
Abstract
Background:
Pyrroline-5-carboxylate reductase 1 (PYCR1) is involved in tumor progression, for instance, breast cancer and prostate cancer. However, its role in tumor metastasis, especially in nonsmall cell lung cancer (NSCLC), is still elusive.
Materials and Methods:
The messenger RNA (mRNA) expression of PYCR1 between NSCLC and normal lung specimens was compared using Oncomine database. The endogenous PYCR1 expressions in NSCLC cell lines 95C and H1299 were knocked down by lentiviral-mediated delivery of short hairpin RNA (shRNA). Then the effects of PYCR1 on the migration and invasion of NSCLC cells were studied by wound healing assay and transwell assay.
Results:
PYCR1 mRNA expression was significantly higher in NSCLC specimens than that in normal lung tissues. Depletion of PYCR1 in NSCLC cell significantly repressed the cell migration and invasion. Moreover, depletion of PYCR1 influenced the expression of epithelial-mesenchymal transition molecules E-cadherin, Vimentin, N-cadherin, and Snail1.
Conclusions:
Our data suggested that PYCR1 plays a positive role in NSCLC metastasis in vitro and might be a promising target for treating NSCLC.
Introduction
Lung cancer is known to be a major killer of cancer-related deaths worldwide. 1 According to the latest estimates of the American Cancer Society, the proportions of new cases and deaths of lung cancer in the United States raise up to 13.5% and 25.3%, respectively. 2 Also, lung cancer has been on the top of the malignant tumor morbidity and mortality for 10 consecutive years documented in China. 3 Currently, it is well established that the main causative factors of lung cancer are smoking, family history, genetic susceptibility, chronic obstructive pulmonary disease, and environmental exposures such as air pollution, asbestos, radon, and ionizing radiation. 4 –6 According to the clinic pathological features, lung cancer is classified into small cell lung cancer and nonsmall cell lung cancer (NSCLC), of which NSCLC accounts for about 85%. 7,8 NSCLC comprises lung adenocarcinoma, squamous-cell lung carcinoma, and large cell lung carcinoma. 8,9 Standard therapeutic paradigms for NSCLC include surgery, chemotherapy, radiotherapy, immunotherapy, and targeted therapy. 10 –13 Despite the constant improvement in therapeutic modalities, the therapeutic results of patients with advanced NSCLC are poor, with the median survival duration <1 year. 14 The recent National Comprehensive Cancer Network (NCCN) guidelines pointed out that advanced NSCLC patients with specific predictive biomarkers undergoing targeted therapy had a better life quality and progression-free survival than those undergoing chemotherapy. 15 Thus, it is important to search for potential predictive biomarkers.
Human pyrroline-5-carboxylate reductase 1 (PYCR1) is localized in the mitochondria, and catalyzes the final step in the NAD(P)H-dependent conversion of pyrroline-5-carboxylate (P5C) to proline (Pro). 16,17 It was recently reported that PYCR1 was activated in prostate cancer, breast cancer, colorectal carcinoma, and esophageal squamous cell cancer in recent years. 18 –21 In addition, PYCR1 expression was remarkably correlated with tumor size, grade, invasive molecular subtypes, and survival of mammary cancer patients, and it was also found that depletion of PYCR1 suppressed the competence of mammary cancer cell growth and invasion in vitro. 22 A recent study indicated PYCR1 enhanced cell proliferation and suppressed cell apoptosis in NSCLC. 23
Taking the previous findings together, we hypothesized that PYCR1 might associate with the metastasis of NSCLC cells and we found that the migration and invasion of 95C and H1299 cells were suppressed by knocking PYCR1 down using short hairpin RNA (shRNA). In other words, PYCR1 promoted cell migration and invasion in vitro.
Materials and Methods
Cell culture
Human NSCLC cell lines 95C and H1299, purchased from Cell Bank of Chinese Academy of Science (Shanghai, China), were incubated in RPMI-1640 medium (Hyclone, Logan, UT) with 10% fetal bovine serum (FBS; Lonsera, Shanghai, China) at 37°C in a humidified atmosphere with 5% CO2.
Expression of PYCR1 in NSCLC and normal lung specimens
Oncomine database (
Lentivirus-mediated shRNA targeting PYCR1 gene
The shRNA sequences, including shPYCR1-S1 (CACAGTTTCTGCTCTCAGGAA), shPYCR1-S2 (CCCTTCATCCTGGATGAAATA), and nontarget Control (shCon, TTCTCCGAACGTGTCACGT), were designed through MISSION Predesigned small interfering RNA (siRNA) website of the Sigma-Aldrich, and were accurately cloned into pGreenPuro™ vector (SBI). Recombinant vectors (pGreenPuro vector-S1, pGreenPuro vector-S2, and pGreenPuro vector-shCon) were transfected individually into HEK-293T cells for lentivirus packaging together with pHelper plasmids (Sigma, St. Louis, MO) using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) following the manufacturer's instructions.
NSCLC cells 95C and H1299 (1 × 105 cell/well) were infected individually using lentivirus with shPYCR1-S1, shPYCR1-S2, or shCon by coculturing cells and lentivirus (multiplicity of infection, 10) for 6 h. Twenty-four hours later, the expression of green fluorescence protein (GFP) was detected by a fluorescence microscope (Olympus, Tokyo, Japan).
Quantitative reverse transcription-polymerase chain reaction
Total RNA was extracted according to Trizol protocol (Invitrogen). Complementary DNA (cDNA) was synthesized using M-MLV Reverse Transcriptase (Promega, Madison, WI). SYBR premixex taq (TAKARA, Dalian, China) was used to perform the amplification on Bio-Rad CFX96Touch™ Real-Time PCR Detection System. The polymerase chain reaction were carried out in triplicates and gene expression was normalized to β-actin. For the analysis of the obtained data, 7500 system SDS software (ABI) was used. The relative mRNA levels were calculated through the 2−ΔΔCT method. The primers were as follows: PYCR1, 5′-GAAGATGGGGGTGAAGTTGA-3′ and 5′-CTCAATGGAGCTGATGGTGA-3′, β-actin, 5′-GTGGACATCCGCAAAGAC-3′ and 5′-AAAGGGTGTAACGCAACTA-3′.
Western blot analysis
Proteins were extracted using 2× sodium dodecyl sulfate (SDS) Sample Buffer and quantified by BCA Protein Assay Kit (Beyotime, Shanghai, China). The target proteins were separated through 10% SDS polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred onto polyvinylidene difluoride (PVDF) membranes. After blocked and washed, the membranes were probed with specific primary antibodies overnight at 4°C, followed by incubated with second antibody Goat anti-rabbit IgG-HRP (Santa Cruz, Dallas, TX) for 4 h. Lastly, the target bands were detected using enhanced chemiluminescence reagents (Amersham, Pittsburgh, PA). The specific primary antibodies were as follows: anti-PYCR1 (1:2000; Abgent, San Diego, CA), anti-E-cadherin (1:1000; BD Biosciences, San Jose, CA), anti-N-cadherin (1:500; BD Biosciences), anti-Vimentin (1:1000; Proteintech, Chicago, IL), anti-Snail (1:1000; Cell Signaling, Danvers, MA), and anti-GAPDH (1:50,000, Proteintech). The relative expressions of target proteins were represented by using the relative gray value (target band gray value/GAPDH gray value) of shCon as 100% and compared the other values with it.
Wound healing assay
95C and H1299 cell lines were cultured in a six-well plate in incubator with 5% CO2 at 37°C. When the cell density reached about 90%, a scraped area was artificially created by a sterile 200 μL pipette tip. After washed with phosphate buffered saline, cells were cultured in serum-low medium (RPMI-1640 medium containing 2% FBS) at 37°C with 5% CO2. The cell-free space was photographed by a microscope after 0 and 20 h for 95C cell line, and after 0 and 36 h for H1299 cell line, respectively. The percentage of wound closure was calculated with ImageJ software (Bethesda, MD).
Transwell assay
Cell suspension (2 × 105 cells/200 μL) was added into the upper chamber without Matrigel (for migration) or with Matrigel (for invasion, Corning, Corning, NY) and then a total of 500 μL of RPMI-1640 medium containing 20% FBS was supplemented into the lower chamber in a 24-well plate. After incubation at 37°C for 24 h, nonmigrated or noninvasive cells on the upper layer of the membrane were removed using a clean cotton swab. Then, the migrated or invasive cells on the under layer of the membrane were fixed in 4% paraformaldehyde, followed by stained in crystal violet (Sigma). Finally, five different fields from each well were selected randomly under a light microscope (Olympus) for photography. Three biological replicates were done and thus 15 photographs were taken in total for each cell line. For calculation, five photographs from each cell line were randomly selected to count the migrated or invasive cells using the counting tool of the Photoshop software (Adobe).
Statistical analysis for quantitative reverse transcription-polymerase chain reaction, wound healing, and transwell assay
The statistical analyses were implemented by GraphPad Prism 5.0 (La Jolla, CA). Student's t-test was applied to determine the difference between shPYCR1 and shCon. Data were presented as mean ± standard error of the mean. *and **indicated the p-values from t-tests are significant at 0.05 and 0.01, respectively.
Results
The abundance of PYCR1 transcripts was elevated in NSCLC specimens
We collected and analyzed PYCR1 expression data from eight lung data sets from the Oncomine platform to explore the expression profiles of PYCR1 in NSCLC specimens and normal lung tissues. As shown in Figure 1, compared with normal lung tissues, PYCR1 mRNA expression was significantly increased in lung adenocarcinoma specimens from the eight lung data sets, which are Su (4-fold), Stearman (7-fold), Selamat (5-fold), Okayama (3-fold), Landi (2-fold), Beer (3-fold), Bhattacharjee (19-fold), and Hou (3-fold). In addition, the abundance levels of PYCR1 transcripts were significantly increased in squamous cell lung carcinoma specimens in Bhattacharjee (12-fold) and Hou (2-fold) lung data sets (Fig. 1). Furthermore, PYCR1 mRNA expression was upregulated to threefold in large cell lung carcinoma in Hou lung data set (Fig. 1). The data indicated that the abundance of PYCR1 transcripts was elevated in all three types of NSCLC specimens.

The expression of PYCR1 transcripts was elevated in NSCLC specimens. PYCR1 mRNA expression was analyzed by eight lung data sets of online Oncomine cancer database, including Su (30 lung samples, 27 lung adenocarcinoma samples, p = 2.90E-08), Sterman (18 lung samples, 20 lung adenocarcinoma samples, p = 1.25E-07), Selamat (58 samples from adjacent nontumor lung tissue, 58 samples from lung adenocarcinoma, p = 2.00E-22), Okayama (20 lung samples, 226 lung adenocarcinoma samples, p = 4.35E-31), Landi (49 lung samples, 58 lung adenocarcinoma samples, p = 2.45E-19), Beer (10 lung samples, 86 lung adenocarcinoma samples, p = 1.00E-14), Bhattacharjee (17 lung samples, 132 lung adenocarcinoma samples, p = 1.74E-05, 21 squamous cell lung carcinoma samples, p = 0.02), and Hou (65 lung samples, 45 lung adenocarcinoma samples, p = 9.34E-14, 27 squamous cell lung carcinoma samples, p = 6.07E-07, 19 large cell lung carcinoma samples, p = 2.74E-06). *p < 0.05; **p < 0.01. p-Values are determined by t-tests. mRNA, messenger RNA; NSCLC, nonsmall cell lung cancer.
PYCR1 depletion suppressed the migration and invasion of NSCLC cells
A previous study in breast cancer cell lines reported that inhibition of PYCR1 significantly reduced the growth and invasion capabilities of the cells. 22 Thus, we had knocked down the endogenic expression of PYCR1 through shRNA to investigate the potential association between PYCR1 and cell metastasis. Two lentiviral shRNA constructions shPYCR1-S1 and shPYCR1-S2 against variant regions of PYCR1 were transfected into two NSCLC cell lines 95C and H1299 to downregulate the expression of PYCR1. Cells transfected by shCon and cells without transfection were employed as controls. As shown in Figure 2A and B, the PYCR1 mRNA expression was greatly decreased in 95C and H1299 cells infected by shPYCR1-S1 and shPYCR1-S2 compared with that in control cells (p < 0.001). And the western blot results indicated that shPYCR1-S1 and shPYCR1-S2 dramatically reduced the protein level of PYCR1 in both 95C and H1299 cell lines (Fig. 2C, D). In addition, shPYCR1-S1 inhibited PYCR1 more efficiently than shPYCR1-S2 in both 95C and H1299 cells (Fig. 2).

PYCR1 was knocked down in 95C and H1299 cells.
After reducing PYCR1 in C95 and H1299, wound healing and transwell invasion assays were performed to verify whether PYCR1 could affect NSCLC cell metastasis in vitro. As shown in Figure 3, the wound healing assay showed that the control cells of 95C and H1299 almost healed the wounds after 20 and 36 h, respectively. However, the healed areas of all cells transfected with shPYCR1-S1 or shPYCR1-S2 were less than half the wounds. After being cultured for 20 h, the healing rates of 95C cells with shPYCR1-S1 or shPYCR1-S2 were 26% and 44%, respectively. Those in H1299 cells with shPYCR1-S1 or shPYCR1-S2 were 30% and 42%, respectively, after 36 h.
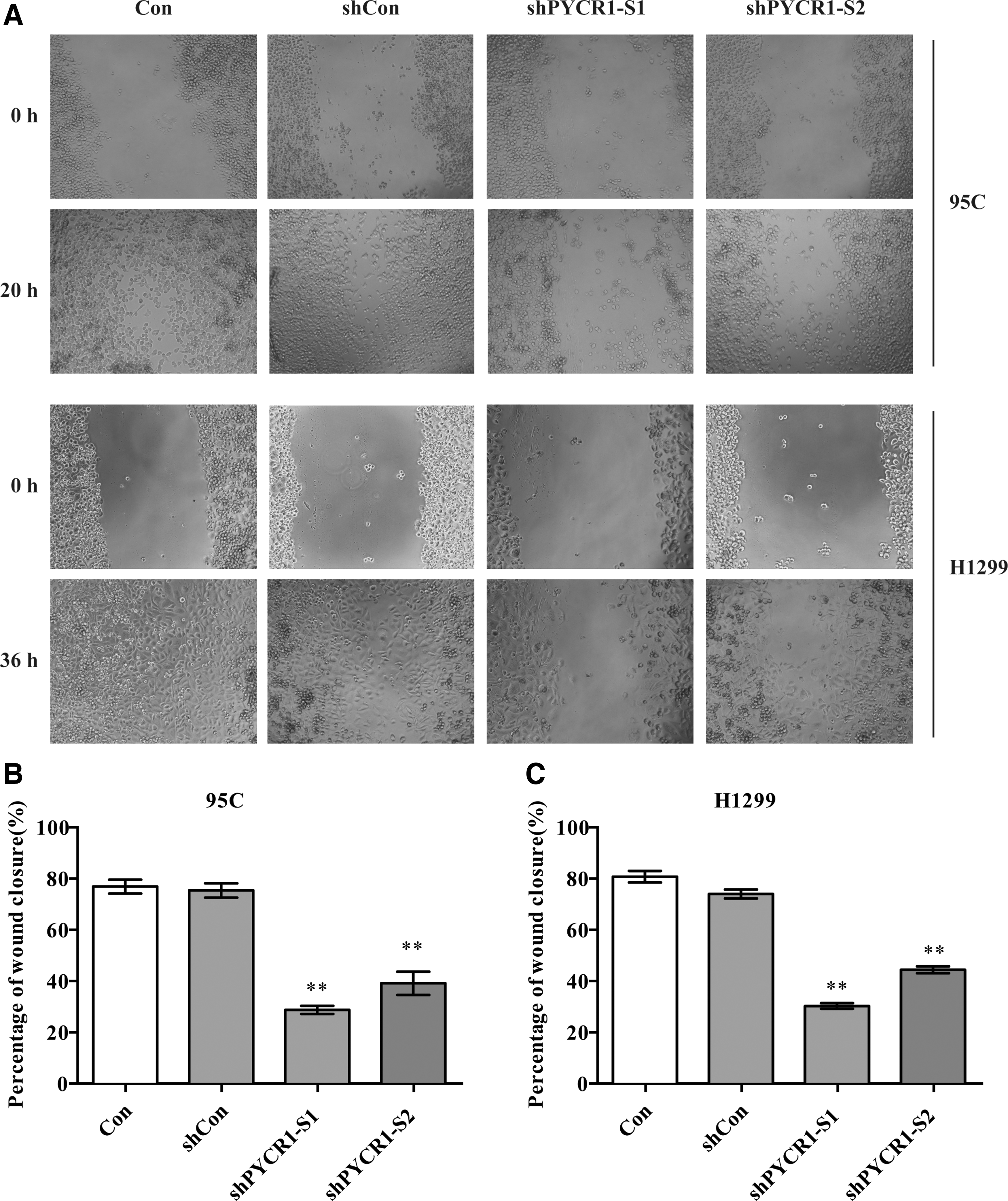
Wound healing assay of PYCR1 depletion NSCLC cells.
In consistent with the wound healing assay, the transwell assay showed that, compared with 95C and H1299 cells transfected with shCon, cells transfected with shPYCR1-S1 or shPYCR1-S2 exhibited much less migrated cells and invasive cells after 24 h (p < 0.001, Fig. 4 and Table 1). In summary, these results indicated that PYCR1 inhibited NSCLC cell metastasis in vitro.
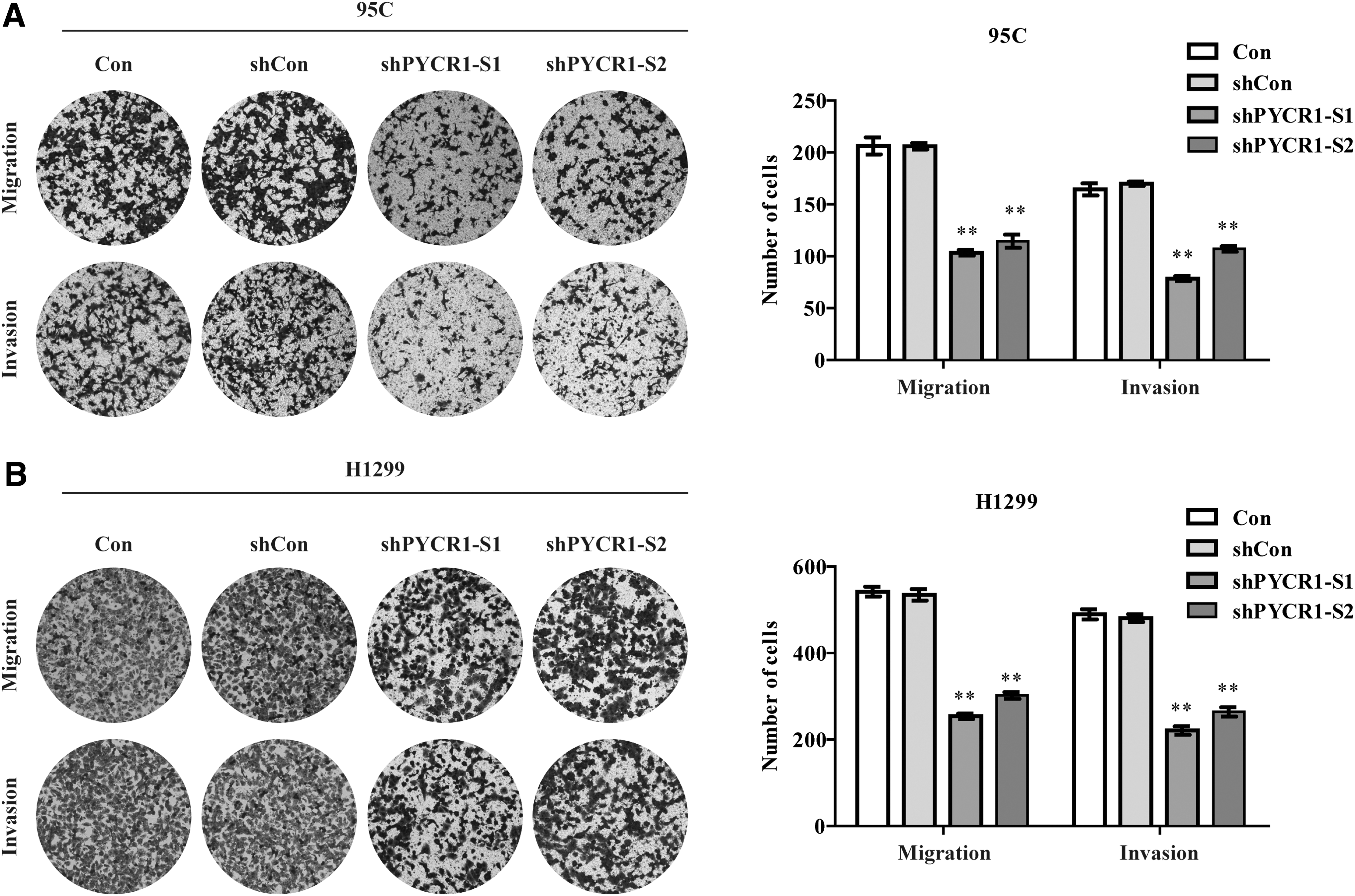
Transwell assay of PYCR1 depletion NSCLC cells.
Transwell Assay of PYCR1 Depletion Nonsmall Cell Lung Cancer Cells
Five different fields of every cell line were selected for calculation. Data presented are mean ± SEM.
Indicates the p-values from t-tests between PYCR1 depletion NSCLC cells and control cells are significant at 0.01.
PYCR1, pyrroline-5-carboxylate reductase 1; NSCLC, nonsmall cell lung cancer; SEM, standard error of the mean.
PYCR1 depletion hampered epithelial-mesenchymal transition in NSCLC cell lines
Epithelial-mesenchymal transition (EMT) is critical for tumor invasion and metastasis. 32,33 To further determine whether knockdown of PYCR1 expression would impact on the activation of EMT pathway, six typical EMT genes were detected using western blot assays. In both 95C and H1299 cells with depleted PYCR1, the protein expression of epithelial cell marker E-cadherin was dramatically increased, whereas the expressions of mesenchymal markers N-cadherin and Vimentin were slightly decreased (Fig. 5). Transcription factor Snail1 represses the transcription of CDH1, which encodes E-cadherin 34,35 and it was obviously reduced in PYCR1-knockdown cells (Fig. 5). The results indicated that PYCR1 depletion promoted E-cadherin expression and inhibited N-cadherin and Vimentin expressions thus hampered EMT in NSCLC.
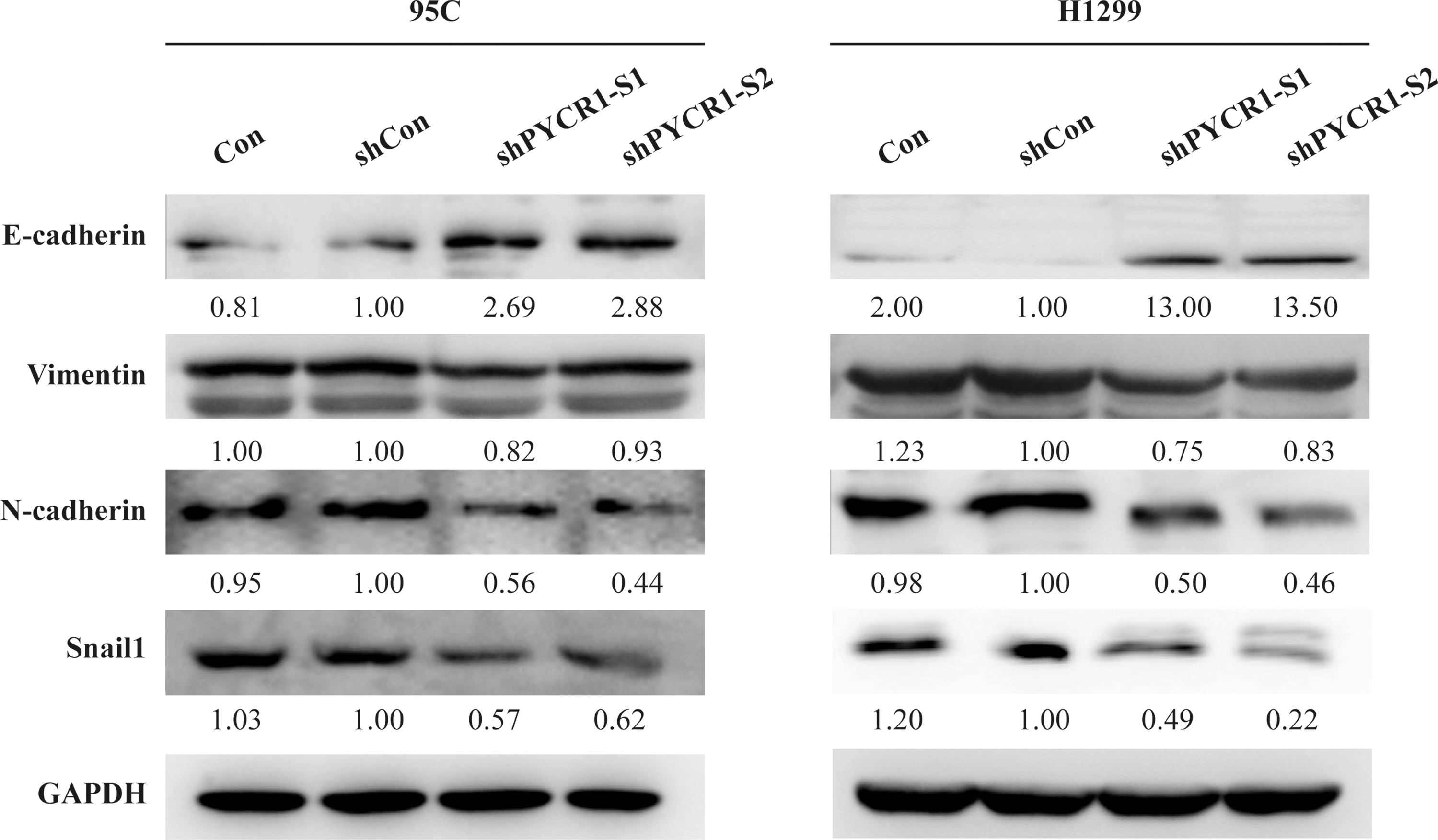
Western blot analysis of EMT-related molecules in PYCR1 depletion NSCLC cells. The expression of E-cadherin, N-cadherin, Vimentin, and Snail1 were analyzed by western blot. Con, wild-type cells; shCon, cell infected by lentivirus with nontarget sequence; shPYCR1-S1 and shPYCR1-S2, cell infected by lentivirus with target sequences S1 and S2, respectively. EMT, epithelial-mesenchymal transition.
Discussion
PYCR1 has been identified as one of the most requisite metabolic genes for tumor genesis in 1981 tumors spanning 19 cancer types. 36 Increasing reports had shown a positive association between the PYCR1 expression and tumor growth in various forms of cancers, including melanoma, breast, prostate, and kidney cancers. 37 –39 In addition, Cai et al. reported high expression of PYCR1 was bound up with higher tumor node metastasis (TNM) stage as well as worse overall survival in patients with NSCLC. 23 The results of this study revealed that knockdown of PYCR1 expression through lentivirus-mediated shRNA slowed down the speed of wound closure and weakened the migrated and invasive capabilities of NSCLC cells, which was consistent with the findings in breast cancer. 22
EMT is a biological process, by which cells suffer a morphological conversion from the epithelial polarization to the mesenchymal fibroblast. 33 EMT implicates the loss of epithelial cell properties and cell–cell adhesions, and the acquiring of migrated and invasive properties. 40 The typical cadherins, such as E-cadherin and N-cadherin, calcium-dependent transmembrane adhesion glycoproteins, associate with actin cytoskeleton. 41 Disorder of cadherins is deemed to be the first critical procedure in the development of EMT. E-cadherins of importance to stable cell adherens junction and signal transduction, and is also a crucial indication of the occurrence of EMT loss. 42 During EMT, epithelial cells decrease E-cadherin and increase N-cadherin to be converted into fusoid mesenchymal cells. 43 Switch from E-cadherin to N-cadherin was reported to play a crucial role in cancer progression. 44 In addition, Vimentin is one of the major cytoskeletal constituents of mesenchymal cells and is strongly linked to differentiation, metastasis, and invasion of cancer cells. 45 Snail1 is a transcription repressor that could inhibit the adherent and tight junctions in epithelial cells to implement EMT, which is strongly associated with an increase of cell migration capacities during tumor progression. 46 In this study, we found that PYCR1 depletion increased E-cadherin expression whereas decreased N-cadherin, Vimentin, and Snail expressions. We speculate that PYCR1 motivated NSCLC migration and invasion through inducing EMT.
Conclusions
In summary, PYCR1 accelerated NSCLC metastasis by promoting EMT pathways in vitro, which could deepen our understanding about the role of PYCR1 in NSCLC progression. The result suggested PYCR1 might be a promising target for treating NSCLC.
Footnotes
Acknowledgment
This study was supported by the Scientific Research Foundation of Guangxi University (No. XGZ160287).
Authors' Contributions
The authors listed as follows have certified that they have made substantial contributions to the conception or design of the study; or the acquisition, analysis, or interpretation of data for the study. They participated in drafting the study or revising it critically for important intellectual content. They did final approval of the version to be published. They agree to be accountable for all aspects of the study in ensuring that questions related to the accuracy or integrity of any part of the study are appropriately investigated and resolved. S.S. conducted experiments, data analysis, and was responsible for the experimental design and preparing the draft. C.Z. and J.S. were in charge of experimental design, data analysis, and revising the article. All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
No competing financial interests exist.