
Abstract
Backgrounds:
Thyroid cancer (
Materials and Methods:
RT-qPCR and western blot were used to detect gene expression levels. Cell counting kit-8 (CCK-8) and ethynyl-2′-deoxyuridine (EdU) assays were used to determine proliferation. Caspase-3 activity assay was used to examine apoptosis. Intermolecular interaction was investigated by luciferase reporter assay and chromatin immunoprecipitation (ChIP) assay.
Results:
We confirmed the elevation of LINC01410 expression in
Conclusions:
This study demonstrated that LINC01410/miR-3619-5p/FOXM1 positive feedback loop regulated cell proliferation and apoptosis in
Introduction
Thyroid cancer (TC) is a prevalent carcinoma in endocrine system characterized with enormous heterogeneity. 1 The most common subtype of TC is papillary thyroid carcinoma (PTC), taking up around 80%–85% of TC cases in adults. 2 Past decades have witnessed the rapid increase in the morbidity of PTC. 3,4 Although prognosis of early PTC patients is satisfactory, 5 patients with advanced PTC only have a 5-year survival rate of <59%. 6,7 This necessitates the in-depth research on mechanism of PTC progression.
Long noncoding RNAs (lncRNAs), a subset of RNA molecules longer than 200 nucleotides, are short of obvious protein-coding capacities. 8,9 LncRNAs have been discovered to express aberrantly in a diversity of cancers, 10,11 including in PTC. 12,13 Long intergenic non-protein coding RNA 1410 (LINC01410) was first reported to promote the angiogenesis and metastasis in gastric cancer. 14 Later, another study illustrated that LINC01410 exerted oncogenic effect in colon cancer by promoting proliferation and invasion. 15 However, the correlation of LINC01410 with PTC has never been investigated.
The regulatory mechanisms of lncRNAs are diverse. In cytoplasm, lncRNAs are discovered to realize their biological functions in cancers through acting as competing endogenous RNAs (ceRNAs) to sponge microRNAs (miRNAs). 16,17 The miRNAs are known as small noncoding RNAs with 21–25 nucleotides, acting as suppressors of gene expression through targeting the 3′ untranslated regions (3′UTR) of the target genes. 18 miR-3619-5p has been reported to perform as a tumor suppressor in bladder carcinoma, 19 prostate cancer, 20 and non-small cell lung cancer. 21 However, it has never been explored in PTC. The lncRNA-mediated ceRNA network has been reported in PTC. 22 Also, LINC01410 has been reported to sponge miR-3128 and miR-532 to exert oncogenic effect in several cancers, 14,15 but its interaction with miR-3619-5p has never been revealed before.
Forkhead box M1 (FOXM1) is one of the transcription factors in forkhead box family. 23 FOXM1 has been demonstrated to facilitate cell cycle progression through regulating the transition of G1/S and G2/M, and it also has been stated as the modulator of chromosomal stability. 24 Emerging studies have proved the participation of FOXM1 in regulating cell proliferation and metastasis in human cancers. 25,26 Furthermore, the oncogenic role of FOXM1 has also been suggested in PTC. 27 Additionally, as a transcription activator, FOXM1 has been illustrated to regulate the expression of lncRNA at transcription level in human cancers. 28 However, its relation with LINC01410 and miR-3619-5p in PTC remains unclear. In summary, the purpose of our study is to investigate the function of LINC01410 and its regulation on FOXM1 in PTC.
Materials and Methods
Cell lines and cell culture
The normal human thyroid epithelium cell line (Nthy-ori 3-1) and PTC cell lines (TPC, BHP5-16, BHP2-7, and K1) were bought from the tumor cell bank of the Chinese Academy of Sciences (Shanghai, China). Cell culture was carried out utilizing Roswell Park Memorial Institute (RPMI)-1640 (Thermo Fisher Scientific, Inc., Waltham, MA) with the supplementation of 10% fetal bovine serum (Gibco BRL, Grand Island, NY), penicillin (100 U/mL), and streptomycin (100 mg/mL) (Invitrogen; Thermo Fisher Scientific, Inc.). The medium was kept in humid air containing 5% CO2 at 37°C.
Cell transfection
LINC01410 was silenced by small interfering RNA (siRNA; GenePharma, Shanghai, China) targeting LINC01410 (siLINC01410#1 and siLINC01410#2), with the scramble siRNAs acting as negative controls (siNC). FOXM1 was overexpressed by pcDNA3.1 inserted with the full-length sequences of FOXM1 (pcDNA3.1/FOXM1), with pcDNA3.1 vector (Invitrogen; Thermo Fisher Scientific, Inc.) as control. The overexpression and knockdown of miR-3619-5p were realized respectively by miR-3619-5p mimic and miR-3619-5p inhibitor (GenePharma), with NC mimic and NC inhibitor as controls. These plasmids were introduced as demanded into the BHP5-16 and TPC cells with the application of Lipofectamine 2000 Reagent (Invitrogen).
RT-qPCR
RNA extraction was accomplished by using TRIzol reagent (Invitrogen, Carlsbad, CA) obeying the manufacturer's protocol. The complementary DNA (cDNA) was reverse transcribed from equal amount of RNAs by the utilization of the SuperScript Reverse Transcriptase Kit (Vazyme, Nanjing, China). Analysis of expressions of LINC01410, FOXM1, and miR-3619-5p was conducted employing SYBR Green PCR Master Mix (Vazyme) with the Fast Real-time PCR 7300 System (Applied Biosystems, Foster City). Analysis of data was on the basis of 2−ΔΔCT method. LINC01410 and FOXM1 expressions were normalized to GAPDH, and miR-3619-5p expression was normalized to U6. Sequences of primers were as follows:
LINC01410 F GTGACAAGAATGGCCCAAGC
LINC01410 R ACTGTGCACCTGTTACACCA
FOXM1 F ATACGTGGATTGAGGACCACT
FOXM1 R TCCAATGTCAAGTAGCGGTTG
U6 F GCTTCGGCAGCACATATACTAAAAT
U6 R CGCTTCACGAATTTGCGTGTCAT
GAPDH F GGGAGCCAAAAGGGTCAT
GAPDH R GAGTCCTTCCACGATACCAA.
Cell counting kit-8
The proliferation of PTC cells was evaluated by the commercially available cell counting kit-8 (CCK-8) assay (Dojindo, Tokyo, Japan). When cells seeded in the 96-well plates (5 × 103 cells/well) were incubated for 0, 1, 2, 3, or 4 d, each well was added with the CCK-8 solution (10 μL). Following the 4 h incubation, the absorbance was determined at 450 nm wavelength under the microculture plate reader (FLUOstar OPTIMA, BMG, Germany).
Ethynyl-2′-deoxyuridine
Ethynyl-2′-deoxyuridine (EdU) assay was conducted with the application of the EdU Apollo DNA in vitro kit (RiboBio Cell Cycle Analysis, Guangzhou, China), on the basis of the manufacturer's instructions. In short, PTC cells underwent incubation for 1 d in each well of a 96-well plate added with 10 μM EdU. Subsequently, cells were subjected to fixation utilizing 4% polyformaldehyde (100 μL) in phosphate-buffered saline (PBS, pH: 7.2–7.4). Then, cells went through incubation with 2 mg/mL glycine solution and permeabilization in the permeabilization buffer (100 μL, 0.5% Triton X-100 in PBS). Thereafter, PTC cells were subjected to the treatment of 1 × Apollo solution (100 μL) and then the treatment with DAPI solution (100 μL) in the dark. After the PBS (pH: 7.2–7.4) washing, samples were analyzed utilizing fluorescence microscopy.
Caspase-3 activity assay
C1115 caspase-3 assay kit (Beyotime, Shanghai) was utilized for the measurement of caspase-3 activity. In short, PTC cells were collected 48 h after indicated transfection, washed by PBS (pH: 7.2–7.4), and then resuspended utilizing cold lysis buffer. Five microliters of caspase-3 substrates were added in the supernatant following the centrifugation. Then, the mixture underwent 4 h incubation in the dark at 37°C. The measurement of absorbance (OD value) was conducted by the use of microplate reader (Infinite M200; Tecan, Männedorf, Switzerland) to analyze cell apoptosis. The OD value of each well was measured at 405 nm. Caspase-3 activity was calculated in arbitrary units using the formula: absorbance of (sample−control)/control. Standard curves were generated and illustrated in Supplementary Table 1 and 2.
Subcellular fractionation
The nuclear and cytosolic fractions of PTC cells were separated utilizing the PARIS Kit (Life Technologies, Norwalk, CT) obeying the manufacturer's instructions. U6 and GAPDH were applied as the fractionation indicators for the quantification PCR.
Dual-luciferase reporter assay
To examine the interaction of miR-3619-5p with LINC01410 and FOXM1, the 3′UTR of LINC01410 or FOXM1 containing the bindings sites for miR-3619-5p were generated into the pMIR-RB-REPORT™ vectors (Ribobio Cell Cycle Analysis), which were named as WT-LINC01410 and WT-FOXM1. The 3′UTR of LINC01410 and FOXM1 containing the mutant sites were generated into pMIR-RB-REPORT vectors (Ribobio Cell Cycle Analysis), which were named as Mut-LINC01410 and Mut-FOXM1. MiR-3619-5p mimics or NC mimics were co-transfected with WT-LINC01410 or Mut-LINC01410, or WT-FOXM1 or Mut-FOXM1 into 293T cells respectively. To examine the binding of FOXM1 to the LINC01410 promoter, the promoter sequences of LINC01410 were generated into the luciferase reporter vector pGL3-basic (Promega, Madison WI), and the plasmids were co-transfected with pcDNA3.1 or pcDNA3.1/FOXM1 into 293T cells. Forty-eight hours subsequent to the transfection, luciferase activities were evaluated by the utilization of Dual-Luciferase Assay System (Promega). Renilla luciferase activity was used as normalized control.
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) assay was implemented with the utilization of ChIP Assay Kit (Beyotime Institute of Biotechnology, Jiangsu, China). In short, the cross-linked chromatin was fixed for 10 min with the utilization of 1% formaldehyde at 37°C. Then, DNA underwent sonication to be separated to fragments with the average length of 200–1000 bp. The immunoprecipitation was carried out with the utilization of antibodies against FOXM1 (ab245309; Abcam, Cambridge, UK) or IgG (ab172730; Abcam) as negative control. Finally, the precipitated DNA was examined by RT-qPCR.
Western blot
Cell lysates were obtained by the use of radioimmunoprecipitation assay lysis buffer with the addition of protease inhibitors (Hoffman-La Roche Ltd., Basel, Switzerland). Quantification of protein density was conducted by Bradford Protein Assay Kit (Thermo Fisher Scientific, Inc.). Then, proteins (30 μg) underwent separation with the utilization of 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). After the proteins were transferred onto the polyvinylidene fluoride (PVDF) membrane (EMD Millipore, Billerica, MA), membranes were sealed in 5% nonfat milk. To probe the membranes, primary antibodies against FOXM1 (ab245309) or GAPDH (ab8245) (all from Abcam) were used for the overnight incubation. Thereafter, membranes were incubated for 2 h with horseradish peroxidase-linked secondary antibody. To visualize the signal intensity, an electrochemiluminescence kit (Pierce Biotechnology, Rockford, IL) was applied.
Statistical analysis
Data were interpreted and the statistics were analyzed with the use of SPSS 17.0 software (SPSS, Inc., Chicago, IL). Data from experiments conducted thrice were presented as the mean ± standard deviation. Student's t-test or one-way ANOVA followed by Turkey's multiple comparison test were used to compare the differences of two or more groups. p < 0.05 was utilized to suggest the statistically significant difference.
Results
Silencing LINC01410 retarded cell proliferation and facilitated apoptosis in PTC
To investigate the role of LINC01410 in PTC, we first detect the expression of LINC01410 in PTC cell lines. We found through RT-qPCR that LINC01410 was expressed highly in four PTC cell lines in comparison with the normal cell line (Fig. 1A). Based on the upregulation of LINC01410 in PTC cells, we selected two PTC cell lines that exhibited the highest LINC01410 level (BHP5-16 and TPC) to silence the endogenous expression of LINC01410 for loss-of-function assays. The results of RT-qPCR confirmed the significant reduction of LINC01410 expression in both cell lines after transfection of siLINC01410#1 or siLINC01410#2 (Fig. 1B).
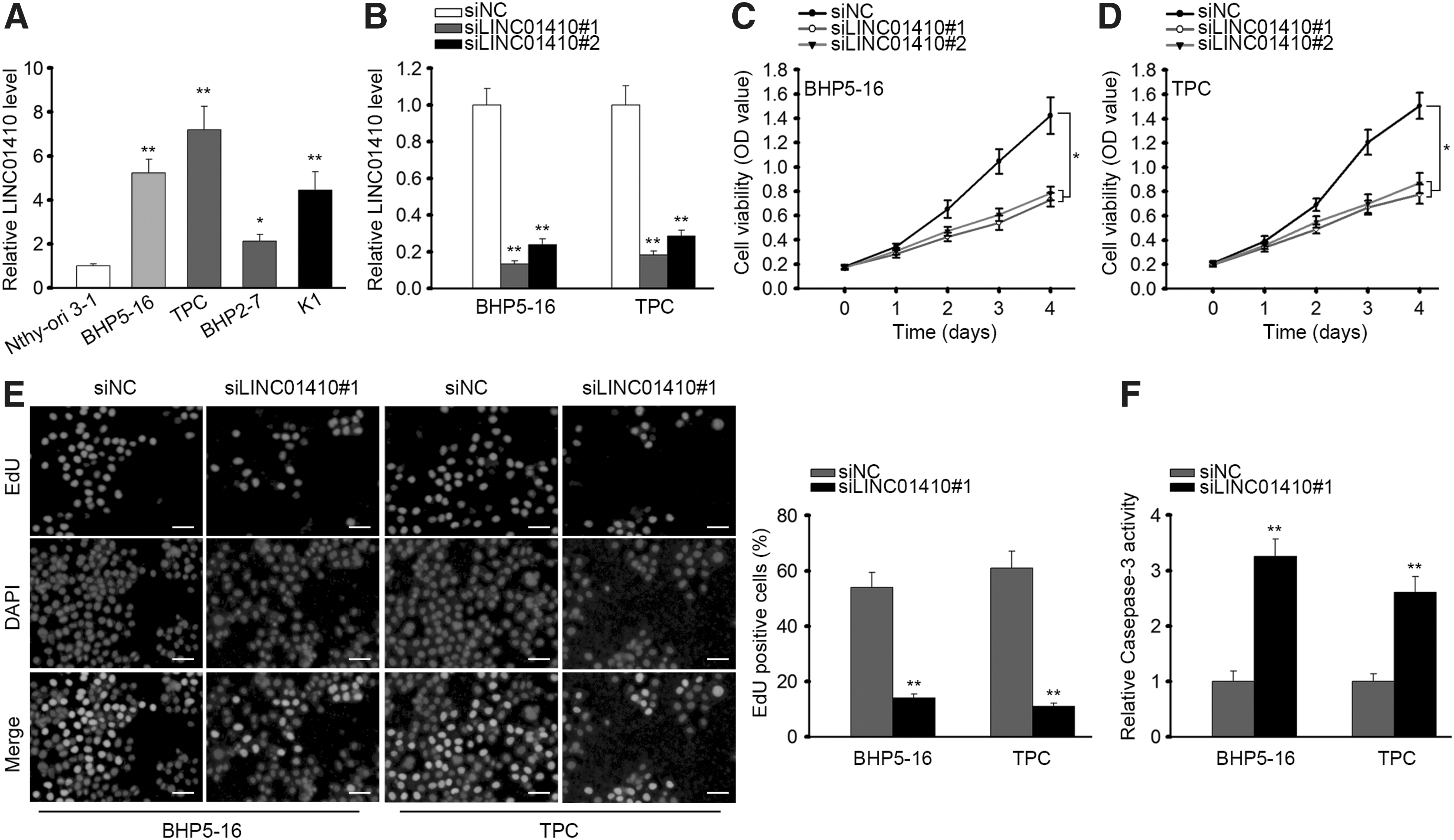
Silencing LINC01410 retarded cell proliferation and facilitated apoptosis in
Next, we observed via CCK-8 silencing LINC01410 attenuated cell proliferation in PTC (Fig. 1C). Since we have validated through RT-qPCR that siLINC01410#1 presented better knockdown efficiency and confirmed through CCK-8 assay that siLINC01410#1 had more significant inhibitory effect on cell proliferation, we used siLINC01410#1 for subsequent assays. EdU assay revealed that LINC01410 knockdown reduced the EdU-positive cell ratio in PTC cells (Fig. 1D). Additionally, we detected caspase-3 activity to examine the effect of silencing LINC01410 on apoptosis of PTC cells. Consequently, we found that LINC01410-silenced PTC cells presented elevated caspase-3 activity compared with control group (Fig. 1E). These data suggested that LINC01410 was upregulated in PTC cells and that silencing LINC01410 attenuated proliferation and induced apoptosis in PTC cells.
LINC01410 positively regulated FOXM1 expression and miR-3619-5p potentially interacted with LINC01410 and FOXM1 in PTC
It is widely known that lncRNAs could exert their impacts in human cancers through activating certain oncogenes. 8 FOXM1 has been newly discovered to express highly and positively regulated proliferation and inhibited apoptosis in PTC. 27 Therefore, we speculated that LINC01410 might had an impact on FOXM1 expression in PTC. First, we confirmed that FOXM1 exhibited an elevated level in PTC cell lines (Fig. 2A). Subsequently, we found through RT-qPCR and western blot results that silencing LINC01410 could downregulate the expression of FOXM1 at mRNA and protein levels (Fig. 2B).
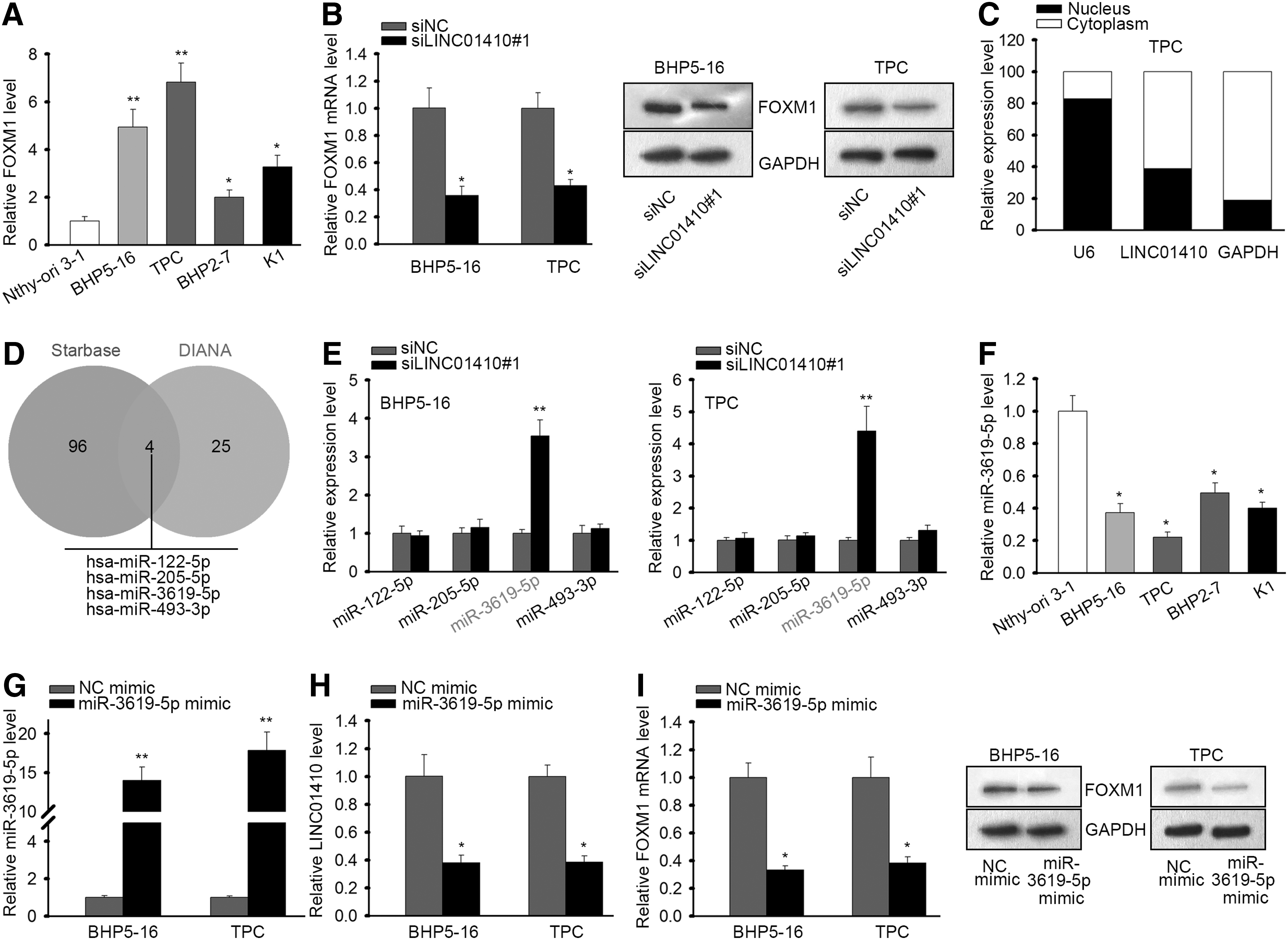
LINC01410 regulated FOXM1 expression and miR-3619-5p potentially interacted with LINC01410 and FOXM1 in PTC.
It is axiomatically acknowledged that the manners whereby lncRNAs regulated gene expression are different according to the localization of their expression. 29 Therefore, we detected the cellular localization of LINC0410 in PTC cells. As a result, LINC01410 was mainly expressed in cytoplasm (Fig. 2C). Because in cytoplasm, lncRNAs could regulate gene expression through performing as ceRNAs, 16,17 and previous studies have reported that LINC01410 could function through ceRNA network in gastric cancer and colon cancer, 14,15 we hypothesized that LINC01410 could also regulate FOXM1 expression through acting as a ceRNA. Hence, we tried to identify the miRNA interacting with both LINC01410 and FOXM1.
We browsed lncBase and picked the 25 miRNAs that had the highest binding score with LINC01410, and browsed starbase and identified 96 miRNAs interacting with FOXM1. The intersection of the results of two databases showed four miRNAs (miR-122-5p, miR-205-5p, miR-3619-5p, and miR-493-3p) potentially interacting with both LINC01410 and FOXM1 (Fig. 2D). To find out the miRNA actually involved in the ceRNA network regulated by LINC01410, we detected the expressions of four miRNAs upon LINC01410 silence. It turned out that in both BHP5-16 and PTC cells, only miR-3619-5p expression was induced by LINC01410 knockdown (Fig. 2E), indicating that miR-3619-5p might be implicated in the LINC01410-regulated ceRNA network. Additionally, we identified the significant downregulation of miR-3619-5p in PTC cell lines (Fig. 2F).
Moreover, we overexpressed miR-3619-5p to evaluate its effect on LINC01410 and FOXM1 expressions. RT-qPCR results validated the overexpression of miR-3619-5p by the transfection of miR-3619-5p mimic (Fig. 2G). We then found that ectopic expression of miR-3619-5p reduced LINC01410 expression and the mRNA and protein expressions of FOXM1 (Fig. 2H, I). These results indicated that LINC01410 regulated FOXM1 expression and miR-3619-5p potentially interacted with LINC01410 and FOXM1 in PTC.
LINC01410 sponged miR-3619-5p to regulate FOXM1 in PTC
Then, we tried to confirm the interaction of miR-3619-5p with LINC01410 and FOXM1 in PTC. The binding sites between LINC01410 and miR-3619-5p, the binding site between FOXM1 and miR-3619-5p, and the respective mutant sequences were presented in Figure 3A and B. Through luciferase reporter assays, we validated that overexpression of miR-3619-5p alleviated the luciferase activity of WT-LINC01410 and WT-FOXM1, but had no influence on that of Mut-LINC01410 and Mut-FOXM1 (Fig. 3C), confirming the interaction of miR-3619-5p with LINC01410 and FOXM1. Next, we evaluated whether FOXM1 was regulated by LINC01410/miR-3619-5p axis. LINC01410 overexpression in two PTC cell lines was confirmed by RT-qPCR results (Fig. 3D). Luciferase reporter assays validated that overexpression of LINC01410 reversed the reductive effect of miR-3619-5p mimic on the luciferase activity of WT-FOXM1 rather than Mut-FOXM1 (Fig. 3E). RT-qPCR and western blot analyses revealed that ectopic expression of LINC01410 restored the mRNA and protein expressions of FOXM1 reduced by miR-3619-5p mimic (Fig. 3F). Together, these data suggested that LINC01410 sponged miR-3619-5p to regulate FOXM1 in PTC.
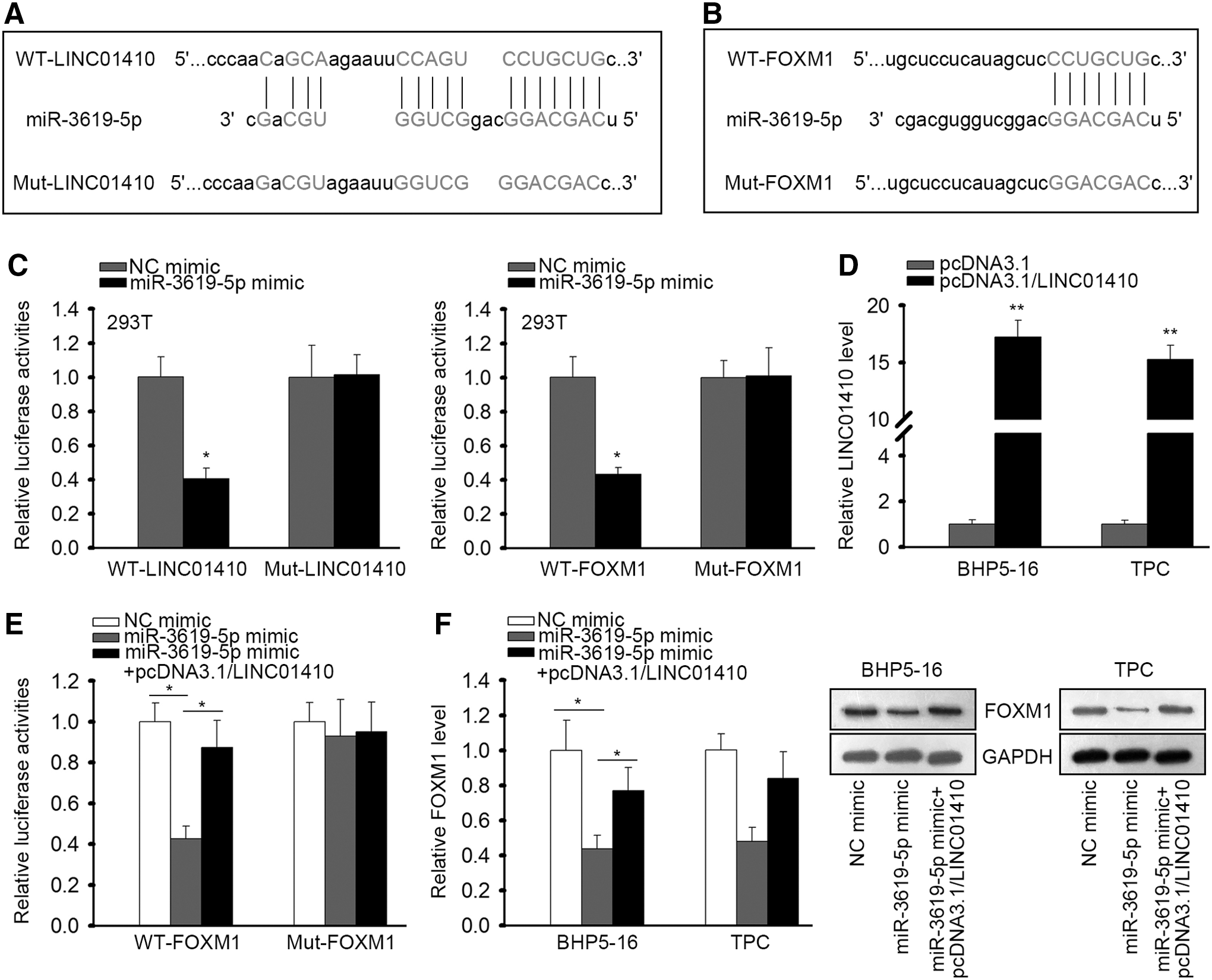
LINC01410 sponged miR-3619-5p to regulate FOXM1 in PTC.
FOXM1 transcriptionally induced LINC01410 expression in PTC
Additionally, since FOXM1 is a transcriptional factor that has been reported to activate the transcription of lncRNA in cancer, 28 we wondered whether FOXM1 could in turn regulate the expression of LINC01410 in PTC. Interestingly, we found through UCSC database that FOXM1 potentially bound to the promoter of LINC01410 (Fig. 4A). We then overexpressed FOXM1 to evaluate its influence on LINC01410 expression. The overexpression of FOXM1 was validated by RT-qPCR analysis in two PTC cell lines (Fig. 4B). Expectedly, overexpression of FOXM1 induced the expression of LINC01410 in PTC cells (Fig. 4C). Furthermore, ChIP assay revealed the enrichment of LINC01410 promoter in the precipitates of FOXM1 antibody, confirming the binding of FOXM1 on the promoter of LINC01410 (Fig. 4D). Finally, luciferase reporter experiments confirmed that overexpressing FOXM1 induced the activity of LINC01410 promoter (Fig. 4E). Collectively, these data implied that FOXM1 transcriptionally induced LINC01410 expression in PTC, which meant that LINC01410/miR-3619-5p/FOXM1 formed a positive feedback loop in PTC.

FOXM1 transcriptionally induced LINC01410 expression in PTC.
LINC01410 regulated cell proliferation and apoptosis through miR-3619-5p/FOXM1 axis in PTC
Eventually, we designed rescue assays to evaluate the role of LINC01410/miR-3619-5p/FOXM1 in PTC. MiR-3619-5p was inhibited in TPC cells as confirmed by RT-qPCR results (Fig. 5A). CCK-8 assay and EdU assay demonstrated that inhibiting miR-3619-5p or overexpressing FOXM1 could impair the inhibitory effect of LINC01410 knockdown on cell proliferation in PTC (Fig. 5B–C). Capase-3 activity induced by LINC01410 silence could be reversed by either miR-3619-5p inhibition or FOXM1 overexpression (Fig. 5D). In conclusion, these data suggested that LINC01410 regulated PTC cell proliferation and apoptosis through miR-3619-5p/FOXM1 axis.
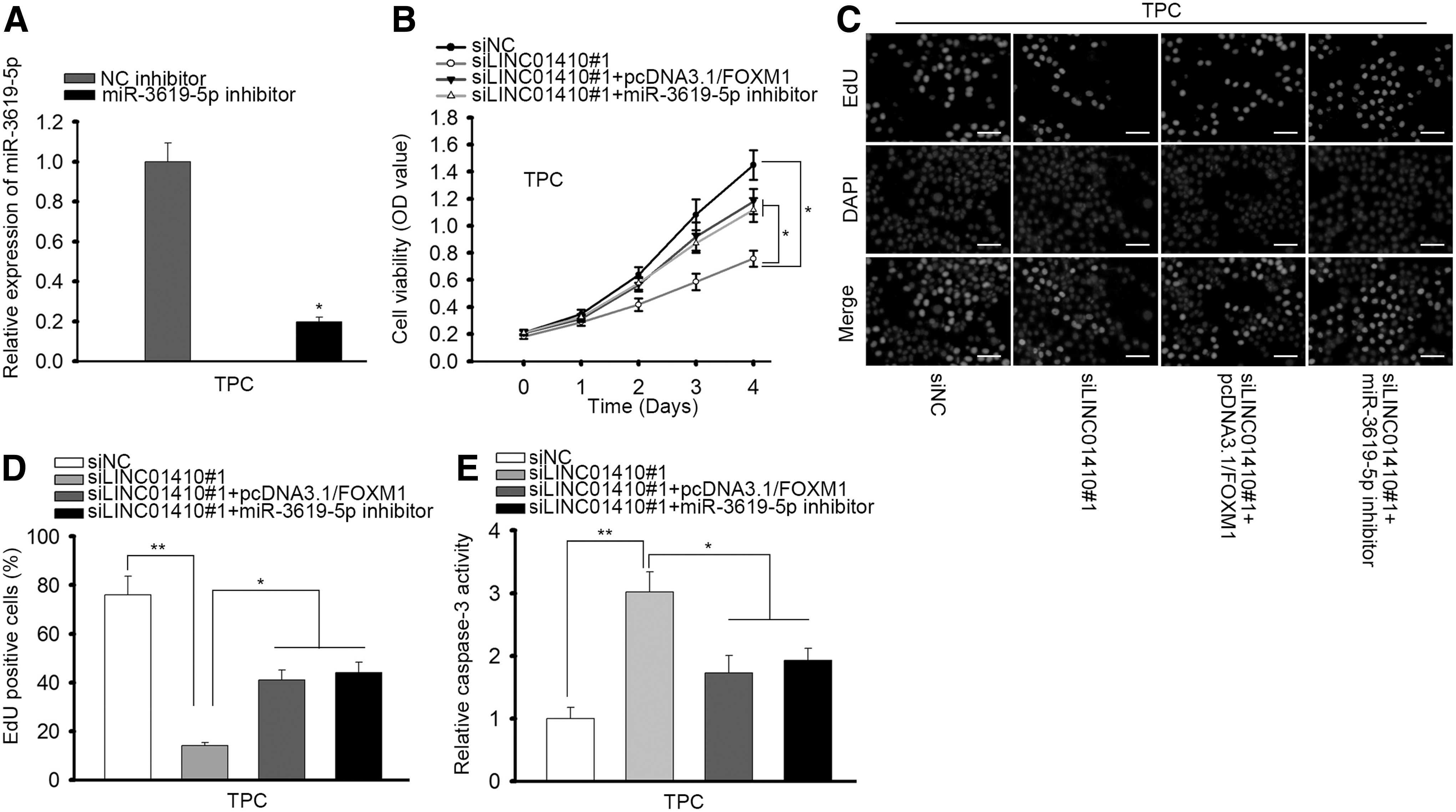
LINC01410 regulated cell proliferation and apoptosis through miR-3619-5p/FOXM1 axis in
Discussion
PTC is a major subtype of TC. 2 PTC patients at advanced stage tend to suffer from dismal prognosis, 6,7 which indicates an urgent need for the further understanding on the underlying mechanism of PTC progression to improve the treatment and survival of PTC patients.
More and more studies have pointed out that lncRNAs are important regulators of human cancer development, 10,11 including in PTC. 12,13 LINC01410 has been demonstrated in previous studies to exhibit carcinogenic function in gastric cancer and colon cancer, aggravating proliferation, angiogenesis, and metastasis. 14,15 Concordantly, we first identified LINC01410 to be upregulated in PTC cell lines, and validated through loss-of-function assays that silencing of LINC01410 attenuated cell proliferation and facilitated apoptosis in PTC cells, suggesting that LINC01410 exerted oncogenic functions in PTC. LncRNAs are known to usually exhibit their effects through regulating downstream target genes. 8 Previously, FOXM1, a transcription factor of forkhead box family, has been revealed to be highly expressed in PTC and regulated cell viability and metastasis. 27 Therefore, we speculated that LINC01410 might regulate FOXM1 in PTC. As expected, we confirmed the upregulation of FOXM1 in PTC cell lines and that LINC01410 positively regulated FOXM1 expression.
The mechanisms whereby lncRNAs regulated gene expressions could be different considering their cellular localization. 29 The ceRNA mechanism is one of the most common ways by which lncRNAs regulate downstream target genes in cytoplasm. 16,17 Formerly, LINC01410 has been reported to perform as ceRNA in gastric and colon cancers. 14,15 Herein, we validated that LINC01410 mainly expressed in cytoplasm, indicating the possibility that LINC01410 could also act as a ceRNA in PTC. 14,15 Through browsing lncBase and Starbase, we found four miRNAs potentially interacted with both LINC01410 and FOXM1. The induction of only miR-3619-5p expression by LINC01410 silencing indicated that miR-3619-5p might relate to LINC01410 in PTC. We then confirmed the downregulation of miR-3619-5p and the negative regulation of miR-3619-5p on FOXM1 expression, suggesting that LINC01410 might regulate FOXM1 through miR-3619-5p. Expectedly, we validated the interaction of miR-3619-5p with LINC01410 and FOXM1, representing the novel discovery of a ceRNA network LINC01410/miR-3619-5p/FOXM1 in PTC.
Former studies have shown that miR-3619-5p was a tumor suppressor several cancers. 19 –21 This study is the first one to suggest the tumor suppressive role of miR-3619-5p in PTC, which was concordant with the previous findings. Rescue assays illustrated that inhibiting miR-3619-5p or overexpressing FOXM1 could reverse the effect of LINC01410 knockdown on cell proliferation and apoptosis, indicating that LINC01410 regulated PTC cell proliferation and apoptosis through miR-3619-5p/FOXM1 axis.
Previous study has shown that FOXM1 could regulate the expression of lncRNA at transcription level in human cancers. 28 In this regard, we investigated whether FOXM1 regulated LINC01410 transcription in PTC cell lines. Surprisingly, we first found through UCSC that FOXM1 potentially bound to the promoter of LINC01410, indicating that FOXM1 might regulate the transactivation of LINC01410. Further mechanism research validated the interaction of FOXM1 with LINC01410 promoter and the transcriptional activation of LINC01410 by FOXM1, indicating that LINC01410/miR-3619-5p/FOXM1 formed a positive feedback loop in PTC. Moreover, FOXM1 had a reverse effect on siLINC01410-medieted PTC cell proliferation and apoptosis, suggesting that FOXM1 involved in PTC cell proliferation and apoptosis by cooperating with LINC01410. In collection, our study first found that LINC01410 formed a positive feedback loop with miR-3619-5p/FOXM1 to regulate proliferation and apoptosis in PTC, suggesting LINC01410 as a novel biological target for the improvement of treatment for PTC patients.
Conclusions
LINC01410 promoted PTC cell proliferation. LINC01410 acted as a ceRNA to upregulate FOXM1 by sponging miR-3619-5p in PTC. FOXM1 transcriptionally activated LINC01410 in PTC cell lines. LINC01410/miR-3619-5p/FOXM1 is a positive feedback loop in PTC cell proliferation and apoptosis.
Footnotes
Acknowledgment
Thank you for all participators involved in this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.