC. 7 –10
MicroRNAs are a type of endogenous small noncoding single-stranded RNA at the length of 22–25 nucleotides. They participate in the biological process by complete or incomplete complementary binding with the 3′-UTR of target gene to promote target gene mRNA degradation or inhibit mRNA translation. MicroRNA accounts for 1% of the human gene, while regulates the expression of more than one-third of the human genes. 11
The role of microRNA expression and dysfunction in tumorigenesis has received more and more attention. 12,13 It was exhibited that abnormal expression of miR-132 is closely related to the development and pathogenesis of CRC 14 –16 as demonstrated by association of miR-132 downregulation with tumor size, distant metastasis, and TNM stage in CRC patients as well as proliferation and invasion of CRC cells possibly through targeting anoctamin 1 or ZEB2. Bioinformatics analysis demonstrated the targeting relationship between miR-132 and ERK1 3′-UTR. This study investigated the role of miR-132 in regulating ERK1 expression and affecting CRC cell proliferation, apoptosis, and adriamycin (ADM) resistance.
Materials and Methods
Main reagents and materials
Human normal colorectal cell line NCM460 and CRC cell lines Lovo and SW480 were purchased from Beijing Beina Bio. HEK293T cell line was purchased from Shanghai Cell Bank of Chinese Academy of Sciences. RPMI 1640, Opti-MEM, and fetal bovine serum (FBS) were purchased from Gibco. Trizol and Lipofectamine 2000 were purchased from Invitrogen. PrimeScript RT reagent kit was purchased from Takara. miR-132 mimic and miR-NC were purchased from RiboBio. Rabbit antihuman ERK1 and β-actin antibodies were purchased from CST. Goat antirabbit immunoglobulin G (H + L) secondary antibody was purchased from Jackson ImmunoResearch. Cell counting kit-8 (CCK-8), BeyoECL Plus, and EdU staining kit were purchased from Beyotime. Annexin V/PI cell apoptosis detection kit was purchased from Shanghai Shengsheng Bio. Dual-Glo Luciferase Assay System and pMIR plasmid were purchased from Promega.
Cell culture
NCM460, Lovo, and SW480 cell line were maintained in RPMI1640 medium containing 10% FBS and cultured at 37°C with 5% CO2. The cells were passaged at 1:4. The cells in logarithmic phase were used for experiments.
ADM drug-resistant cell line establishment
Lovo cells in logarithmic phase were treated by ADM from 0.0125 μg/mL for 2 weeks. Then cells were kept growing and treated with increased concentration of ADM up to 0.2 μg/mL until stable growth in ADM. At last, the cells were stably passaged in ADM to obtain ADM-resistant CRC cell line Lovo/ADM.
SW480 cells in logarithmic phase were treated with ADM from 0.125 μg/mL for 2 weeks. Then cells were kept growing and treated with increased concentration of ADM up to 2.0 μg/mL until stable growth in ADM. At last, cells were stably passaged in ADM to obtain ADM-resistant CRC cell line SW480/ADM.
Lovo and Lovo/ADM cells were treated with different concentrations (0, 0.01, 0.1, 1, 10, and 100 μg/mL) of ADM for 48 h. SW480 and SW480/ADM cells were treated with different concentrations (0, 0.1, 1, 10, 100, and 1000 μg/mL) of ADM for 48 h. Next, the cells were added with CCK-8 to measure the absorbance value (A450). Inhibition rate = (1 − A450 in drug group)/A450 in control × 100%. IC50 was calculated using SPSS18.0 software. Resistance index (RI) = IC50 of drug-resistant cell/IC50 of parent cell.
Dual luciferase reporter gene assay
The PCR product of the ERK1 3′-UTR full-length fragment or mutant fragment was double-digested and then ligated into the pMIR vector. After sequencing, the plasmid was designated as pMIR-ERK1-WT and pMIR-ERK1-MUT. The HEK293T cells were transfected with pMIR-ERK1-WT (or pMIR-ERK1-MUT) together with miR-132 mimic (or miR-NC) by Lipofectamine 2000. After incubated for 48 h, luciferase activity was detected by Dual-Glo Luciferase Assay System kit according to the manual.
Cell transfection and grouping
Lovo/ADM and SW480/ADM cells were divided into miR-NC group and miR-132 mimic group. A total of 10 μL of Lipofectamine 2000, 50 nmoL miR-NC, and 50 nmoL miR-132 mimic were diluted with 100 μL serum-free Opti-MEM medium, and incubated for 5 min at room temperature, respectively. The mixture was added to the cell culture medium for 72 h. Finally, cells were collected for detection.
The cells were seeded into a six-well plate and treated with 0.2 or 2.0 μg/mL ADM for 48 h. Next, the cells were tested on Beckmann FC500 MCL flow cytometry to evaluate cell apoptosis.
The cells were digested and incubated with 10 μM EdU for 2 h. Next, the cells were treated with 0.2 or 2.0 μg/mL ADM for 48 h. Then the cell proliferation was determined by EdU staining kit according to the manual.
qRT-PCR
PrimeScript RT reagent kit was used to reverse transcribe RNA to cDNA. The reverse transcription system contained oligdT Primer (50 μM) 0.5 μL, Random 6 mers (100 μM) 0.5 μL, PrimeScript RT Enzyme Mix 0.5 μL, RNA 1.0 μg, 5 × PrimeScript Buffer 2 μL, and RNase Free H2O. The reverse transcription was performed at 37°C for 15 min and 85°C for 5 s. The PCR system was composed of SYBR Fast qPCR Mix 10.0 μL, forward primer (10 μM) 0.8 μL, reverse primer (10 μM) 0.8 μL, cDNA 2.0 μL, and RNase Free dH2O 6.4 μL. The reaction was performed on Bio-Rad CFX96 at 95°C for 10 min, followed by 40 cycles of 95°C for 10 s, 60°C for 20 s, and 72°C for 15 s.
Western blot
Total protein was extracted from the cells by RIPA. After quantified by the BCA method, a total of 40 μg protein was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membrane at 300 mA for 100 min. After being blocked by 5% skim milk at room temperature, the membrane was incubated with primary antibody at 4°C overnight (ERK1 1:1000 and β-actin 1:5000). After being washed by PBST, the membrane was further incubated with horseradish peroxidase conjugated secondary antibody at room temperature for 60 min (1:5000). At last, the membrane was treated with BeyoECL Plus chemiluminiscence reagent and developed.
Cell apoptosis detection
The cells were digested by enzyme and collected. After resuspending in 100 μL binding buffer, 5 μL Annexin V-fluorescein isothiocyanate and 5 μL PI were added to the cells and incubated at room temperature avoiding light for 15 min. Then the cells were evaluated by flow cytometry.
Cell proliferation detection
The cells were added with EdU solution at 10 μM in logarithmic phase. After incubation for 2 h, the cells were seeded for 48 h and digested by trypsin. After fixing in paraformaldehyde, the cells were incubated in 100 μL Triton X-100 at room temperature and in 500 μL reaction fluid at room temperature avoiding light for 30 min. Finally, the cell were tested on flow cytometry.
Statistical analysis
SPSS 18.0 software was applied for data analysis. Measurement data were presented as mean ± standard deviation and compared by t-test or one-way analysis of variance. p < 0.05 was depicted as significant difference.
Results
The targeted regulatory relationship between miR-132 and ERK1
Bioinformatics analysis showed the complementary binding site between miR-132 and the 3′-UTR of ERK1 mRNA (Fig. 1A). Dual luciferase reporter gene assay exhibited that miR-132 mimic transfection significantly reduced the relative luciferase activity in HEK293T cells transfected by pMIR-ERK1-WT but not by pMIR-ERK1-MUT, confirming that ERK1 was the target gene of miR-132 (Fig. 1B).
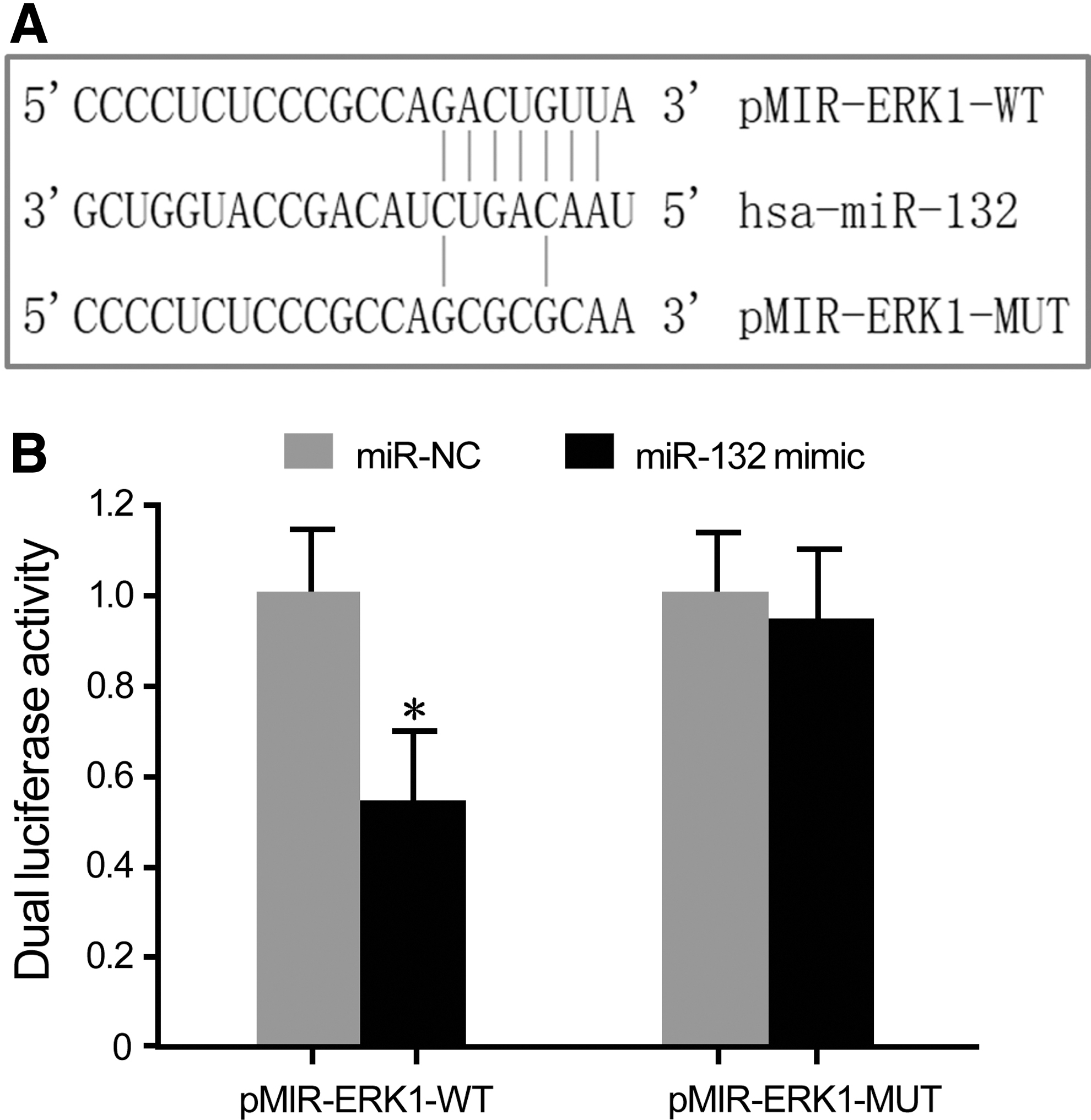
The targeted regulatory relationship between miR-132 and ERK1.
Drug-resistant CRC cells exhibited strong proliferation and drug resistance
CCK-8 assay showed that the proliferative activities of parental Lovo (Fig. 2A) and SW480 cells (Fig. 2B) were significantly lower than those of Lovo/ADM and SW480/ADM cells under the treatment of same dose of ADM. The IC50 of Lovo cells was 0.21 ± 0.04 μg/mL, whereas it was 2.76 ± 0.29 μg/mL in the drug-resistant Lovo/ADM cells, with the RI of Lovo/ADM cells being 13.14. The IC50 of SW480 cells was 6.22 ± 0.71 μg/mL, and the IC50 of SW480/ADM cells was 59.85 ± 4.19 μg/mL, with the RI of SW480/ADM cells being 9.62.
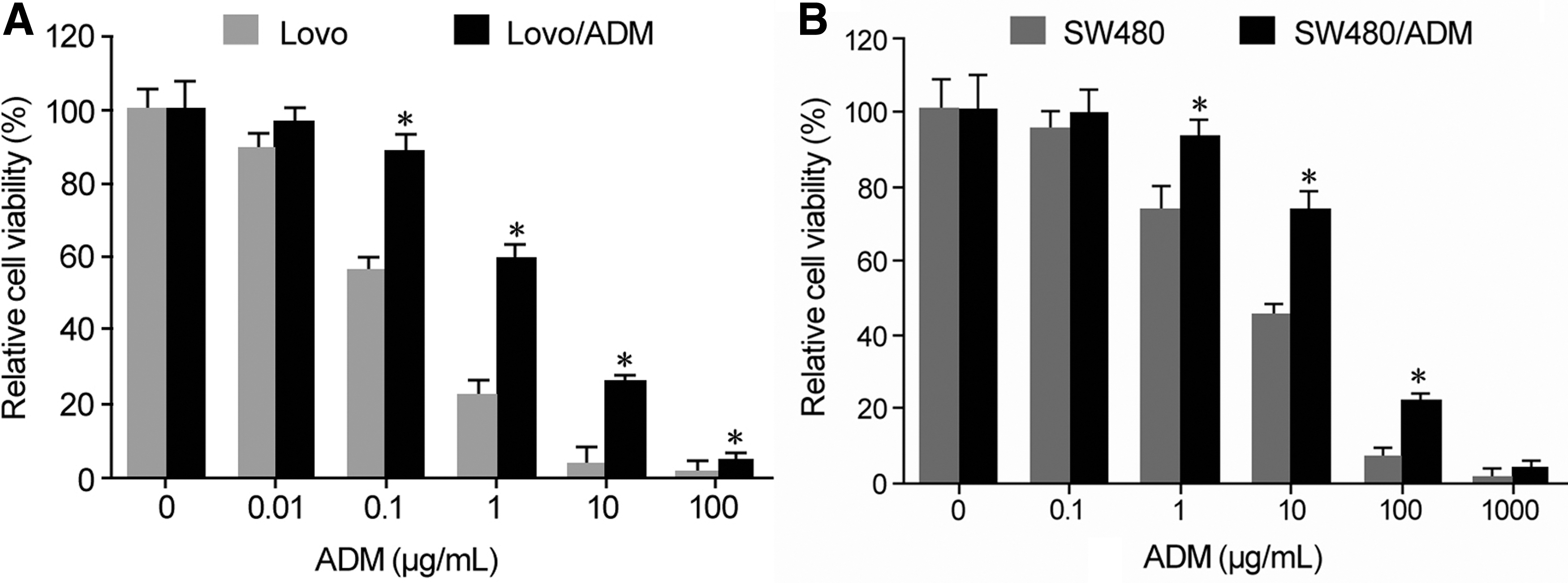
Drug-resistant CRC cells exhibited strong proliferation and drug resistance.
miR-132 reduced, whereas ERK1 upregulated in drug-resistant cells
qRT-PCR demonstrated that compared with human normal colorectal NCM460 cells, miR-132 expression was significantly decreased in CRC cells, and its level was significantly lower in drug-resistant cells than that of parental CRC cells (Fig. 3A). qRT-PCR revealed that the ERK1 mRNA expression in CRC cells was significantly higher than that in NCM460 cells, and it was significantly higher in CRC-resistant cells than in the parental cells (Fig. 3B). Western blot exhibited that compared with NCM460 cells, ERK1 protein expression in CRC cells was significantly upregulated, and its level in CRC drug-resistant cells obviously enhanced (Fig. 3C).
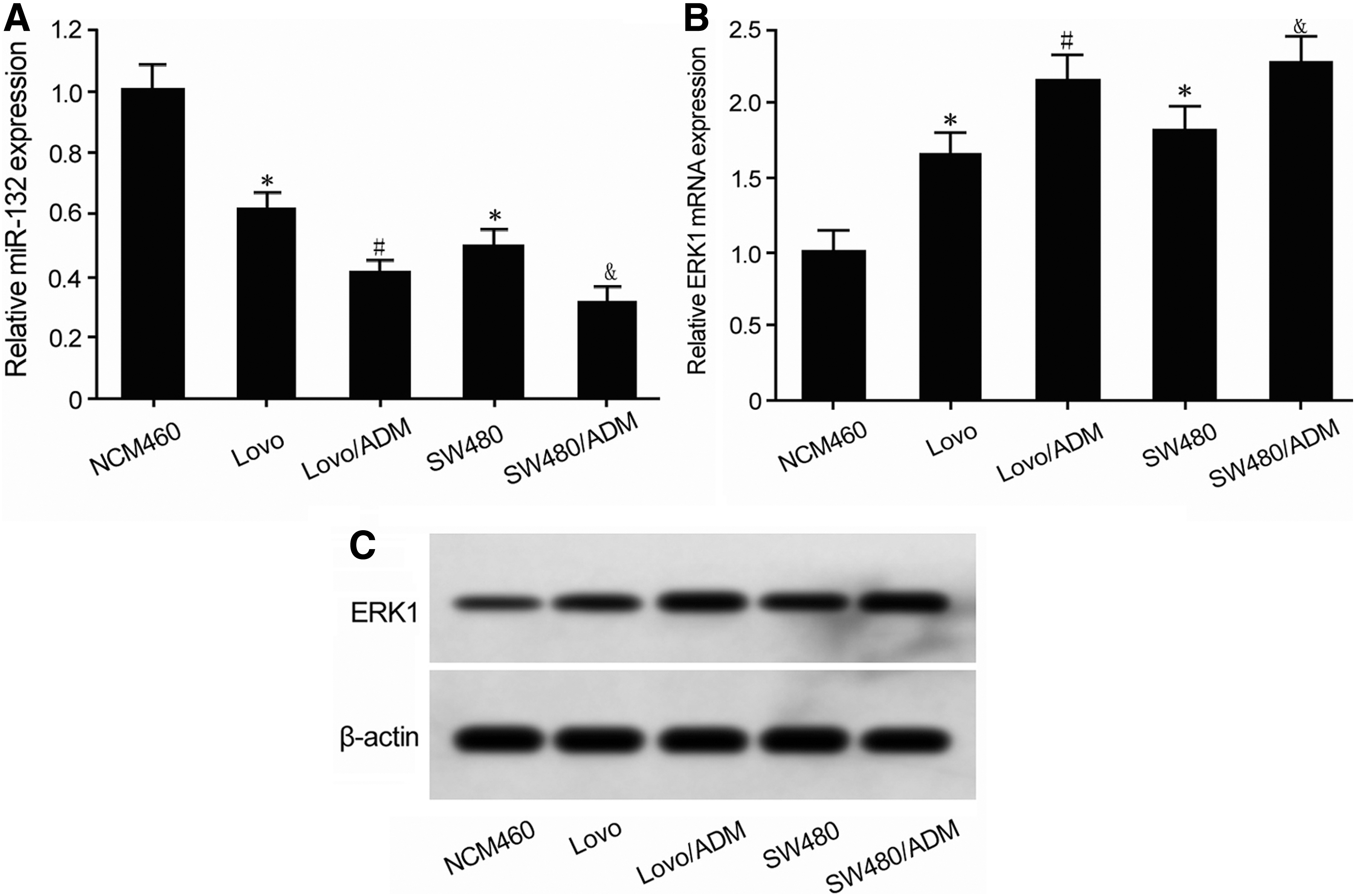
miR-132 reduced, whereas ERK1 upregulated in drug-resistant cells.
miR-132 overexpression promoted CRC cell apoptosis and reduced ADM resistance
qRT-PCR showed that miR-132 mimic transfection significantly upregulated miR-132 expression and declined ERK1 mRNA level in Lovo/ADM and SW480/ADM cells compared with miR-NC group (Fig. 4A, B). Western blot demonstrated that miR-132 mimic transfection reduced ERK1 protein level in Lovo/ADM and SW480/ADM cells (Fig. 4C). Flow cytometry revealed that transfection of miR-132 mimic markedly enhanced cell apoptosis, whereas inhibited cell proliferation in Lovo/ADM and SW480/ADM cells (Fig. 4D, E).
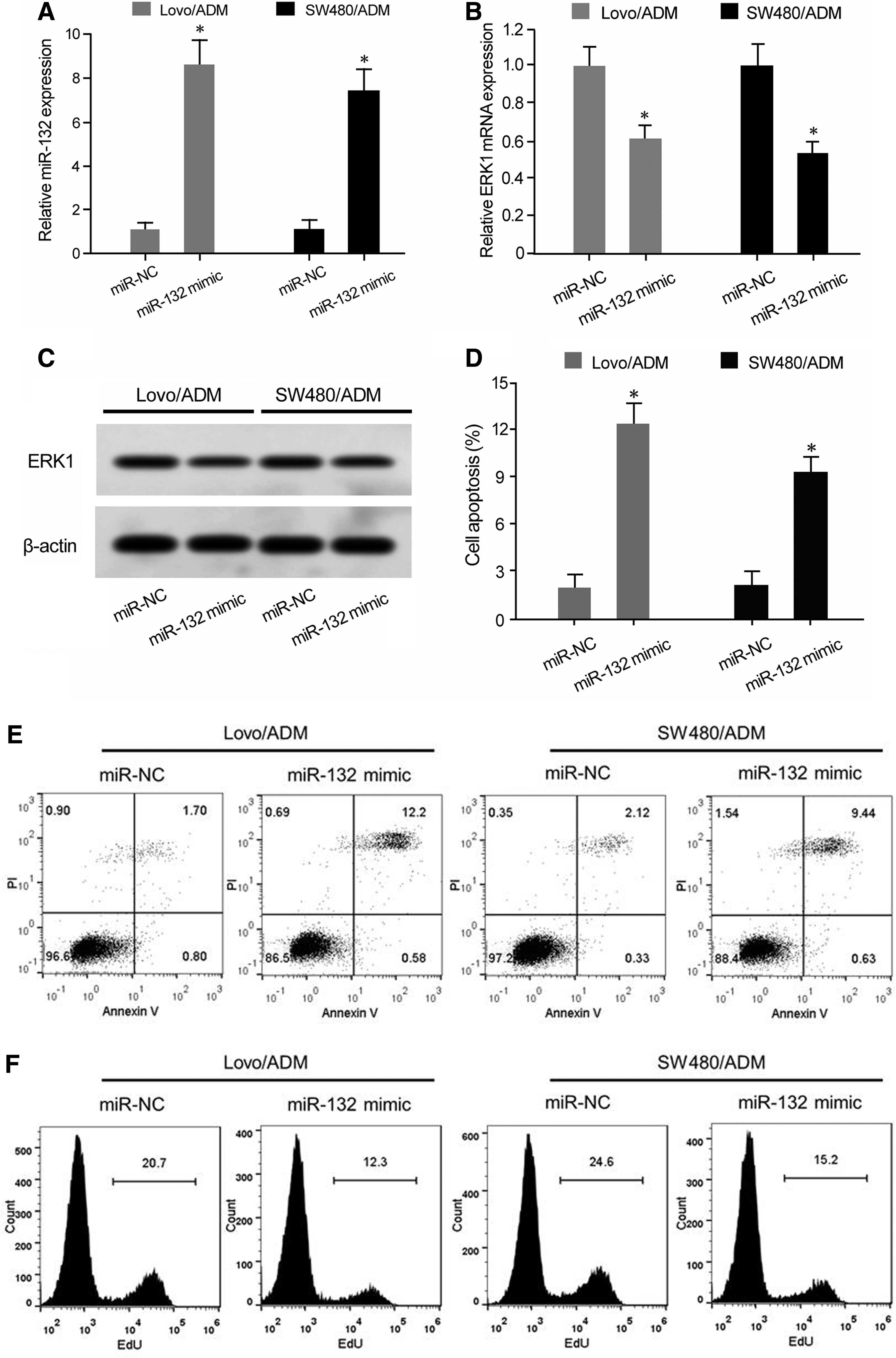
miR-132 overexpression promoted CRC cell apoptosis and reduced ADM resistance.
Discussion
ERK/MAPK signaling pathway is widely expressed in various tissues and cells, and can regulate a variety of biological processes, such as cell proliferation, cycle, apoptosis, migration, and invasion. 17,18 ERK1 is a silk/threonine kinase that is one of the important subtypes of ERK protein. After receiving the upstream cascade signal, ERK1 can phosphorylate the cytoplasmic protein and translocate into the nucleus. It further regulates various nuclear transcription factors, such as c-fos and c-Jun, and thus participates in the regulation of cell proliferation and apoptosis. 19,20 The expression and functional activity enhancement of ERK1 play an important role in regulating the progression, metastasis, and drug resistance of ovarian cancer, lung cancer, and breast cancer. 21 –24 A number of studies reported that ERK1 expression and activity enhancement are associated with the pathogenesis, progression, invasion, metastasis, and drug resistance of CRC, suggesting that abnormal elevation of ERK1 is a cancer-promoting factor in CRC. 5 –8
miR-132 is found to be decreased in multiple tumors, such as ovarian cancer, 25 lung cancer, 26 and thyroid cancer. 27 It was shown that reduced expression of miR-132 is associated with CRC occurrence, progression, and metastasis, thus is a tumor suppressor gene in CRC. 14 –16 Bioinformatics analysis demonstrated the targeting relationship between miR-132 and ERK1 3′-UTR. This study investigated the role of miR-132 in regulating ERK1 expression and affecting CRC cell proliferation, apoptosis, and doxorubicin (ADM) resistance.
In this study, dual luciferase reporter gene assay exhibited that miR-132 mimic transfection significantly reduced the relative luciferase activity in HEK293T cells transfected by pMIR-ERK1-WT but not by pMIR-ERK1-MUT, confirming the targeted relationship between ERK1 and miR-132. CCK-8 assay showed that the proliferative activities of parental Lovo and SW480 cells were significantly lower than those of Lovo/ADM and SW480/ADM cells under the treatment of same dose of ADM, indicating that the suppressing effect of ADM on drug-resistant cells was attenuated. According to the results of CCK-8 assay, the IC50 of drug-resistant CRC cells was significantly higher than that of parental cells, revealing that the ADM-resistant CRC cell line was successfully established. It was demonstrated that compared with significantly lower miR-132 expression, ERK1 level was significantly higher in drug-resistant cells than that of parental CRC cells. Moreover, miR-132 decreased, whereas ERK1 was upregulated in both drug-resistant and drug-sensitive CRC cells compared with normal colorectal NCM460 cells. It was shown that miR-132 reduction was associated with increased ERK1 expression. Their abnormal expression levels were not only related to the occurrence and malignant biological characteristics of CRC, but also involved in the regulation of drug resistance in CRC cells. Liu et al. 28 showed that miR-132 was significantly decreased in CRC, which was caused by the upregulation of LncRNA MIAT. Mokutani et al. 14 demonstrated that miR-132 was obviously declined in tumor tissues of patients with liver metastatic CRC compared with tumor tissues of patients without liver metastasis. Furthermore, lower miR-132 expression resulted in lower the disease-free survival rate (p = 0.0220) and overall survival rate of CRC patients (p = 0.0021). Qin et al. 15 revealed that the expression of miR-132 in CRC SW620, HT-29, SW480, HCT116, HCT-8, and Lovo cells was markedly lower than that of normal colorectal FHC cells. The expression level of miR-132 was also apparently lower in tumor tissues of clinical samples than in its corresponding adjacent tissues. Zheng et al. 16 exhibited that compared with normal intestinal epithelial NCM460 cells, the expression of miR-132 in CRC HT-29, HCT116, SW480, and SW620 cells was significantly decreased. Compared with CRC patients with no distant metastasis, the expression of miR-132 was significantly decreased in tumor tissue with distant metastasis. miR-132 expression was related to tumor size, distant metastasis, TNM stage, survival, and prognosis. All of the mentioned studies showed that miR-132 functions as a tumor suppressor gene in CRC.
At present, there are few studies on the relationship between miR-132 and drug resistance of CRC cells. Therefore, this study further explored whether miR-132 affects ADM resistance in CRC cells by regulating ERK1. It was observed that transfection of miR-132 mimic significantly reduced the expression of ERK1, enhanced cell apoptosis, attenuated cell proliferation, and reduced ADM resistance in drug-resistant Lovo/ADM and SW480/ADM cells. Liu et al. 28 found that transfection of miR-132 inhibitor significantly promoted the proliferation of CRC LoVo and SW480 cells, and significantly restrained cell apoptosis. It plays a role as a tumor suppressor gene in CRC. Mokutani et al. 14 showed that transfection of miR-132 mimic significantly inhibited cell proliferation, attenuated clonality and invasion in DLD-1 and HCT116 cells through targeted regulating ANO1 gene. Qin et al. 15 revealed that transfection of miR-132 mimic can significantly reduce the invasive ability of CRC Lovo and SW480 cells by targeted inhibition of paxillin gene. Song et al. 29 reported that LncRNA XIST promoted cell proliferation and regulated cell cycle by targeting the tumor suppressor gene miR-132. Zheng et al. 16 showed that overexpression of miR-132 significantly attenuated the process of epithelial–mesenchymal transition (EMT) and invasion of CRC cells, whereas downregulation of miR-132 expression enhanced EMT process and invasion by targeting the ZEB2 gene. This study combined the targeted regulatory relationship between miR-132 and ERK1, revealing that miR-132 plays a role in targeting inhibition of ERK1 expression to promote cell apoptosis and attenuate ADM resistance. However, whether miR-132 regulating ERK1 is related to the drug resistance of CRC patients is still unclear and requires further investigations.
Conclusions
miR-132 reduction and ERK1 elevation are related to ADM resistance in CRC cells. Upregulation of miR-132 expression inhibits CRC cell proliferation, induces apoptosis, and reduces ADM resistance possibly by targeting ERK1 expression.
Footnotes
Acknowledgment
This work was supported by Yan'an Science and Technology Plan Project Mechanism and clinical application of cadherin in the development of domestic gastric cancer (Grant No. 2012KW_05).
Disclosure Statement
There are no existing financial conflicts.