
Abstract
Objective:
Aberrantly expressed microRNAs (miRs) have associated with the development and progression of osteosarcoma (OS). In this study, the authors aimed to investigate the biological function of miR-5195-3p and the underlying mechanisms.
Methods:
Quantitative real-time polymerase chain reaction analysis was performed to determine the expression of miR-5195-3p in OS tissues and cell lines. Then, two OS cell lines (MG-63 and U2OS) were transfected with miR-5195-3p mimics to obtain stably miR-5195-3p overexpression cell lines. A series of functional assays, including Cell Counting Kit-8 assay, colony formation assay, flow cytometry assay, and Hoechst staining were performed to analyze cell proliferation and apoptosis.
Results:
The authors first observed downregulation of miR-5195-3p in OS tissues and cell lines. A series of functional assays demonstrated that miR-5195-3p overexpression significantly attenuated OS cell proliferative activity and induced apoptosis. At a molecular level, the neural precursor cell which expressed developmentally downregulated protein 9 (NEDD9), was inversely correlated with the expression level of miR-5195-3p. Furthermore, ectopic expression of NEDD9 counteracted the antiproliferative and apoptotic effects of miR-5195-3p overexpression in OS cells.
Conclusions:
In summary, the miR-5195-3p/NEDD9 axis may be a promising antitumor agent for OS.
Introduction
Osteosarcoma (OS), the most common primary bone tumor, originated from osteoblasts or their precursors, which is frequently seen in children, adolescents, and young adults. 1 OS represents 0.1% of the United States incidence burden, according to the American Cancer Society and their morbidity continues to slightly increase in recent years. 2,3 The adoption of modern, multiagent, and dose-intensive chemotherapy enormously improves the prognosis for patients with localized OS, with reports of long-term survival rates increased from 20% to 60%–70%. 4 However, management of patients with recurrent and metastatic is yet unsolved due to the malignant biological behaviors of OS. 5 Therefore, understanding the molecular mechanisms underlying the pathobiology may provide new clues for exploring therapeutic opportunities.
Small noncoding RNAs (19–23 nucleotides) named miRs, are conserved posttranscriptional modulators of gene expression in diverse biological processes. 6 miRs function in feedback mechanisms through ensuring various fundamental procedures, including cell proliferation, inflammation, apoptosis, and several metabolic functions under normal physiologic conditions. 7 Each miR can regulate hundreds of protein-encoding genes by binding to the 3′UTR regions of their target mRNAs with complementary paring. 8
Recent advances have highlighted the critical roles of numerous miRs in OS tumorigenesis. For instance, miR-543 is associated with OS cell proliferation and glycolysis. 9 miR-646 is associated with cell metastasis, 10 and miR-143 is correlated with apoptosis. 11 miR-5195-3p is a newly discovered member of the miRs family. In nonsmall cell lung cancer, the tumor suppressor miR-5195-3p prevents cell growth, migration, and invasion by suppressing the activities of Myosin VI (MYO6). 12 By targeting the Krüppel-like factor 5 gene (KLF5), enforced expression of miR-5195-3p could also exert tumor-suppressive effects in bladder cancer. 13 However, the biological behavior of miR-5195-3p in OS still remains unclear.
Neural precursor cell expressed developmentally downregulated protein 9 (NEDD9, also called HEF1 and CAS-L) was originally identified to be downregulated in mouse brain. 14 As a scaffold protein with focal adhesion, NEDD9 assembles signal transduction networks to response to growth factors, hormones, and extracellular matrix microenvironment. 15 NEDD9 belongs to a family of Crk-associated substrate (CAS) that involved in oncogenetic signaling in multiple solid and hematologic tumors. 16
Based on early identification of NEDD9 as components of the integrin-dependent signaling cascades, numerous subsequent studies and evidence support its roles in cancer biology through implementing these functions. 17,18 Recent studies provide mechanistic insight into the impact of miRs/NEDD9 axis in tumor progression and development. 19 –21 Zheng et al. 19 found that miR-125-5p inhibits lung adenocarcinoma cell proliferation and survival by negatively regulating NEDD9. NEDD9 is responsible for miR-145 overexpression-mediated suppression of tumorigenesis in pancreatic cancer 20 and renal cell carcinoma. 21
In the present study, the expression patterns of miR-5195-3p were analyzed in OS tissues and cell lines. By using bioinformatics tools, the authors predicted that NEDD9 might be a target of miR-5195-3p. The pivotal effect of the crosstalk between miR-5195-3p and NEDD9 in the regulation of OS cells proliferation and apoptosis was further investigated.
Materials and Methods
Tissue sample collection
A total of 26 paired fresh surgically cancerous and adjacent noncancerous (nontumor [NT]) tissues were collected from OS patients (mean age: 55 years; range, 43–61 years, 10 female, and 16 male) after diagnosed by 2 independent pathologist at the institute of orthopedics, Electric Power Hospital of Shanghai (Shanghai, China). All tissues samples were snap-frozen in liquid nitrogen and stored at −80°C. Written informed consent for the study was provided by all participants and this research was approved by the Research Ethics Committee of the Shanghai Electric Power Hospital.
Cell culture and transfection
Three OS cell lines (U2OS, MG-63, Saos-2, and SW1353) and normal human osteoblastic cell line hFOB 1.19 were obtained from the American Type Culture Collection (ATCC, Manassas, VA) and grown in Dulbecco's modified Eagle's medium (Gibco, Grand Island) with 10% heat-inactivated fetal bovine serum, penicillin (100 U/mL), and streptomycin (100 μg/mL). All cells were maintained in a humidified atmosphere containing 5% CO2 at 37°C.
For cell transfection, MG-63 and U2OS cells were seeded onto 12-well plates and transfected with miR-5195-3p mimics or corresponding miRNA negative control (miR-NC) (all purchased from Sangon Biotechnology Co. Ltd., China) using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to whatever instructions were supplied. The full-length cDNA from NEDD9 was cloned into the pcDNA3.1 plasmid (Genepharma, Shanghai, China) to produce pcDNA3.1-NEDD9 vector (NEDD9). To perform rescue experiments, they cotransfected miR-5195-3p mimics and NEDD9 or an empty vector into MG-3 cells.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNAs were extracted using TRIzol reagent (Takara Biotechnology, Co., Ltd., Dalian, China), then the products were reversed transcribed with PrimeScript RT Reagent kit (Takara Biotechnology, Co., Ltd) to generate cDNA according to the manufacturer's instructions. To determine miR-5195-3p expression pattern in OS cells and/or tissues, they assessed by ABI 7500 Real-Time PCR System (Applied Biosystems) using TaqMan MicroRNA assays (Applied Biosystems, Carlsbad, CA) and its relative expression was normalized to U6 small nuclear RNA. For NEDD9 mRNA analysis, cDNA was amplified using SYBR Green Real-Time PCR Master Mix (Toyobo, Osaka, Japan) with primers (Forward: 5′-GAGCTGGATGGATGACTACGA-3′ and reverse: 5′-AGCTCTTTCTGTTGCCTCTCA-3′). The relative expression of NEDD9 was normalized against GAPDH. The quantitative real-time polymerase chain reaction (PCR) was performed according to the following thermocycling conditions: 50°C for 2 min, followed by 40 amplification cycles at 95°C for 15 s and 60°C for 1 min. Relative expression of miR-5195-3p or NEDD9 was calculated using the 2−ΔΔCt method.
Cell Counting Kit-8 assay
Measurement by Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Kumamoto, Japan) was used to determine the cell proliferation between different groups. In brief, transfected cells were seeded into 96-well plates at the appropriate cell density (2 × 103 cells per well) and incubated in complete medium. At the 24, 48, 72, and 96 h time point, CCK-8 solution (10 μL per well) was added into each well and further incubated for another 2 h. The cell proliferation curve was plotted according to optical density values at 450 nm using a microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA).
Colony formation assay
The colony formation assay was performed to address the question “Whether miR-5195-3p impact on OS proliferation.” For each well of a six-well plate, 500 transfected cells per well were incubated for 2 weeks. Afterward, the cells were washed, fixed with 4% paraformaldehyde, and stained with 0.1% crystal violet dye for 20 min. Subsequently, the colonies were observed and counted under a light microscope (magnification, 200 × ).
Detection of apoptosis
Assessment of cell apoptosis was performed by flow cytometry using an Annexin V/propidium iodide (PI) double staining assay. In brief, transfected cells were collected by trypsinization and washed twice in phosphate-buffered saline (PBS). Then, cells were incubated with 500 μL binding buffer solutions containing 5 μL Annexin V·FITC and 5 μL PI (BD Biosciences, Franklin Lakes, NJ) for 20 min at room temperature in the dark. The percentage of early (Annexin V+/PI−) and late (Annexin V+/PI+) apoptotic cells were acquired using FACS Calibur flow cytometry (BD Biosciences) equipped with FlowJo 10.0 software (FlowJo LLC, Ashland, OR).
Hoechst staining
Transfected MG-63 and U2OS cells were stained with Hoechst Kit (Beyotime Institute of Biotechnology, Shanghai, China) according to the manufacturer's instructions to morphologic analysis of apoptosis. In brief, these cells were collected and fixed in 50 μL cold 4% formaldehyde for 30 min. Following washes with PBS three times, cells were stained with Hoechst 33258 (10 μg/mL) for 15 min. Nuclear morphologic changes were examined under a fluorescence microscope (Olympus, Tokyo, Japan). Strong blue fluorescence presented apoptotic cells.
Prediction of miR-5195-3p targets
The potential target genes of miR-5195-3p were predicted by using three bioinformatics tools (TargetScan program,
Plasmid construction and dual-luciferase reporter assay
The dual-luciferase reporter assay was performed to examine whether NEDD9 is a direct target of miR-5195-3p. In brief, the 3′UTR of NEDD9 that contains the putative binding sites for miR-5195-3p was amplified by PCR and cloned into the psiCHECK-2 receptor vector (Promega Corporation, Madison, WI) to generate wild-type reporter constructs (WT-3′UTR-NEDD9). Meanwhile, the PCR-amplified NEDD9 with the putative miR-5195-3p binding site mutations were inserted into the psiCHECK-2 receptor vector to obtain mutant reporter constructs (MUT-3′UTR-NEDD9). Subsequently, MG-3 or U2OS cells were cotransfected with 50 nM miR-5195-3p mimics or miR-NC and 100 ng WT-3′UTR-NEDD9 or MUT-3′UTR-NEDD9 with Lipofectamine 2000 Transfection Reagent according to the manufacturer's instructions. After 48 h transfection, the luciferase activity was examined by using a Promega Dual-Luciferase Reporter assay system and the relative Renilla/Firefly luciferase activity.
Western blot analysis
Proteins in OS cells were extracted by using a radioimmunoprecipitation assay lysis buffer and quantified with a bicinochoninc acid assay (Beyotime Institute of Biotechnology) following the manufacturers' instruction. Equal protein samples (30 μg) were separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel and then transferred to polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA). Then membranes were blocked with 5% skimmed milk in PBS at room temperature for 2 h and then incubated with primary antibodies against NEDD9 (1:1000, ab88584; Abcam) and GAPDH (1:5000, ab14247; Abcam) overnight at 4°C. Subsequently, the proteins were detected by using BeyoECL Plus kit (Beyotime Institute of Biotechnology) following incubation with horseradish peroxidase-conjugated secondary antibodies for 2 h at room temperature.
Statistical analyses
All statistical analyses were carried out using the SPSS 17.0 software tool (IBM Corp., Armonk, NY). Quantitative data are expressed as mean ± standard deviation of at least three experiments. Student's t-test or one-way analysis of variance with Dunnett's post hoc test was used to evaluate differences between two groups or more than two groups, respectively. Pearson's correlation test was performed to determine the correlation between NEDD9 and miR-5195-3p expression in OS tissues. Only p-values with less than 0.05 were considered to be statistically significant.
Results
miR-5195-3p was low-expressed in OS tissues and cell lines
First, quantitative real-time PCR was used to measure the expression of miR-5195-3p in OS tissues and adjacent NT tissues. As revealed in Figure 1A, miR-5195-3p expression was low-expressed in tumor samples from 26 tissues compared with expressions in adjacent, noncancerous samples (p < 0.001).

The expression pattern of miR-5195-3p in OS tissues and cell lines.
In accordance with clinical data, the expression level of miR-5195-3p was then measured in all the four OS cell lines (U2OS, MG-63, Saos-2, and SW1353) and normal human osteoblastic cell line hFOB 1.19, and this analysis confirmed that miR-51954-3p is indeed differentially expressed (Fig. 1B, p < 0.01, p < 0.001). These data imply that miR-5195-3p is low-expressed in both OS tissues and cell lines, and might play crucial roles in the initiation and progression of OS.
Overexpression of miR-5195-3p suppressed cell proliferation and promoted apoptosis in OS cells
Next, the authors investigated the potential biological functions of miR-5195-3p in OS in vitro by performing gain-of-function assays. Among the four OS cell lines, MG-63 and U2OS exhibited the lowest level of miR-5195-3p (Fig. 1B), therefore, both types of cells were transfected with miR-5195-3p mimics or miR-NC to construct miR-5195-3p overexpressing cell model. As demonstrated in Figure 2A, quantitative real-time PCR analysis showed that miR-5195-3p expression levels were significantly higher in these two kinds of OS cells transfected with miR-5195-3p mimics than cells transfected with a control, miR-NC (p < 0.001). The results from CCK-8 assay revealed impairment of proliferative capacity in miR-5195-3p mimics-transfected MG-63 and U2OS cells compared to miR-NC transfected cells (Fig. 2B, p < 0.001). Consistently, the number of colonies formed was remarkably reduced after miR-5195-3p overexpression in MG-63 (p < 0.01) and U2OS (p < 0.001) cells by cell colony formation assays (Fig. 2C). Moreover, they analyzed cell apoptosis using flow cytometry analysis. From Figure 3A, they can see that the percentage of early apoptotic cells (Annexin V+/PI−) in miR-5195-3p mimics group was significantly elevated compared with that in miR-NC group in MG-63 (10.9% ± 0.41% vs. 1.80% ± 0.12%, p < 0.001) and U2OS cells (6.60% ± 0.10% vs. 2.30% ± 0.11%, p < 0.01). Accordingly, the percentage of late apoptotic cells (Annexin V+/PI+) presented similar trends. In addition, miR-5195-3p mimics-treated MG-63 and U2OS cells displayed more condensed chromatin and fragmented nuclei (Fig. 3B), which are typical morphologic features of apoptotic cells, while in contrast, miR-NC-treated MG-63 and U2OS cells had no obvious morphologic signs of apoptosis. These data suggest that miR-5195-3p might function as a tumor suppressor in OS cells.
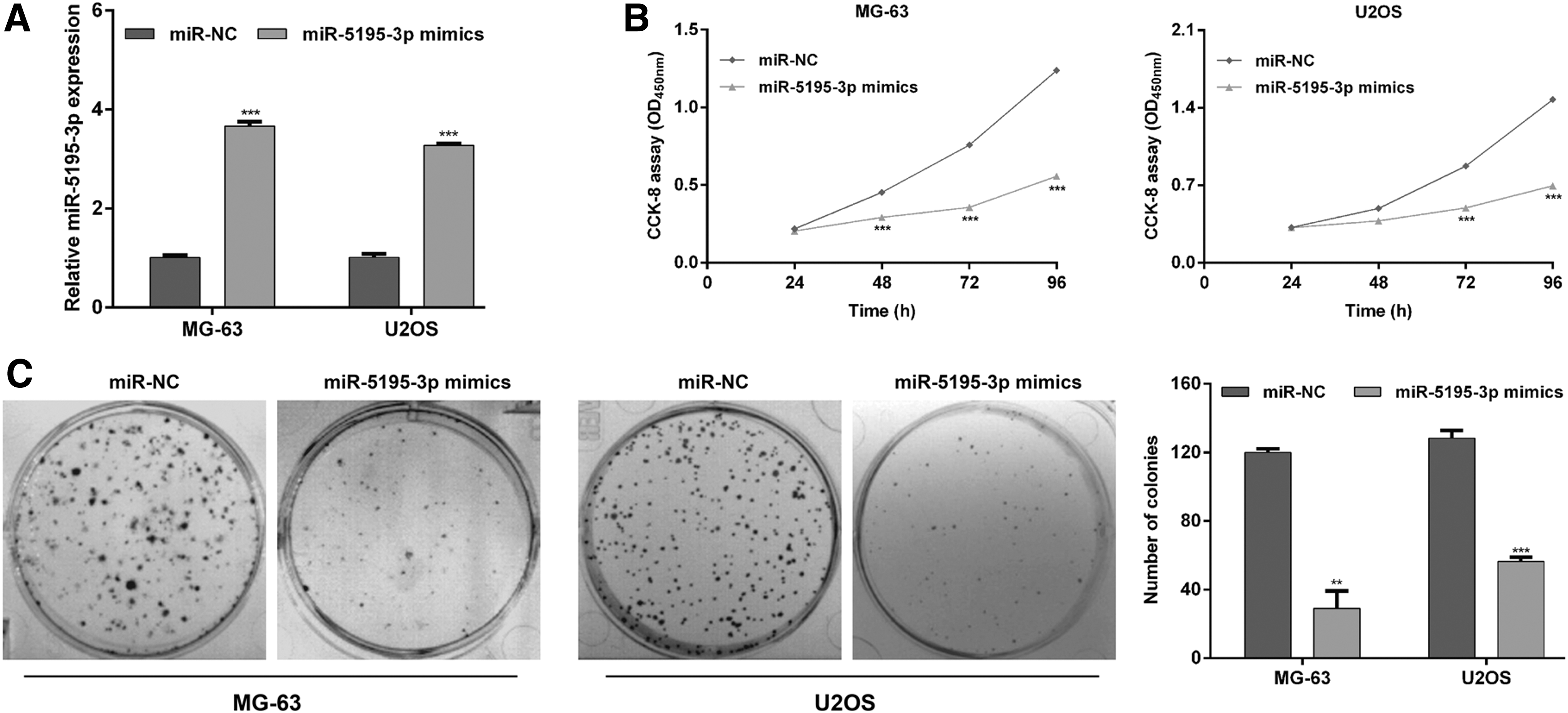
Effects of miR-5195-3p overexpression on OS cell proliferation. MG-63 and U2OS cells were transfected with miR-5195-3p mimics or miR-NC, respectively.
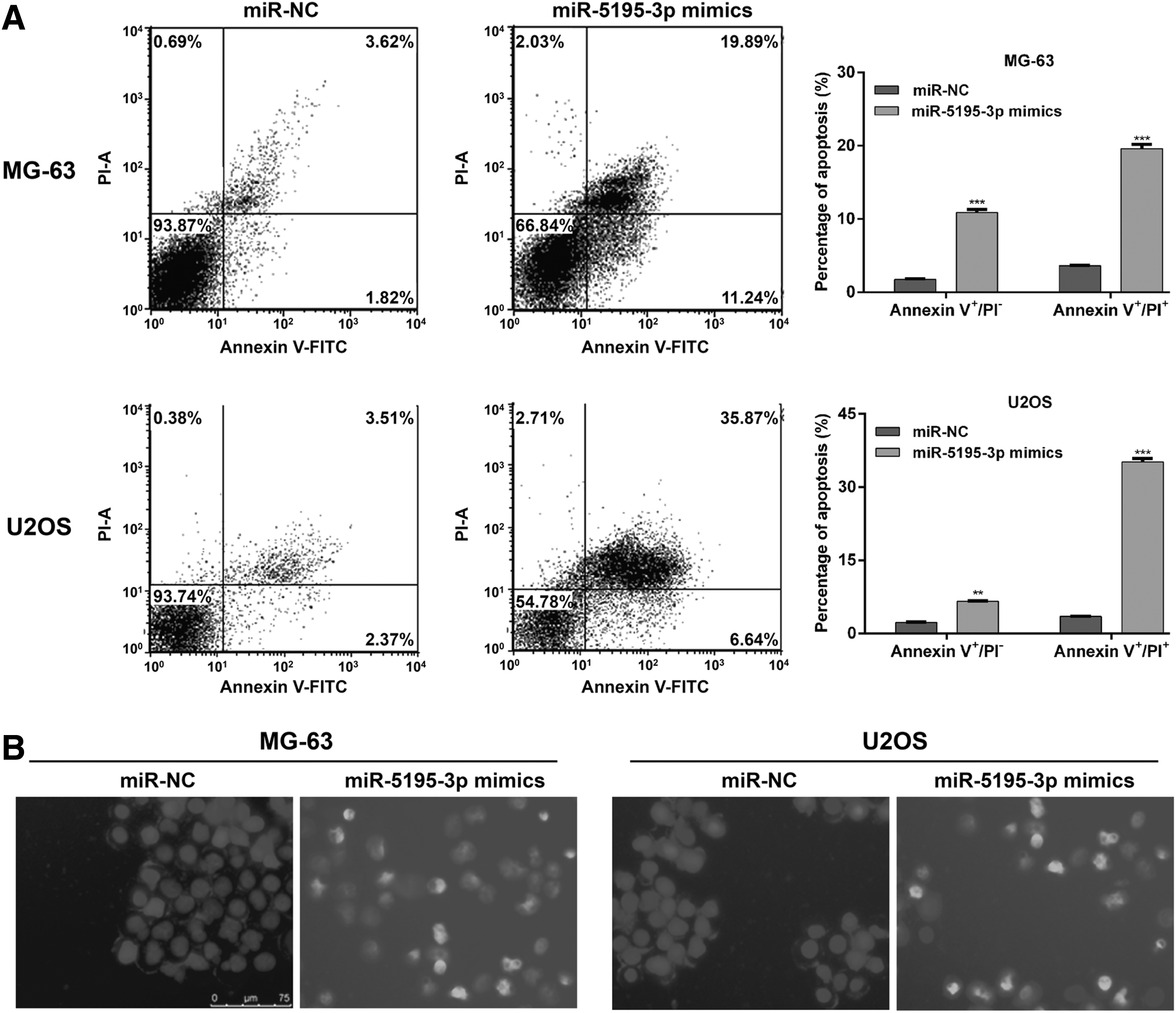
Effects of miR-5195-3p overexpression on OS cell apoptosis. MG-63 and U2OS cells were transfected with miR-5195-3p mimics or miR-NC, respectively.
NEDD9 was a direct target gene of miR-5195-3p in OS cells
Furthermore, the potential targets of miR-5195-3p were searched using online databases. Among the predicted target genes, NEDD9, has been implicated as an oncogene in several tumors, was selected as a promising target of miR-5195-3p. As shown in Figure 4A, the 3′UTR of NEDD9 contained the miR-5195-3p predicted target site. Then, luciferase reporter assays were performed to confirm whether miR-5195-3p targeted NEDD9 in OS cells. The authors can see from Figure 4B that in vitro transfection of the miR-5195-3p mimics lead to decreased luciferase activity of WT-3′UTR-NEDD9, but not in the MUT-3′UTR-NEDD9 (MG-63, p < 0.001; U2OS, p < 0.001). Furthermore, the expression of NEDD9 was examined following miR-5195-3p overexpression in OS cells. As illustrated in Figure 4D, elevated expression of miR-5195-3p suppressed NEDD9 mRNA expression in MG-63 and U2OS cells according to data from quantitative real-time PCR experiments (p < 0.001). Subsequent western blot analysis validated the results from quantitative real-time PCR analysis (Fig. 4E). In addition, the expression of NEDD9 was determined in samples from 26 pairs of OS tissues and the data showed that it was significantly elevated in OS tissues compared with matched normal tissues (Fig. 4F, p < 0.001). Further analysis revealed that the relative expression of NEDD9 was inversely correlated with the expression of miR-5195-3p in OS tissues (Fig. 4G, r = −0.4470, p = 0.0220). Collectively, these results demonstrated that NEDD9 might be a direct target of miR-5195-3p in OS.

Interaction between miR-5195-3p and NEDD9 in OS.
Reintroduction of NEDD9 rescued the impacts of miR-5195-3p on OS cells
As miR-5195-3p negatively regulated NEDD9 in OS cells, the authors further investigated whether miR-5195-3p targeting NEDD9 affects OS cell proliferation and apoptosis by performing rescue experiments. First, the expression of NEDD9 was restored by transfecting NEDD9 expression vector (pcDNA3.1-NEDD9) into the miR-5195-3p overexpressing-MG-63 cells. Obviously, quantitative real-time PCR (Fig. 5A, p < 0.001) and western blot (Fig. 5B) confirmed that the expression of NEDD9 was reduced after the above transfection. The result from CCK-8 assay demonstrated that NEDD9 overexpression partially alleviated the inhibition of miR-5195-3p amplification in MG-63 cells (Fig. 5C, p < 0.01, p < 0.001). Moreover, flow cytometry analysis showed that ectopic expression of NEDD9 decreased the miR-5195-3p overexpressing MG-63 cell early (p < 0.01) and late (p < 0.001) apoptosis (Fig. 5D). These data further indicated that NEDD9 might be a negative regulator in the suppressive effects of miR-5195-3p on OS cells proliferation.

Restoration of NEDD9 affected cell proliferation and apoptosis in miR-5195-3p overexpressing-MG-63 cells. NEDD9 expression vector (pcDNA3.1-NEDD9) or empty vector was transfected into the miR-5195-3p overexpressing-MG-63 cells.
Discussion
In this study, the authors show that miR-5195-3p was downregulated in OS tissues and cell lines. The in vitro experiments showed that upregulation of miR-5195-3p intensely inhibited cells growth and induced apoptosis in OS cells. Mechanically, they further found that NEDD9 might be a direct target gene of miR-5195-3p in OS cells.
Nowadays, noncoding RNAs expression profiles, DNA methylation, histone modifications, and nucleosome positioning are emerging as key players in cancer initiation and progression. 22 Until now, there has been little research on the biological role of miR-5195-3p in human cancers. Recently, Jiang et al. 13 have shown that miR-5195-3p attenuates proliferation and invasion behavior of bladder cancer cells by downregulating KLF5. The tumor suppressive role of miR-5195-3p was also evidenced by its target MYO6 network in nonsmall cell lung cancer. 12 Consistently, they also observed that reexpression of miR-5195-3p inhibited proliferation in MG-63 and U2OS cells. Simultaneously, their work also first identified miR-5195-3p as an inducer of apoptosis in OS cells. Bioinformatics prediction and luciferase reporter assay revealed that miR-5195-3p targeted the oncogene NEDD9 by binding to its 3′-UTR and downregulating its mRNA and protein levels.
In recent years, aberrant expression of the NEDD9 scaffolding protein has been established as a prometastatic marker in multiple cancer types. 16 Amplification of NEDD9 is often mentioned in human cancers. 23 For instance, serum NEDD9 levels are reported to be higher in pancreatic cancer compared with normal serum samples. 24 Downregulation of NEDD9 by miR-451 could increase chemosensitivity of prostate cancer cells. 25 In addition, NEDD9 is also shown to promote tumor progression, including ovarian cancer, 16 renal cell carcinoma, 26 and acute myeloid leukemia. 27 In this study, they observed that NEDD9 is overexpressed in OS tissues, and there is a negative correlation between NEDD9 and miR-5195-3p, implying a highly relevant cancer gene governing cell growth potential in OS. Temporally and spatially controlled activation of NEDD9 regulates multiple signaling pathways and controls diverse cellular functions in proliferation, fate determination, and depression of tumorigenesis. 28 NEDD9 has a crucial role in coordinating the FAK and Src signaling cascades, serves as the driving force of the integrin-mediated survival, migration, and adhesion. 15,23 From a previous literature, knockdown of NEDD9 is involved in inactivation of Akt and ERK signaling pathways to attenuate proliferation of pancreatic cancer cells. 28 It has been shown that decreased phosphorylation of mammalian target of rapamycin are correlated with inhibition of the upstream Akt signaling and contribute to the cell cycle block and apoptosis. 29 While, ERK activation by MEK plays a key role in cell growth and apoptosis. 30 The emergence of NEDD9 in the epithelial–mesenchymal transition and invasion of prostate cancer cells has been reported to be associated with transforming growth factor-β (TGF-β). 31 Particularly, TGF-β also acts in the promotion of cancer cells proliferation and survival. 32 Except for implicate in several signaling cascades, NEDD9 also functions as a scaffold for mitotic kinase Aurora-A and MMP14. 18,33 Previous studies showed that NEDD9 depletion leads to AURKA destabilization and MMP14 redistribution in mice harboring xenografts of breast tumors, and leading to impairment in tumor growth and metastasis. 18,33 The proposed molecular mechanisms about the role of NEDD9 in cell proliferation and survival are the focus of future investigations.
In summary, downregulation of miR-5195-3p was found in OS tissues and cell lines, and enforced expression of miR-5195-3p inhibited the OS cell proliferation and induced apoptosis through targeting the expression of NEDD9. The findings of the authors may provide an experimental validation and theoretical foundation for developing precise cancer therapy.
Availability of Data and Materials
The data and materials are available under the permission of the authors.
Ethics Approval and Consent to Participate
All tissue samples were collected with written informed consent in accordance to the Declaration of Helsinki. This study was approved by the Research Ethics Committee of the Shanghai Electric Power Hospital (Shanghai, China).
Footnotes
Disclosure Statement
No competing financial interests exist.