
Abstract
Background:
Glioblastoma is one of the most malignant tumors in the brain with high mortality. In recent years, immunotherapy and targeted therapy show great prospects in the treatments for glioblastoma, whereas more effective therapeutic targets are still urgently needed to be developed. Nucleobindin-2 (NUCB2) is the precursor protein of nesfatin-1, which have a variety of metabolic functions, such as food intake and temperature regulation. In recent years, the potential link between NUCB2 and the development of multiple cancer was gradually revealed; however, the effects of NUCB2 on the progression of glioblastoma are still unclear.
Methods:
Immunohistochemical assays were performed to explore the NUCB2 expression levels in 94 samples of glioblastoma and corresponding nontumor tissues; patients were divided into NUCB2 high expression group and low expression group. Clinical analysis related to the clinical features, the potential link between NUCB2 expression levels, and clinical features were analyzed; the effects of NUCB2 on cell proliferation and invasion of glioblastoma were detected through colony formation and MTT assay, and transwell assay respectively. The possible effects of NUCB2 on tumor growth and metastasis were measured in mice.
Results:
In this study, we demonstrated that NUCB2 over-expression was correlated with the high degree of recurrence of patients with glioblastoma. Further, we also revealed that NUCB2 promoted cell proliferation and invasion of glioblastoma in vitro and promoted the growth and metastasis of glioblastoma in mice.
Conclusion:
This study provided evidence that NUCB2 might be a novel therapeutic target of glioblastoma.
Introduction
Glioblastoma, which belongs to astrocytes tumors, is one of the most malignant tumors in the brain. 1,2 It is estimated that the 5-year survival rate of glioblastoma patients is <4%. 3 The growth manner of glioblastoma is mainly invasive growth, thus aggravating the difficulty of clinical treatment. 4,5 Main treatments for glioblastoma include surgery, radiotherapy, chemotherapy and combined treatment, etc. 6 The surgical treatment is difficult for the total removal, and the recurrent rate is extremely high. 7 Nowadays, the combination therapy obviously improves the 5-year survival rate of patients, whereas glioblastoma is still difficult to cure through this treatment. 8 Therefore, more effective treatment methods are extremely needed. In recent years, immunotherapy and targeted therapy show great prospects. 9 A series of novel molecular targets, such as STAT3 and CD70, are of great significance for the treatment of glioblastoma, but the precise molecular mechanism is a requirement to be further explored. 10,11 More effective therapeutic targets are still urgently needed to be developed.
Nucleobindin-2 (NUCB2), as the precursor protein of nesfatin-1, was first discovered in 2006. 12 The previous functional studies on NUCB2 were focused on its properties of appetite regulation. 13 Subsequently, pharmacological studies indicated the critical role of NUCB2 in a variety of metabolic functions, including glucose homeostasis, the intake of water and food, and the regulation of temperature. 14 –16 In recent years, the potential link between NUCB2 and tumorigenesis was gradually revealed. NUCB2 promotes the development of several cancers, such as prostate cancer, colon cancer, and breast cancer. 17 –19 Studies showed that NUCB2 promotes cancer development due to the promotion of cell proliferation and invasion. 20 In addition, NUCB2 is widely expressed in multiple human tissues, such as pancreas, brain, liver, and kidney. 21 The widespread distribution of NUCB2 suggested the potential involvement of this protein in the development of multiple tumors. However, the effects of NUCB2 on the progression of glioblastoma are still unclear.
Here, we found that NUCB2 expression was upregulated in patients with glioblastoma. Knockdown of NUCB2 dramatically blocked cell proliferation and migration in vitro and further suppressed the growth and metastasis of glioblastoma in mice. Therefore, NUCB2 could represent a novel therapeutic target of glioblastoma.
Materials and Methods
Antibodies, primers, and shRNA plasmids
Anti-NUCB2 (1:1000 dilution for Immunoblot, 1:50 dilution for immunohistochemistry, ab229683; Abcam, Cambridge, United Kingdom), Anti-β-actin (1:1000 dilution, ab8226; Abcam). The qRT-PCR primer sequences of NUCB2 were as follows: forward, 5′-CCTGTGGAAAGTGCGAAGATAG-3′ and reverse, 5′-GCCTCCCACTCTTTATTTCCTC-3′; The qRT-PCR primer sequences of GAPDH were as follows: forward, 5′-GAGTCAACGGATTTGGTCGT-3′ and reverse, 5′-TTGATTTTGGAGGGATCTCG-3′. The shRNA clone for NUCB2 was obtained from the National Core.
Human tissue samples and immunohistochemical analysis
The human glioblastoma tissues were collected from patients treated with surgery in Tianjin Huanhu Hospital. Subsequently, we performed the immunohistochemical (IHC) assays. NUCB2 is mainly expressed in the nucleus region of glioblastoma cells. The 5-μm sections were cut from paraffin blocks, which were used for IHC. Then, tissue sections were performed with citrate buffer (pH = 6.0) and 1.5% hydrogen peroxide in methanol for antigen retrieval. Next, sections were blocked with 2% bovine serum albumin and the slides were incubated with the antibody at 4°C overnight. Then, the second antibody horseradish peroxidase (HRP) was incubated at room temperature for 1 h and diaminobenzidine was added for visualization. The images were collected by a microscope (Nikon, Tokyo, Japan).
All the samples were manually classified into four groups according to the staining intensity (0 = negative level; 1 = low level; 2 = moderate level; 3 = high level). In addition, one score was given according to the percentage of positive cells as: <5% of the cells:1; 6%–35% of the cells: 2; 36%–70% of the cells: 3; and >70% of the cells: 4. According to our score methods, the score of staining intensity × score of stained cells percentage ≤4 as low staining and >4 as high staining. The sections of each patient were observed within six visual fields at least, and two experienced pathologists read the sections.
Cell culture and transfection
U251 and U87 human glioblastoma cells were bought from ATCC and maintained in our lab. Both U251 and U87 cells were cultured in ATCC-formulated Eagle's Minimum Essential Medium (No. 30-2003) culture medium and supplemented with 10% of fetal bovine serum and incubated at 37°C in a 5% CO2 incubator.
The shRNA plasmid of NUCB2 were transfected into glioblastoma cells by using lipofectamine 3000 (L3000015; Invitrogen, Carlsbad, CA). NUCB2 stable depletion U251 cells were screened by shRNA lentivirus infection and used for the in vivo assays.
Quantitative PCR assay
Total RNA was extracted from U251 and U87 cells through the use of Trizol reagent (15596-018; Invitrogen). Then, total RNA in cells was reverse-transcribed by using M-MLV reverse transcriptase (M1701; Promega, Madison). Quantitative real-time PCR was performed by using SYBR mixture (RR420A; Takara, Japan), and the relative expression level of NUCB2 was normalized to GAPDH.
Immunoblot assays
Total protein samples were extracted from both U251 and U87 cells, and the indicated proteins such as NUCB2 were separated and analyzed through sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Subsequently, the polyvinylidene fluoride membranes were transmembraned for 2 h, blocked with 5% dry milk for 2 h, and incubated with the specific primary antibodies for the detection of NUCB2 and β-actin for 1.5 h. Then, the membranes were incubated with HRP-conjugate secondary antibodies for 1 h. Interest signals were visualizal with an ECL kit.
Cell proliferation assays
For colony formation assay, ∼1000 cancer cells were seeded into a six-well culture plate, transfected with the indicated shRNA plasmids, and maintained at 37°C for 48 h. Cells were subsequently fixed with 4% paraformaldehyde (PFA) at room temperature for 25 min and stained with 0.1% crystal violet at room temperature for another 30 min. Then, cells were washed twice with phosphate-buffered saline (PBS). Colony numbers were manually counted.
For MTT assays, cells were plated in 96-well plates with a density of 500 cancer cells for each well, transfected with the indicated shRNA plasmids, and cultured for 48 h. Cells were then incubated with MTT for 3 h, and the medium was removed. Cells were then washed with PBS twice to remove the remaining MTT. Stained cells were extracted by 150 μL dimethylsulfoxide, and the OD value was quantified with a microplate reader at 570 nm wavelength.
Cell motility assays
For transwell assays, U251 and U87 cells were transfected with control or NUCB2-shRNA plasmids and resuspended in serum-free medium after 2 d. The upper chambers of transwell filters were coated with 20% matrigel, and the filters were incubated at 37°C for 1 h. A total number of 105 cells in 100 μL of serum-free medium were then seeded into the chambers of the inserts to induce the migration toward the bottom chambers, which contained complete culture medium. After 48 h, underside cells were fixed in 4% PFA and stained with 0.1% crystal violet for 30 min, and photographs were then taken to count the cells.
For wound closure assays, both U251 and U87 cells were transfected with the indicated shRNA plasmids and grown for 48 h to induce confluent monolayers. Then, the wound was mechanically made by using a 20-μL pipette tip. Cell debris was effectively washed by PBS buffer twice, and the complete culture medium was added and maintained for 24 h. Photographs were taken at 0 and 24 h, and the extent of wound closure was measured.
In vivo tumor growth and metastasis assays
Nude BalB/c mice (6–8 weeks, 18–22 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). All animal assay procedures were approved by our Institutional Animal Care and Use Committee.
For tumor growth assay, U251 cells were first stably transfected with control or NUCB2 shRNA lentivirus. Approximately 2 × 106 cells were implanted into athymic nude mice. After 2 weeks, tumors were isolated and photographed; the tumor volume was measured each week to get the tumor growth curve.
For metastasis assay, nearly 1 × 106 U251/shControl, U251/shNUCB2 stable depletion cancer cells were resuspended in 100 μL PBS buffer and then injected through the tail vein to induce the lung metastasis. Mice were raised for 4 weeks. Subsequently, all mice were sacrificed and the lungs of mice were isolated and photographed. The levels of pulmonary metastases were then measured.
Statistics
Data were analyzed with SPSS 22.0 software and shown as the mean ± standard deviation in this study. Student's t-test was used for statistical comparisons, and p < 0.05 is considered significant. The potential correlation between NUCB2 expression levels and clinical characteristics was studied through the χ 2 test. “*” indicates p < 0.05.
Results
NUCB2 is highly expressed in glioblastoma samples and its clinical significance
To explore the potential role of NUCB2 in the growth and development of glioblastoma, bioinformation analysis was performed in an interactive web server GEPIA with the sequencing expression data of 163 tumors. mRNA expression level of NUCB2 in GBM showed significantly higher than normal tissues (p < 0.05, number = 163, normal tissues number = 207, Fig. 1A). Then, IHC assays were performed, and the expression of NUBC2 in tissue samples surgically removed from 82 patients with glioblastoma in our hospital was detected. Notably, the results showed that NUCB2 was mainly localized in the nucleus and highly expressed in the glioblastoma tissue (Fig. 1B). According to the expression level of NUCB2, samples are divided into two groups: high and low NUCB2 expression groups (Fig. 1B). As a comparison, NUCB2 showed obvious low expression in the adjacent tissues (Fig. 1C), suggesting that NUCB2 might play a role in the growth and development of glioblastoma.
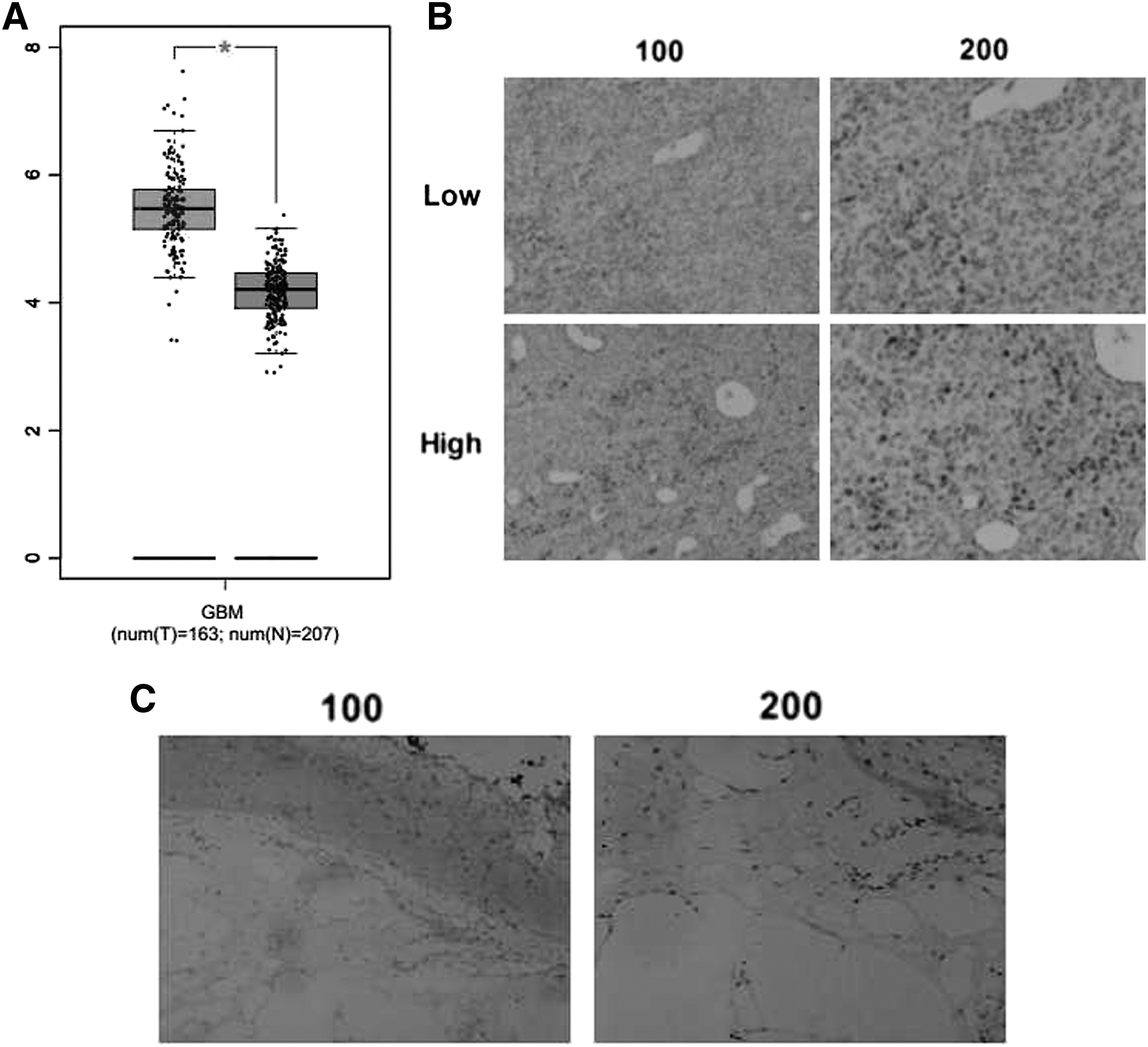
NUCB2 expression was correlated with the poor prognosis of patients who underwent glioblastoma.
The analysis of clinicopathological characteristic showed the difference between low and high NUCB2 expression groups. Interestingly, we noticed that NUCB2 expression in the glioblastoma tissues was remarkably related with the degree recurrence, suggesting a possible clinical link between NUCB2 expression and glioblastoma (Table 1). However, there were no marked differences between high and low NUCB2 groups with regard to other clinical feature aspects, such as patient age, gender, tumor lateralization, or IDH1 mutations (Table 1).
Relationships between Nucleobindin-2 and Clinicopathological Characteristics in Ninety-Four Patients with Glioblastoma
NUCB2, nucleobindin-2.
p < 0.05.
NUCB2 depletion blocked cell proliferation and invasion of glioblastoma
Abnormal cell proliferation and migration promotes tumor occurrence and metastasis. Therefore, we examined whether the effects of NUCB2 on glioblastoma were due to the promotion of cell proliferation or migration.
We then used an NUCB2-targeted shRNA to deplete its expression in two types of human glioblastoma cells: U251 and U87 cells, respectively. The silencing efficiency of NUCB2-targeted shRNA was detected through both quantitative PCR and immunoblot assays. As expected, results indicated that NUCB2 expression level obviously dropped in NUCB2-shRNA transfected U251 and U87 cells (Fig. 2A, B).

NUCB2 was effectively knocked down in U251 and U87 cells.
We further explore the role of NUCB2 in cell proliferation of glioblastoma by colony formation and MTT assays. From the staining results, we found that the colony formation capacity was significantly restrained by NUCB2 depletion, consistent with the obviously decreased colony numbers. (Fig. 3A). In addition, the results of MTT assays showed that NUCB2 ablation resulted in a significant decreased absorbance value at 570 nm wavelength in both U251 and U87 cells, suggesting a decline in proliferative capacity (Fig. 3B).

NUCB2 promotes the proliferation and invasion of glioblastoma cells in vitro.
Further, we studied the effects of NUCB2 on the migration and invasion of glioblastoma cells. Interestingly, knockdown of NUCB2 inhibited the extent of wound closure in both U251 and U87 cells (Fig. 3C). Through transwell assays, both U251 and U87 cells exhibited a markedly low invasive capacity through the membranes caused by NUCB2 ablation, with obviously decreased cell numbers (Fig. 3D).
Together, we demonstrated the involvement of NUCB2 in the regulation of cell proliferation and invasion of glioblastoma in vitro.
NUCB2 promotes the growth and metastasis of glioblastoma in mice
To explore the relationship between NUCB2 expression and glioblastoma in mice, U251 cells were infected with NUCB2 shRNA lentivirus and subsequently injected into nude mice. After 2 weeks following injection, tumors began to develop, and tumor volume was measured each week. According to the results, the tumor volume of the NUCB2 ablation group was significantly smaller than control (Fig. 4A).
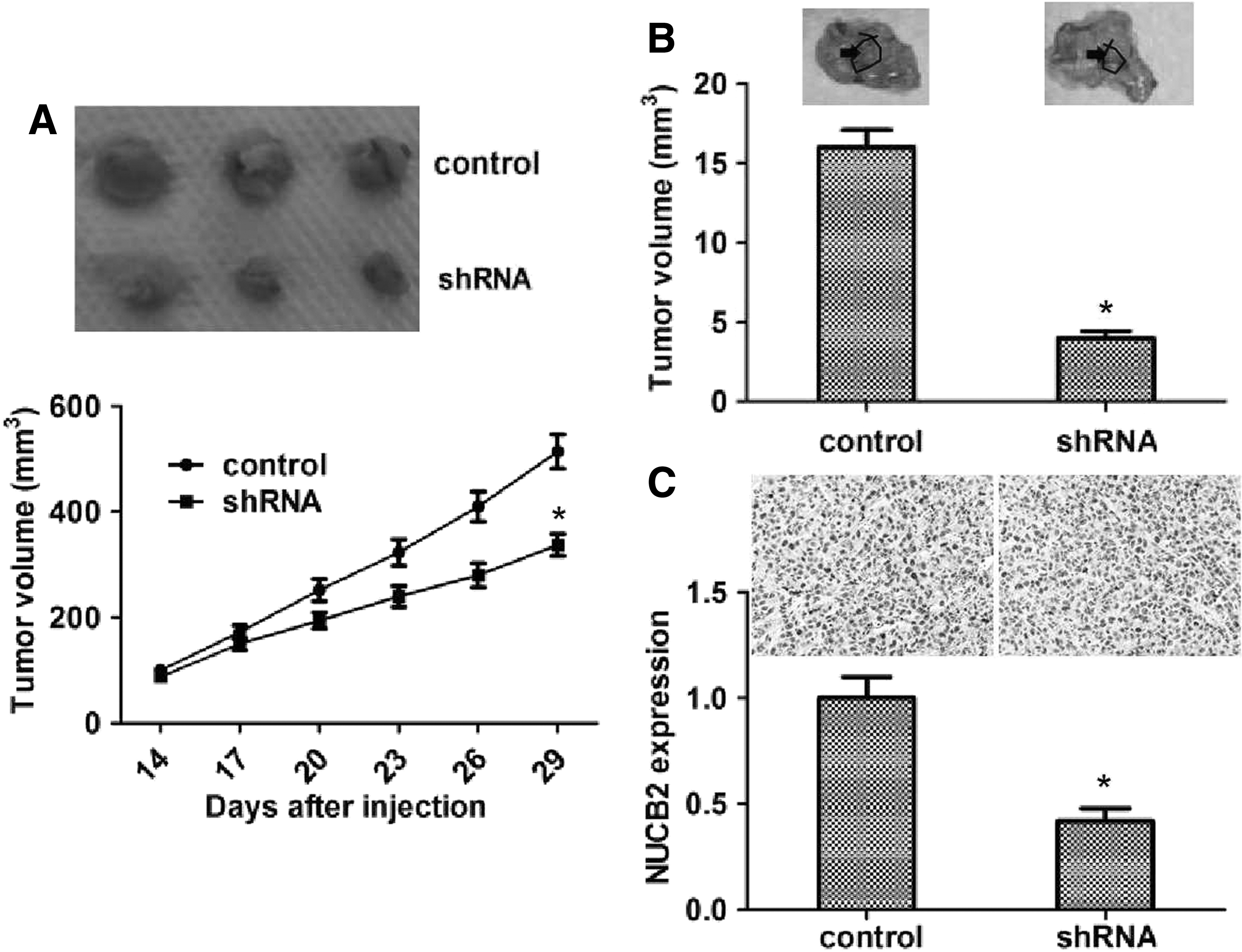
Knockdown of NUCB2 impaired the growth and metastasis of glioblastoma in mice.
Based on previous results, we performed lung metastasis assay in mice and found that the incidence of lung metastasis of NUCB2 knockdown U251 cells was obviously decreased, compared with control (Fig. 4B).
We then detected the expression level of NUCB2 in tumor tissues by immunoblot and immunohistochemistry assays. Expectably, data showed that the NUCB2 expression level in the depletion group was obviously decreased compared with control (Fig. 4C). Collectively, these data demonstrated that NUCB2 plays a critical role in the growth and metastasis of glioblastoma.
Discussion
The diagnosis and treatment of glioblastoma is still a worldwide problem. Due to the high invasive and poor prognosis, glioblastoma has the second highest mortality, second only to hepatocellular carcinoma. 22 Globally, glioblastoma is still lacking effective treatment methods. 23 The combination treatment of surgery and chemotherapy improved the patients' survival rate. 24 However, due to the high recurrence rate, glioblastoma is still in urgent need of more effective treatments. 25 Nowadays, targeted therapy has broad prospects. 26 Recently, the pathogenesis of glioblastoma has been deeply studied, and multiple molecular targets have been found. 27 For an example, KPNB1 promotes the development of glioblastoma through the inhibition of cell apoptosis. 28 Here, we demonstrated that NUCB2 was associated with the poor prognosis of glioblastoma, providing a potential therapeutic target for this disease. However, more sophisticated regulatory mechanisms need further study.
A previous study demonstrated that a high expression level of NUCB2 in prostate cancer was significantly associated with the recurrence-free survival rate, and NUCB2 could be a prognostic factor in patients with prostate cancer. 17 Previous studies also indicated that NUCB2 expression level was associated with the poor prognosis of patients with renal cell carcinoma. 29 Interestingly, we found that NUCB2 expression was positively associated with overall survival rate and disease-free survival rate of patients with glioblastoma. NUCB2 may have critical effects on the prognosis of different tumors.
We also found that the expression of NUCB2 was correlated with clinical features, including the degree of tumor resection and tumor recurrence, which induced the high metastasis and poor prognosis of glioblastoma. Several studies have indicated poorer prognosis in younger colorectal cancer patients compared with older patients, suggesting the critical effect of patient age. 30 However, there was no obvious link between NUCB2 expression and other clinical characteristics of patients with glioblastoma, such as patient age and tumor stage.
NUCB2 is widely distributed in the brain and is believed to be involved in multiple brain functions, such as the stress response via activation of the HPA axis. 31 Notably, although NUCB2 has important function in the brain, we found that high expression of NUCB2 promoted the poor prognosis of glioblastoma. Our data further proved that NUCB2 promoted cell proliferation and invasion of glioblastoma. Previous studies showed that NUCB2 played critical roles in metabolism, such as food intake, glucose homeostasis, and appetite regulation. 14,21 However, the precise role of NUCB2 in the regulation of both cell proliferation and invasion was still unclear. In addition, high expression of NUCB2 was related to the poor prognosis of multiple cancers, including gastric cancer and bladder cancer, due to the promotion of cell proliferation and migration. 17,20 NUCB2 inhibited the proliferation of ovarian cancer cells arresting cell cycle and promoted cell apoptosis, which could be blocked by the mTOR signaling pathway. 32 It has been reported that NUCB2 promoted invasion and EMT of colon cancer cells through the LKB1/ZEB1 pathway. 18 The modecluar mechanism underlying NUCB2 regulating the proliferation and migration of tumor cells is worth further research. Also, NUCB2 promoted cell apoptosis of renal cell carcinoma, suggesting that NUCB2 has multiple effects on cancer development. 29,33 Whether NUCB2 promotes the poor prognosis of glioblastoma through inhibiting apoptosis also needs further study.
In concusion, an obvious link was detected between the expression level of NUCB2 and clinical outcome or features of glioblastoma. This strongly suggested that NUCB2 is an oncogene that is correlated with the progression of glioblastoma. Through the analysis of clinical samples, in vitro assays, and animal assays, we declared NUCB2 as a novel therapeutic target of glioblastoma.
Footnotes
Ethical Approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Author Contributions
Conceived and designed the experiments: L.-B.W. and Q.-J.L. Performed the experiments: L.-B.W., J.-X.L., and J.L. Contributed reagents/materials/analysis tools: J.-X.L. and X.-B.Z. Wrote the article: L.-B.W. and J.L.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This work was supported by Tianjin Public Health Bureau grants (2013KG124).