
Abstract
Background:
MicroRNA-589-5p (miR-589-5p) has been recently reported to be aberrantly regulated in hepatocellular carcinoma, but its functional role and molecular mechanisms still remains unknown in the endometrial carcinoma (EC) as one of the most common female malignancies.
Methods:
EC tissues and adjacent tissues were collected to determine the expression of miR-589-5p and thyroid receptor interacting protein 6 (TRIP6) using quantitative real time-PCR. Subsequently, two EC cell lines HEC-1B and AN3CA were transfected with miR-589-5p to achieve miR-589-5p overexpression. Using Cell Counting Kit-8 (CCK-8), a wound healing assay and the Transwell assay, we analyzed cell proliferation, migration and invasion. Dual-luciferase reporter assay confirmed that thyroid receptor interacting protein 6 (TRIP6) was a direct target of miR-589-5p.
Results:
We first observed that miR-589-5p was down-regulated in EC tissues compared with normal endometrial tissues. MiR-589-5p overexpression significantly suppressed EC cell proliferation, migration and invasion. Thyroid receptor interacting protein 6 (TRIP6) was a direct target of miR-589-5p. Besides, TRIP6 knockdown presented similar effects on cell proliferation, migration and invasion to miR-589-5p overexpression. Furthermore, TRIP6 knockdown efficiently enhanced the effects of miR-589-5p on the above cellular function. Moreover, miR-589-5p up-regulated E-cadherin expression, but down-regulated N-cadherin and Vimentin by targeting TRIP6.
Conclusions:
In summary, miR-589-5p might function as a tumor suppressor by targeting TRIP6, which will provide new insights into the molecular mechanism underlying the development of EC.
Introduction
Endometrial carcinoma (EC) remains the most common malignancy of the female genital tract in developing and developed countries. 1,2 EC is a disease of pre-, peri-, and postmenopausal women, and rarely occurs in young women: the proportion of patients, ages 40 years and younger, was from 2% to 14%. 3 Based on clinical and endocrine features, EC can be classified into the major subtypes, type I and type II carcinoma, which accounts for 80% and 20% of all EC cases, respectively. 4,5 Despite mortality produced by EC being relatively low compared with other gynecological malignancies, it represents 34.6% of cancer-related deaths in the female reproductive system in the United States. 6 An emerging body of evidence suggests that many oncogenes, antioncogenes, and mutant genes are implicated in the progress of EC pathology. A deep understanding of the pathological mechanism of EC may have a significant impact on future modalities and initiate the development of a novel therapeutic strategy for this disease.
MicroRNAs (miRNAs) are an abundant class of ∼22-nucleotide-long, noncoding RNAs that powerfully regulate the expression of target messenger RNAs (mRNAs), which harbor partially complementary sequences. 7 During the past decade, aberrant expression of miRNAs in malignant cells is known to play a key role in the multistep process of carcinogenesis, from initiation and development to progression toward the acquisition of metastatic features. 8 The earliest observation that supplied a potential link between miRNA and carcinoma biology was the regulation of developmental timing by let-7 in Caenorhabditis elegans. 9 From then on, a large number of miRNAs that are dysregulated in tumors were uncovered by many studies. 10 Those miRNAs expressed at high levels in cancers and which facilitate tumorigenesis are known as “oncomiRs,” while the low-expressed miRNAs that function to inhibit tumor growth are called “anti-oncomiRs.” 11 MicroRNA-589-5p (miR-589-5p), which is a candidate prognostic biomarker of hepatocellular carcinoma (HCC), as it is overexpressed in HepG2 and HuH-7 cells, was sufficient to promote cancer cell proliferation, migration, and invasion. 12 –14 Hypermethylation of the miR-589 gene promoter caused downregulation of miR-589-5p expression in nonsmall-cell lung cancer (NSCLC) cells. 15 Despite miR-589-5p being reported in HCC and NSCLC, the biological behavior of miR-589-5p in EC remains to be elucidated.
Thyroid receptor interacting protein 6 (TRIP6), also called zyxin-related protein 1, has 476-amino acid residues, and belongs to a subfamily of LIM domain protein. 16,17 TRIP6 is located on chromosome 7q22 and mainly localized to the cytoplasm or focal adhesion plaques. 18 It is capable of nucleocytoplasmic shuttling and has been suggested to serve as a transcriptional coregulator. 19 Previous studies have shown that TRIP6 could interact with different proteins to act as a platform for numerous signaling molecules' recruitment, associated with multiple processes such as proliferation, differentiation, apoptosis, invasion, and actin cytoskeleton rearrangement. 20 –22 Recently, the positive role of TRIP6 has been defined in several types of cancer. A proproliferative function of TRIP6 was reported in non-Hodgkin's lymphoma (NHL), 23 HCC, 20 and ovarian cancer. 23 In Ewing's sarcoma, depletion of TRIP6 resulted in decreased migration, invasion, and cell growth. 24 Prior cross talk analysis revealed a connection between miR-138-5p and TRIP6 in regulating neural stem cell proliferation and differentiation. 21 However, the biological role of TRIP6 and its relationship with miR-589-5p in EC remain unexplored.
The objective of this study is to decipher the action of miR-589-5p and TRIP6 in EC and identify the correlation between these two genes. To address this, the authors evaluated the expression of both genes in EC tissues and matched adjacent endometrial tissues. Pearson's correlation coefficient analysis was performed to determine the correlation between miR-589-5p and TRIP6. Then, they used a “gain-of-function” approach to assess the contribution of miR-589-5p in EC proliferation, invasion, and migration. The results from bioinformatic algorithms, luciferase reporter assay, and intensified experiment showed that miR-589-5p could act as tumor suppressor in EC tumorigenesis through direct targeting of TRIP6. The role of miR-589-5p/TRIP6 axis in EC progression will no doubt enrich the knowledge of EC pathology.
Materials and Methods
Patient samples
A total of 40 pairs of tumor specimens and matched adjacent nontumor endometrial tissues were collected from EC patients (mean age: 45 years, range: 38–64 years) after signing a written informed consent in the People's Hospital of Dezhou (Shandong, China). Tissues were snap-frozen immediately after surgery and preserved at −80°C until RNA extraction. Some basic patient characteristics are summarized in Table 1. According to the International Federation of Gynecology and Obstetrics classification system 25 and World Health Organization criteria, tumor stage, histological subtypes, and grading were determined, respectively. This study was performed according to the principles of the Declaration of Helsinki and obtained the approval from the Ethics Committee of People's Hospital of Dezhou.
Patient Characteristics (n = 40)
FIGO, International Federation of Gynecology and Obstetrics.
Cell culture and transfection
The EC cell lines, HEC-1B and AN3CA, were purchased from the American Tissue Culture Collection (ATCC, Manassas, VA) and cultured in Eagle's Minimum Essential Medium (Invitrogen, Carlsbad, CA) with 10% fetal bovine serum (FBS; Gibco, Grand Island, NY) in a humidified atmosphere of 5% carbon dioxide at 37°C.
The plasmids encoding miR-589-5p mimic and TRIP6 small interfering RNA (siTRIP6), as well as their corresponding negative control vector (miR-NC and siNC, respectively), were purchased from GenePharma (Shanghai, China). The HEC-1B or AN3CA cells were seeded into a 12-well plate at a density of 1.5 × 105 per well and transfected with the above plasmids by Lipofectamine 2000 (Invitrogen) according to the manufacturer's protocol. After a 24 or 48 h transfection, the transfected cells were used for the following experiments.
Quantitative real-time PCR
The total RNA was extracted by the TRIzol reagent (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer's protocol. After synthesizing complementary DNA using the TIAN Script RT Kit (Tiangen Biotech, Beijing, China), quantitative real-time PCR (qRT-PCR) was performed on ABI 7300 system (Applied Biosystems, Foster City, CA) with TaqMan Human MiRNA Assay (GeneCopoeia, Guangzhou, China) for miR-589-5p and SYBR Green Premix (Takara, Dalian, China) for TRIP6 using U6 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as internal control, respectively.
Cell proliferation assay
The EC cell proliferation capacity was determined using the Cell Counting Kit-8 (CCK-8; Dojindo, Japan). Approximately 4000 cells were seeded per well in a 96-well plate and cultured overnight. At the indicated time points (24, 48, and 72 h, respectively), 10 μL CCK-8 reagent was added per well and incubated for 2 h at 37°C. The absorbance of each well was measured with microplate reader at 450 nm and the growth curve was drawn according to the absorbance.
Wound healing assay
EC cell migration ability was accessed by the wound healing assay. In brief, cells were grown in six-well plates until ∼90% confluency and wounds were artificially created with a 100-μL pipette tip through the monolayer. At 0 and 24 h after wounding, the images of cells migrating to the site of wound were observed using microscopy (Olympus, Tokyo, Japan). By measured the widths of the wounded areas at 0 (W0) and 24 h (W24) in five randomly selected areas, the authors calculated the relative average migration distance with the following formula: (W0–W24)/W0 × 100%.
Transwell invasion assay
EC cell migration ability was evaluated using Transwell plates with 8 μM pore filters (Corning, NY) precoated with Matrigel (BD Biosciences, Franklin Lakes, NJ). Briefly, cells were suspended in 200 μL of serum-free medium and at a density of 5 × 104 per well reseeded in the upper chambers. Subsequently, 500 μL of culture medium containing 10% FBS as the chemoattractant was added into the lower chambers. After a 24-h incubation, the cells that migrated to the lower chamber were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. The average number of invasive cells was calculated in five random fields under an inverted fluorescence microscope (Nikon, Japan).
Target gene predication and dual-luciferase reporter assay
First, the target genes of miR-589-5p were predicted using TargetScan (
Western blot analysis
Total protein was extracted with RIPA lysis buffer (Beyotime Biotechnology, Shanghai, China) and quantified using the BCA Protein Assay Kit (Beyotime Biotechnology). Next, equal amounts of protein were separated by 10% sodium dodecyl sulfate/polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Burlington, MA). After being probed with primary antibodies against TRIP6, E-cadherin, N-cadherin, Vimentin, or GAPDH overnight at 4°C, the membranes were subsequently incubated with horseradish peroxidase-conjugated secondary antibodies for 2 h at room temperature. Finally, the immunoreactive bands were visualized by enhanced chemiluminescence (Pierce, Cambridge, MA). GAPDH was used as a loading control.
Statistical analyses
All statistical analyses were performed with SPSS version 17.0 (SPSS, Inc., Chicago, IL). Data are expressed as the mean ± standard deviation of at least three independent experiments. Student's t-test was used for comparisons between 2 independent groups, and one-way analysis of variance was used to compare differences among multiple groups followed by Dunnett's post hoc test. Pearson's correlation coefficient analyses were used for the determination of correlation. The values of p < 0.05 were considered to be statistically significant.
Results
MiR-589-5p was downregulated and negatively correlated with the expression of TRIP6 in EC tissues
A total of 40 pairs of EC tissues and matched adjacent endometrial tissues were detected for miR-589-5p and TRIP6 expression using qRT-PCR. As shown in Figure 1A, the expression of miR-589-5p was significantly downregulated in EC tissues compared with adjacent tissues (p < 0.001). On the contrary, the authors found that the mRNA levels of TRIP6 in EC tissues were significantly higher than those in adjacent endometrial tissues (Fig. 1B, p < 0.001). Using Pearson's correlation coefficient analyses, they found that the expression of miR-589-5p was inversely correlated with the TRIP6 expression in EC tissues (Fig. 1C, p = 0.0189). These data indicated that altered miR-589-5p and TRIP6 expression levels might play an important role in EC progression and initiation.
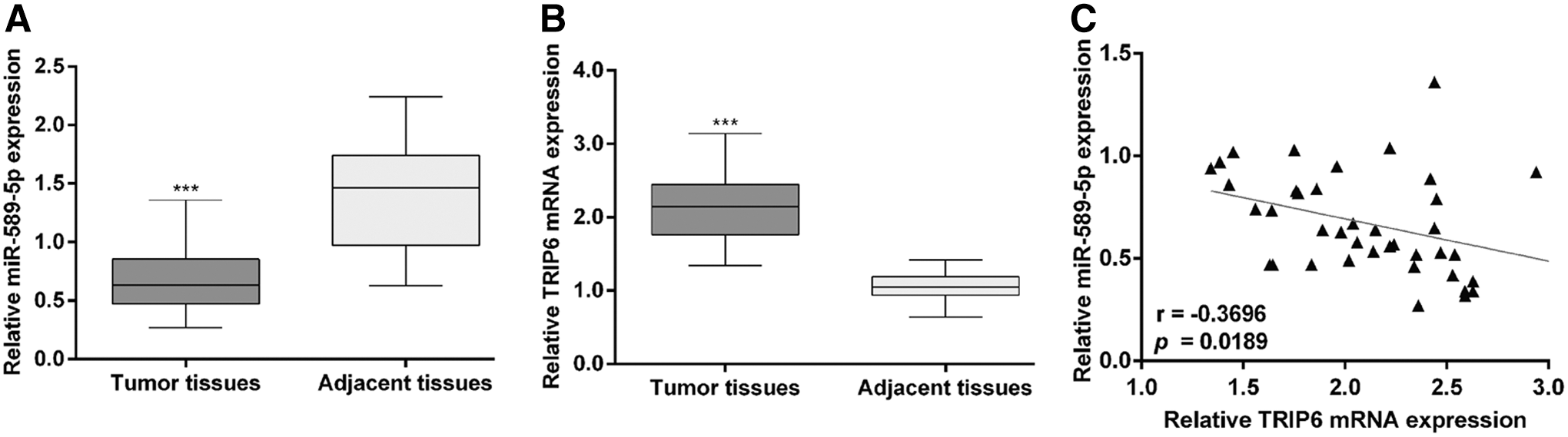
The expression of miR-589-5p and TRIP6 in EC tissues. Forty pairs of EC tissues and matched adjacent endometrial tissues were detected by qRT-PCR for
Overexpression of miR-589-5p dramatically suppressed EC cell proliferation, migration, and invasion
To better understand the biological function of miR-589-5p in EC in vitro, two EC cell lines, HEC-1B and AN3CA, were transfected with miR-589-5p mimic or miR-NC as a negative control. The successful overexpression of miR-589-5p was first validated by qRT-PCR (Fig. 2A) in HEC-1B (p < 0.001) and AN3CA (p < 0.001) cells. The CCK-8 assay showed that overexpression of miR-589-5p significantly suppressed cell proliferation in HEC-1B and AN3CA cells (Fig. 2B, p < 0.001). In addition, the effects of miR-589-5p on cell migration and invasion were also determined. As shown in Figure 2C, the wound healing assay showed that HEC-1B and AN3CA cells transfected with miR-589-5p mimic presented slower closing of the scratch wound compared with the control cells (p < 0.01, p < 0.001). The transwell assays (Fig. 2D) showed that the number of invasive cells was significantly decreased in the miR-589-5p mimic group compared with miR-NC group in HEC-1B (41.7 ± 6.5 vs 107.0 ± 4.6, p < 0.001) and AN3CA (69.0 ± 6.2 vs 161.0 ± 10.4, p < 0.001) cells. Collectively, these results suggested that overexpression of miR-589-5p played a negative role in EC cellular function.

Effects of miR-589-5p overexpression on EC cell proliferation, migration, and invasion. HEC-1B and AN3CA cells were, respectively, transfected with miR-589-5p mimic or miR-NC.
TRIP6 was a direct target of miR-589-5p in EC cells
The qRT-PCR analysis indicated that miR-589-5p was inversely correlated with the TRIP6 expression in EC tissues, implying TRIP6 may be a target gene of miR-589-5p. To confirm this hypothesis, the authors used online TargetScan and MiRanda databases to predict the target genes of miR-589-5p. As expected, they found that the complementary sequence of miR-589-5p was identified in the 3′-UTR of TRIP6 mRNA (Fig. 3A). Moreover, luciferase activity of 3′-UTR of TRIP6 was significantly reduced after cotransfection with miR-589-5p mimic and WT-TRIP6, compared with miR-NC transfection, but did not obviously change after cotransfection with miR-589-5p mimic and MUT-TRIP6 in HEC-1B (Fig. 3B) and AN3CA (Fig. 3C) cells. In addition, the qRT-PCR and Western blotting results showed that miR-589-5p mimic markedly decreased TRIP6 mRNA (Fig. 3D, p < 0.01) and protein (Fig. 3E) expression levels compared with miR-NC groups in HEC-1B and AN3CA cells.
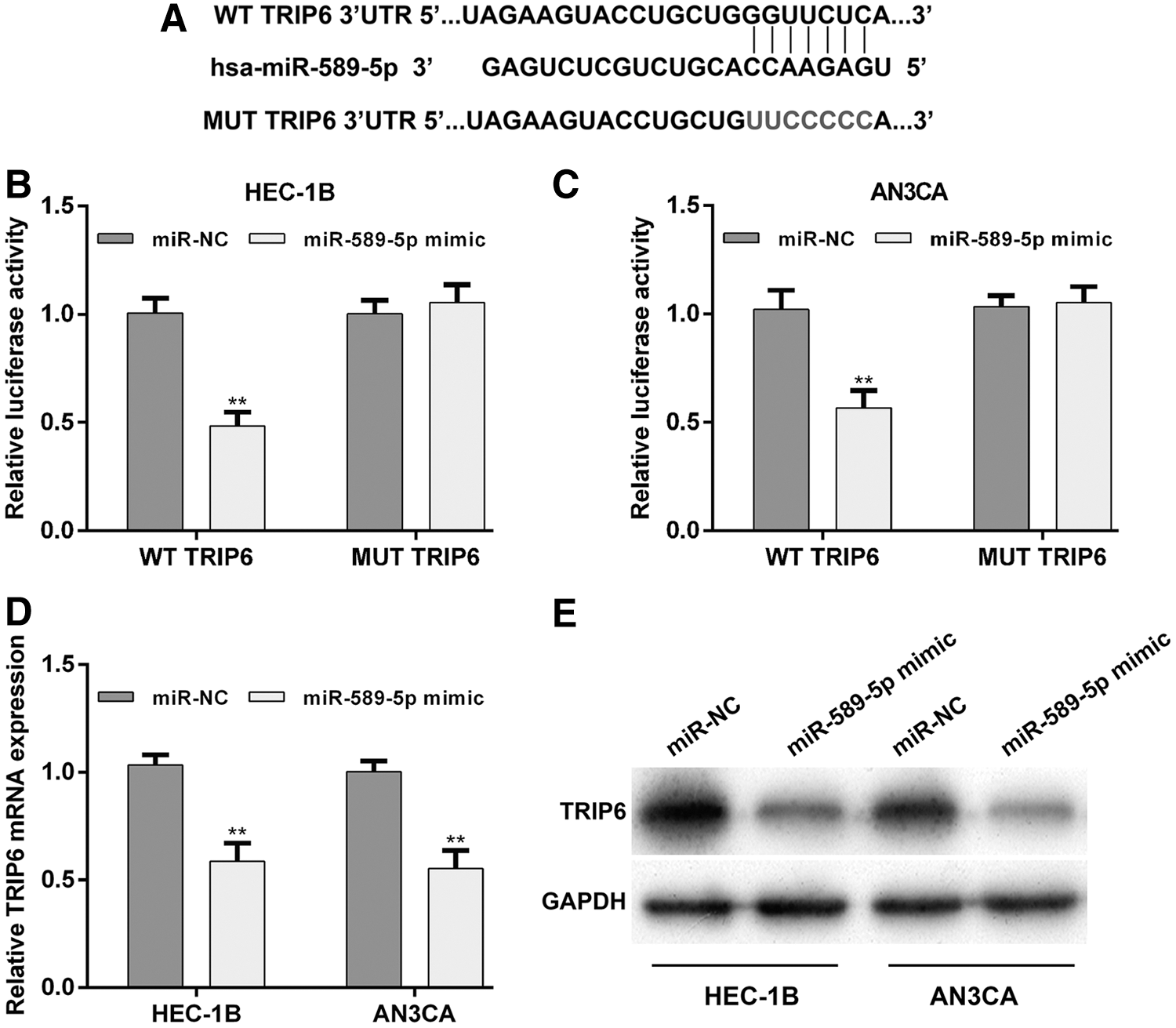
TRIP6 was a direct target of miR-589-5p.
Downregulation of TRIP6 enhanced the inhibitory effects of miR-589-5p on EC cell proliferation, migration, and invasion
Considering TRIP6 was upregulated in EC tissues, the authors speculated that it might be an oncogene involved in the progression of EC. Thus, they performed loss-of-function assays on HEC-1B cells with or without miR-589-5p mimic transfection by transfecting with siTRIP6 or siNC. As shown in Figure 4A and B, the mRNA and protein expression levels of TRIP6 were obviously downregulated after siTRIP6 transfection compared with siNC transfection, remarkably reduced after cotransfection with miR-589-5p mimic and siTRIP6, in comparison with sole miR-589-5p mimic transfection. Results from CCK-8 assay (Fig. 4C) showed that sole TRIP6 knockdown significantly inhibited cell proliferation (p < 0.001), and a combination of TRIP6 knockdown and miR-589-5p overexpression caused the lowest cell growth rate in HEC-1B cells (p < 0.01). In addition, TRIP6 knockdown not only impaired cell migration and invasion ability but also enhanced the suppressive effects of miR-589-5p overexpression on cell migration and invasion, as determined by wound healing assay (Fig. 4D) and transwell assay (Fig. 4E), respectively. These data demonstrated that TRIP6 might be a direct functional downstream target of miR-589-5p in EC cells.
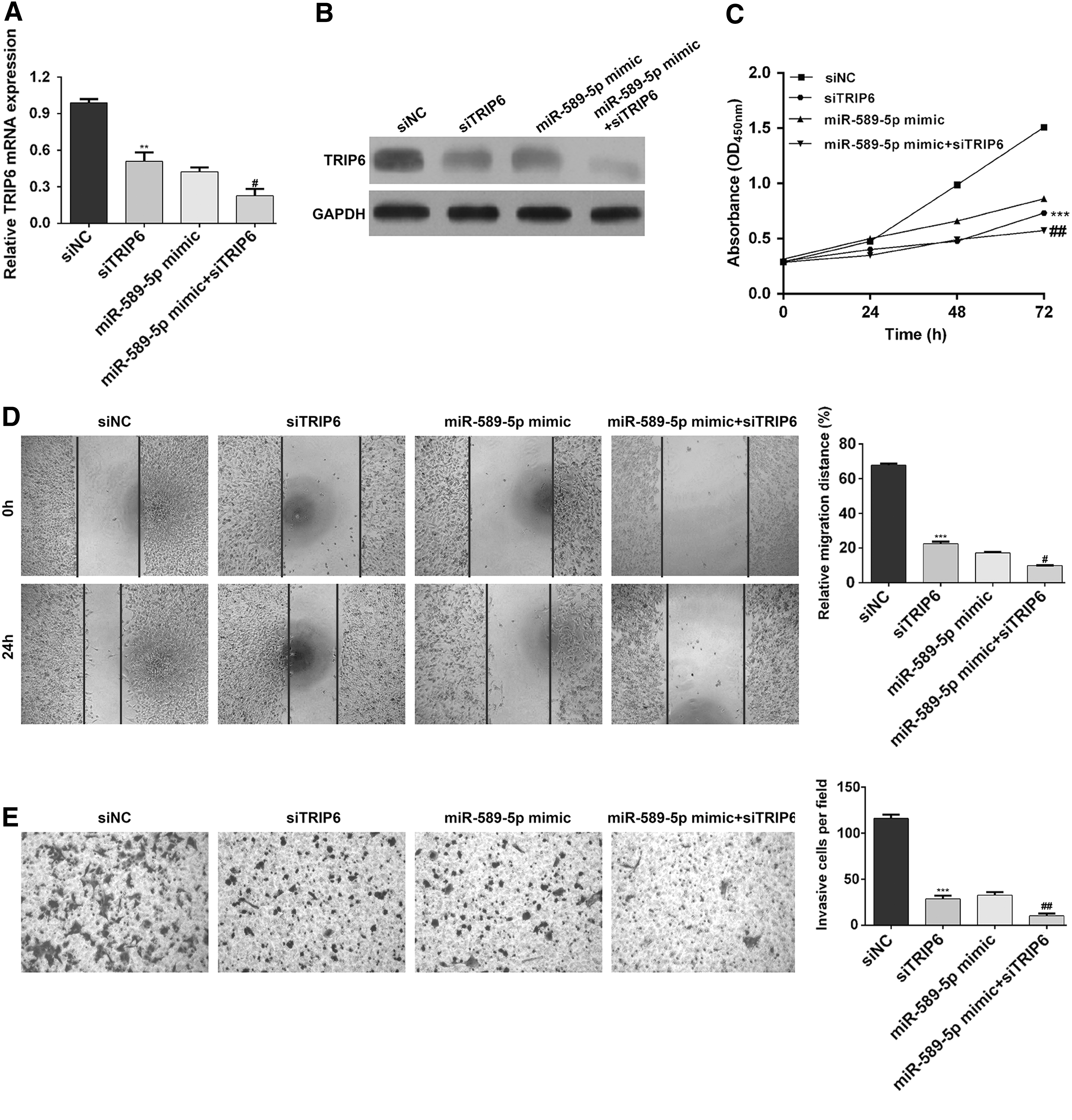
Downregulation of TRIP6 enhanced the inhibitory effects of miR-589-5p on EC cell proliferation, migration, and invasion. HEC-1B cells were transfected with siTRIP6, siNC, miR-589-5p mimic, or miR-589-5p mimic + siTRIP6, respectively. The mRNA and protein expression levels of TRIP6 were determined in HEC-1B cells using qRT-PCR
MiR-589-5p upregulated E-cadherin expression by targeting TRIP6
The authors further investigated the molecular mechanisms involved in miR-589-5p-mediated regulation of cell migration and invasion in EC cells using Western blotting. As shown in Figure 5, TRIP6 knockdown notably increased E-cadherin expression, but decreased the expression of N-cadherin and Vimentin. Interestingly, the elevated E-cadherin and reduced N-cadherin and Vimentin expression by miR-589-5p overexpression was obviously strengthened by TRIP6 knockdown. Therefore, they concluded that the inhibited cell migration and invasion by miR-589-5p might be partially via downregulating TRIP6 in EC cells.
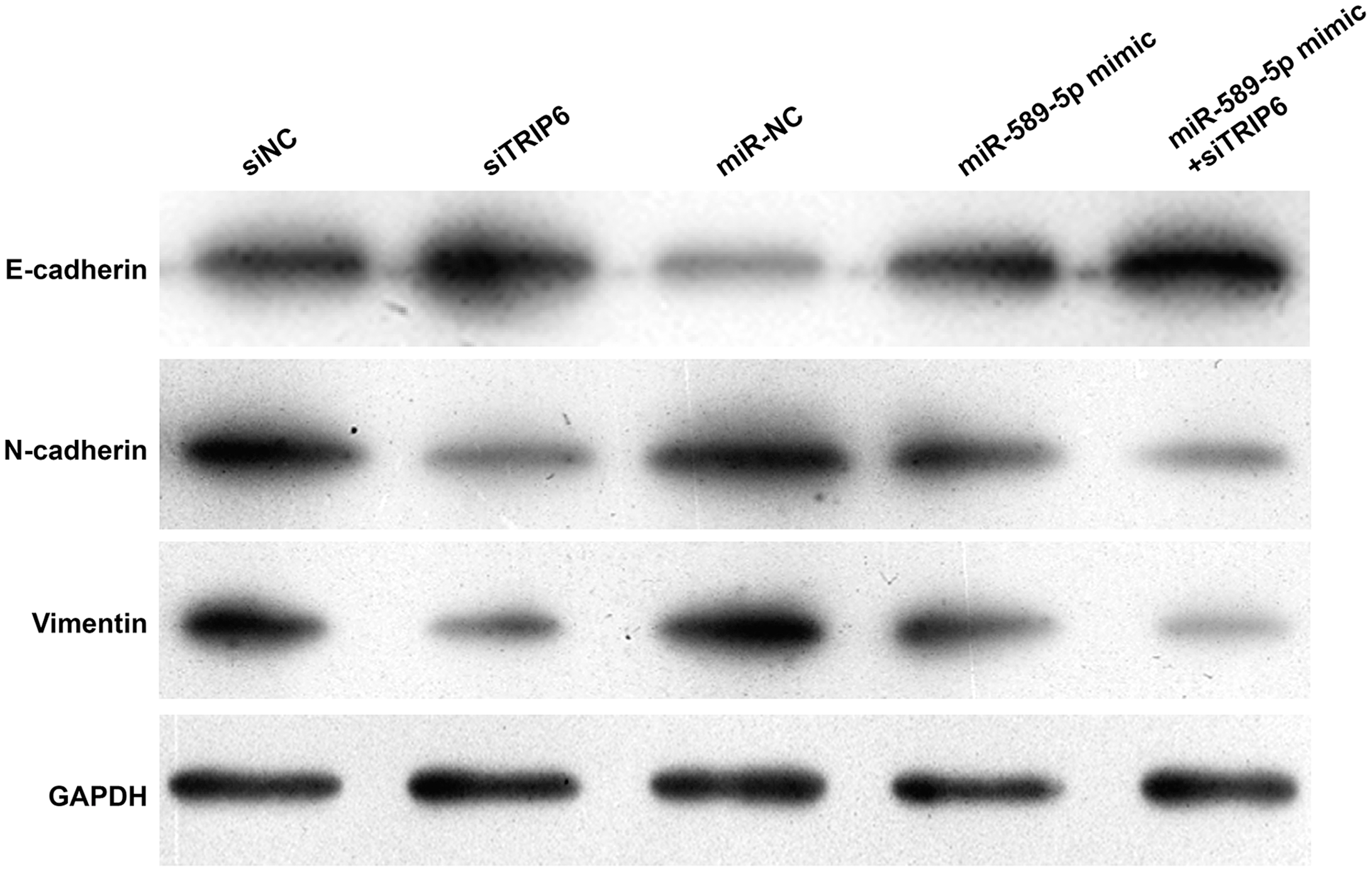
MiR-589-5p transfection regulated the expression of E-cadherin, N-cadherin, and Vimentin by targeting TRIP6. HEC-1B cells were transfected with siTRIP6, siNC, miR-NC, miR-589-5p mimic, or miR-589-5p mimic + siTRIP6, respectively. After transfection, Western blotting assay was used to determine the protein expression of E-cadherin, N-cadherin, and Vimentin.
Discussion
In the present study, the authors report, for the first time, that the miRNA miR-589-5p is downregulated and the LIM domain protein TRIP6 is upregulated in EC tissues compared with their matched adjacent endometrial tissues. MiR-589-5p expression was inversely correlated with TRIP6 expression. Furthermore, they uncover a functional connection between miR-589-5p and TRIP6 in the malignant context. They observed that TRIP6 is a direct target of miR-589-5p, and overexpression of miR-589-5p by mimics is sufficient to induce the downregulation of endogenous levels of TRIP6. Cotransfection of cells with miR-589-5p mimic and siTRIP6 enhanced the inhibitory effects of miR-589-5p on cell proliferation, migration, and invasion, indicating that increased expression of miR-589-5p enhanced the malignant phenotype of EC partially through targeting TRIP6.
Uncontrolled proliferation is considered a primary feature of cancer, and malignant cells have typically acquired damage to genes that directly or indirectly modulate their cell cycle progression. 26 In this study, miR-589-5p mimic transfection resulted in a defect in TRIP6 expression. Based on previous findings, the pleiotropic nature of TRIP6 indicates that gene expression profiles and signaling pathways can be influenced by the abnormal expression of TRIP6, and therefore obviously guiding cancer cell proliferation. For instance, enhanced expression of TRIP6 has been shown to facilitate proliferation in NHL through increased expression of a cell cycle inhibitor p27Kip1 by decreasing phosphorylation of p27Kip1 at T157. 23 Zhao et al. 20 reported that TRIP6 can stimulate HCC cell proliferation via attenuation of FOXO3a. Activation of Yes-associated protein, which acts as a tumor promoter, could, in part, be caused by TRIP6 through suppressing the LATS1/2 kinases. 16 Structural and functional analyses have revealed that TRIP6 has multiple transactivation domains that can trigger transcription. 27 Although the precise mechanisms of TRIP6 deficiency involved in miR-589-5p-mediated inhibition of proliferation remain to be elucidated, the authors will focus on the above factors in future work.
By gain-of-function and loss-of-function studies, the authors show that knockdown of TRIP6 is an important reason for miR-589-5p-mediated inhibition of EC cell migration and invasion. Consistent with the reduced migratory ability and less invasive phenotype of TRIP6 knockdown and miRNA-589-5p mimic, E-cadherin was upregulated, but N-cadherin and Vimentin were downregulated. E-cadherin is a cell–cell adhesion molecule associated with epithelial to mesenchymal transition. 28 Downregulation of E-cadherin is considered a critical molecular feature of invasiveness and aggressiveness in multiple malignancies, including EC. 29 In fact, decreased invasive and migratory capabilities of the miR-589-5p mimic EC cells are associated with the reduction of E-cadherin expression and increase of N-cadherin and Vimentin. Protein–protein interactions mediated by multidomain have underscored the role of TRIP6 as a platform for the recruitment of numerous signaling molecules that are associated with migration and invasion. 19 Some such examples include the following: (1) TRIP6 could contribute to the lysophosphatidic acid 2 (LPA2)-induced cell migration due to interaction between the LIM domains of TRIP6 and the carboxyl-terminal tail of the LPA2 receptor 30 ; (2) TRIP6 also interacts with RIP2, a protein that is crucially implicated in nuclear factor-κB and ERK activation, both of which are inducers of cancer cell migration and proliferation 31 ; (3) TRIP6 is sufficient to promote invasiveness by binding directly to the fifth PDZ domain of the MAGI-1b. 32 In this study, there may exist a physical and functional association between TRIP6 and some molecules that are involved in migration and invasion, thereby leading to a significant upregulation of E-cadherin and downregulation of N-cadherin and Vimentin.
In summary, the authors obtained evidence that miR-589-5p can act as an “anti-oncomiR” repressing the expression of the tumorigenesis inducer TRIP6. Although more studies are necessary to thoroughly determine the downstream genes and/or signaling pathways of the miR-589-5p/TRIP6 axis, these discoveries may open the way for a series of future studies that the miR-589-5p/TRIP6 axis may be useful in exploration of EC prevention strategies.
Footnotes
Funding Information
The study was supported by The People's Hospital of Dezhou.
Disclosure Statement
The authors declare that they have no competing interests.