
Abstract
Cancer Biotherapy and Radiopharmaceuticals
officially retracts the paper entitled, “MiR-181a Promotes Apoptosis and Reduces Cisplatin Resistance by Inhibiting Osteopontin in Cervical Cancer Cells,” by Xiaofei Xu, Xiaofei Jiang, Liping Chen, Yu Zhao, Zhihua Huang, Huifang Zhou, and Mingqing Shi (Cancer Biother Radiopharm. 2019;34(9)559–565; doi: 10.1089/cbr.2019.2858) due to the discovery that the paper was submitted from a paper mill. This is a violation of the journal's standard protocols and is considered an infraction against the rigorous standards of scientific publishing.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves, and does not tolerate any improprieties.
Introduction
Cervical carcinoma (CC) is a common clinical female malignant tumor of the reproductive system, which is second only to breast cancer in female malignant tumors. 1 Recently, incidence of cervical cancer is increased and shows younger trend, posing a serious threat to the lives and health of female patients. 2,3
Osteopontin (OPN) is a calcium-binding phosphorylated glycoprotein, which was first discovered in bone matrix. Several studies reported abnormal expression of OPN in various tumor tissues. 4 –7 It was found that OPN plays an important role in the regulation of drug resistance in several cancer cells, such as liver cancer, 8 oral squamous cell carcinoma, 9 and prostate cancer. 10 However, there is still lack of report about its role in the drug resistance of cervical cancer. 8 –10
MicroRNA is a newly discovered noncoding single-stranded small RNA at 19–25 nucleotides in length. It is an important gene regulatory substance to degrade target mRNA or inhibit post-transcriptional translation through complementary binding to the 3′-untranslated region (3′-UTR) of target mRNA, thereby participating in the regulation of biological processes, such as cell growth, differentiation, apoptosis, and migration. 11,12 Multiple studies showed that abnormal expression of miR-181a can be detected in cervical cancer tumor tissues, and was associated with cervical cancer development, progression, and drug resistance. 13 –15
Bioinformatics analysis showed a target relationship between miR-181a and the 3′-UTR of OPN. In this study, the authors established a cervical cancer cisplatin (DDP) drug-resistant cell line to explore the role of miR-181a in the modulation of OPN expression and the proliferation, apoptosis, as well as DDP resistance of cervical cancer cells.
Materials and Methods
Main reagents and materials
Human normal cervical epithelial cells Ect1/E6E7 and cervical cancer cells Caski and Hela were purchased from Beijing Beina biological. HEK293T cells were purchased from the Chinese Academy of Sciences cell bank. RPMI 1640 medium, Opti-MEM, and fetal bovine serum (FBS) were purchased from Gibco. Trizol and Lipofectamine 2000 transfection reagent were purchased from Invitrogen. PrimeScript RT reagent Kit was purchased from Takara. MiR-181a mimic and miR-NC were purchased from RiboBio. Rabbit anti-human OPN polyclonal antibody was purchased from Abcam. Rabbit anti-human β-actin polyclonal antibody was purchased from Santa Cruz. Goat anti-rabbit IgG (H+L) secondary antibody was purchased from Sangon. CCK-8 cell proliferation activity detection reagent and Annexin V/PI apoptosis detection reagent were purchased from Shanghai Yu Sheng Biotechnology Co., Ltd. BeyoECL Plus and EdU cell proliferation assay reagent were purchased from Beyotime. Dual-Glo Luciferase Assay System kit was purchased from Promega. pMIR-REPORT Luciferase carrier was purchased from Changsha Youbao Bio.
Cell culture
Ect1/E6E7, Caski, and Hela cell lines were maintained in RPMI1640 medium containing 10% FBS and cultured at 37°C and 5% CO2 (FORMA, Thermo). The cells were passaged at 1:3–1:4. Cells in logarithmic phase were selected for further studies.
DDP drug-resistant cell line establishment
Caski or Hela cells were treated with DDP from 0.2 μg/mL for 2 weeks followed by treatment of increased concentration of DDP up to 3.2 μg/mL until the cells can stably grow in DDP. After that, cells can be stably passaged in the presence of DDP, which were named as DDP-resistant Caski/DDP or Hela/DDP cell line.
Caski Hela, Caski/DDP, or Hela/DDP cells were treated with different concentrations (0, 0.1, 1, 10, 100, and 1000 μg/mL) of DDP for 48 h followed by addition of CCK-8 reagent for the measurement of the absorbance value (A450). Inhibition rate = (1-A450 in drug group)/A450 in control × 100%. IC50 was calculated using SPSS software. Resistance index (RI) = IC50 of drug resistant cell/IC50 of parent cell.
Dual luciferase reporter gene assay
The polymerase chain reaction (PCR) product of the OPN 3′-UTR full-length fragment or mutant fragment was double digested and then ligated into the pMIR vector. After sequencing, the plasmid was designated as pMIR-OPN-WT and pMIR-OPN-MUT. The HEK293T cells were transfected with pMIR-OPN-WT (or pMIR-OPN-MUT) together with miR-181a mimic (or miR-NC) by Lipofectamine 2000 reagent. After incubation for 48 h, the relative luciferase activity was measured using Dual-Glo Luciferase Assay System kit according to the manual.
Cell transfection
Caski/DDP or Hela/DDP cells were divided into miR-NC group and miR-181a mimic group. A total of 10 μL of Lip2000, 50 nmoL miR-NC, and 50 nmoL miR-181a mimic were diluted with 100 μL serum-free Opti-MEM medium, and incubated for 5 min at room temperature, respectively. The mixture was added to the cell culture medium for 72 h. At last, the cells were collected for detection.
The cells were seeded in a six-well plate and treated with 3.2 μg/mL DDP for 48 h followed by measuring cell apoptosis on Beckmann FC500 MCL flow cytometry.
The cells were incubated with 10 μM EdU for 2 h. Next, the cells were treated with 3.2 μg/mL DDP for 48 h. Then, the cell proliferation was determined by EdU staining kit according to the manual.
quantitative real-time PCR
PrimeScript RT reagent Kit was used to reversely transcribe RNA to cDNA. The reverse transcription system contained oligdT Primer (50 μM) 0.5 μL, Random 6 mers (100 μM) 0.5 μL, PrimeScript RT Enzyme Mix 0.5 μL, RNA 1.0 μg, 5 × PrimeScript Buffer 2 μL, and RNase-Free H2O. The reverse transcription was performed at 37°C for 15 min and 85°C for 5 s. The PCR was performed on Bio-Rad CFX96, including SYBR Fast qPCR Mix 10.0 μL, forward primer (10 μM) 0.8 μL, reverse primer (10 μM) 0.8 μL, cDNA 2.0 μL, and RNase-free dH2O 6.4 μL. The PCR condition was designed as follows: 95°C for 10 min, followed by 40 cycles of 95°C for 10 s, 60°C for 20 s, and 72°C for 15 s. The primer sequences for miR-181a were as follows: Forwad-5′-GGGCAGCCTTAAGAGGA-3′ and Reverse-5′-CAGTGCGTGTCGTGGA-3′; U6: Forward-5′-GCTTCGGCAGCACATATACTAAAAT-3′ and Reverse-5′-CGCTTCACGAATTTGCGTGTCAT-3′; OPN: Forward-5′-ACTCCAATCGTCCCTACAGTCG-3′ and Reverse-5′-TGAGGTCCTCATCTGTGGCAT-3′; GAPDH: Forward-5′-AGGTCGGTGTGAACGGATTTG-3′ and Reverse-5′-TGTAGACCATGTAGTTGAGGTCA-3′.
Western blot
Total protein was isolated from cells using RIPA lysis buffer and quantified by BCA method. Then, 40 μg protein was loaded and separated by SDS-PAGE, followed by being transferred to polyvinylidene difluoride membrane at 300 mA for 100 min and subsequent blockage with 5% skim milk at room temperature. After that, primary antibody (OPN 1:2000 and β-actin 1:8000) was added into the membrane and incubated at 4°C overnight, followed by addition of horseradish peroxidase-conjugated secondary antibody (1:15,000) and incubation at room temperature for 60 min. Finally, BeyoECL Plus chemiluminiscence reagent was added into the membrane for visualization of the protein band.
Cell apoptosis analysis
The cells were collected and resuspended in 100 μL binding buffer followed by addition of 5 μL Annexin V-FITC and 5 μL PI, and incubation at room temperature for 15 min under dark. After that, cell apoptosis was measured by flow cytometry.
Cell proliferation detection
The cells were added with EdU solution at 10 μM in logarithmic phase and incubated for 2 h; cells were seeded for 48 h and digested by trypsin. After being fixed in paraformaldehyde, the cells were incubated in 100 μL TritonX-100 at room temperature and in 500 μL reaction fluid at room temperature for 30 min under dark, followed by analysis of cell proliferation by flow cytometry.
Clone formation experiment
Forty microliters of cell suspension (1 × 104/mL) was uniformly inoculated into 60 mm culture dishes, and each group was set up with three parallel dishes. After the cells were attached, the cells were transfected into miR-181a and NC groups, and cultured for 2 to 3 weeks. The culture solution was discarded, and the methanol solution was fixed for 20 min. After staining with crystal violet, cell colonies (≥50 cells) were counted, each group was counted five times, and the average was calculated.
Statistical analysis
SPSS 18.0 software was used for processing data. Measurement data were displayed as mean ± standard deviation and compared by unpaired student t-test or one-way analysis of variance. p < 0.05 was considered a significant difference.
Results
The relationship between miR-181a and OPN mRNA
Bioinformatics analysis revealed a complementary binding site of 3′-UTR of OPN mRNA in miR-181a (Fig. 1A). Dual luciferase reporter gene assay exhibited that miR-181a mimic transfection significantly decreased the relative luciferase activity in HEK293T cells, which were transfected with pMIR-OPN-WT, but not with pMIR-OPN-MUT, confirming that OPN was the target gene of miR-181a (Fig. 1B).

The targeted regulatory relationship between miR-181a and OPN.
Drug-resistant cervical carcinoma cells exhibited strong proliferation and drug resistance
CCK-8 assay showed that the proliferative activities of parental Caski or Hela cells were significantly lower than those of Caski/DDP cells under the same dose treatment of DDP (cisplatin) (Fig. 2A). The IC50 was 2.91 ± 0.24 μg/mL for Caski cells and 2.62 ± 0.41 μg/mL for Hela cells, while it was 34.57 ± 2.89 μg/mL in the drug-resistant Caski/DDP cells and 30.64 ± 0.37 μg/mL in the drug-resistant Hela/DDP cells, with the RI being 11.88 for Caski/DDP cells and 11.69 for Hela/DDP cells (Fig. 2B). Consistently, miR-181 significantly inhibited the clone formation (Fig. 2C).
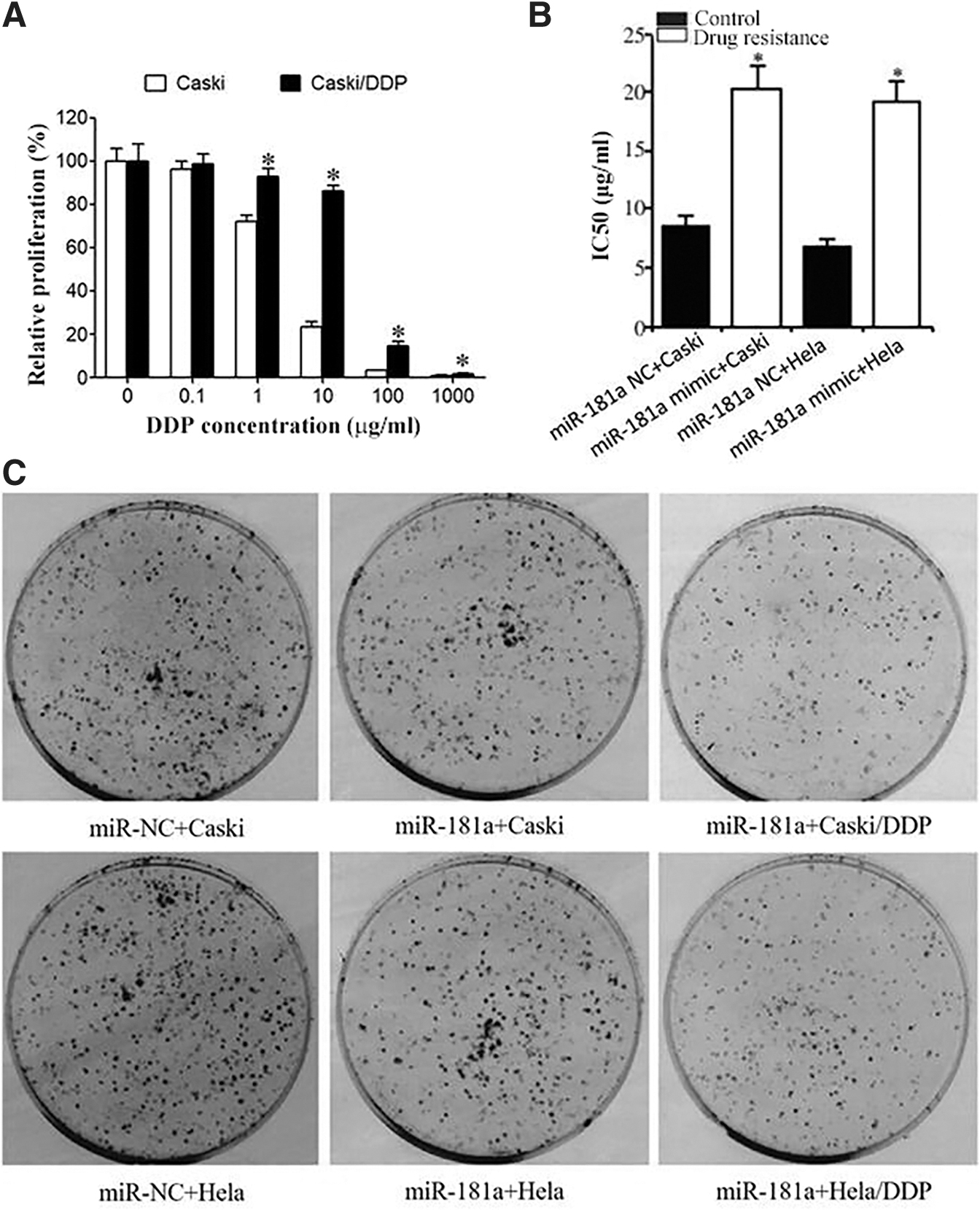
CCK-8 detection of Caski, Hela, Caski/DDP, and Hela/DDP cell viabilities.
MiR-181a reduced, while OPN upregulated in drug-resistant cells
quantitative real-time PCR (qRT-PCR) demonstrated that compared with human normal Ect1/E6E7 cells, miR-181 expression was significantly decreased in Caski or Hela cells, and its level was significantly lower in Caski/DDP or Hela/DDP cells compared with parental cells (Fig. 3A). qRT-PCR revealed that the OPN mRNA expression in Caski or Hela cells was significantly higher compared with Ect1/E6E7 cells, and it was apparently higher in Caski/DDP or Hela/DDP cells compared with the parental cells (Fig. 3B). Western blot exhibited that compared with Ect1/E6E7 cells, OPN protein expression in Caski or Hela cells was significantly upregulated, and its/level in Caski/DDP or Hela/DDP cells significantly enhanced (Fig. 3C).
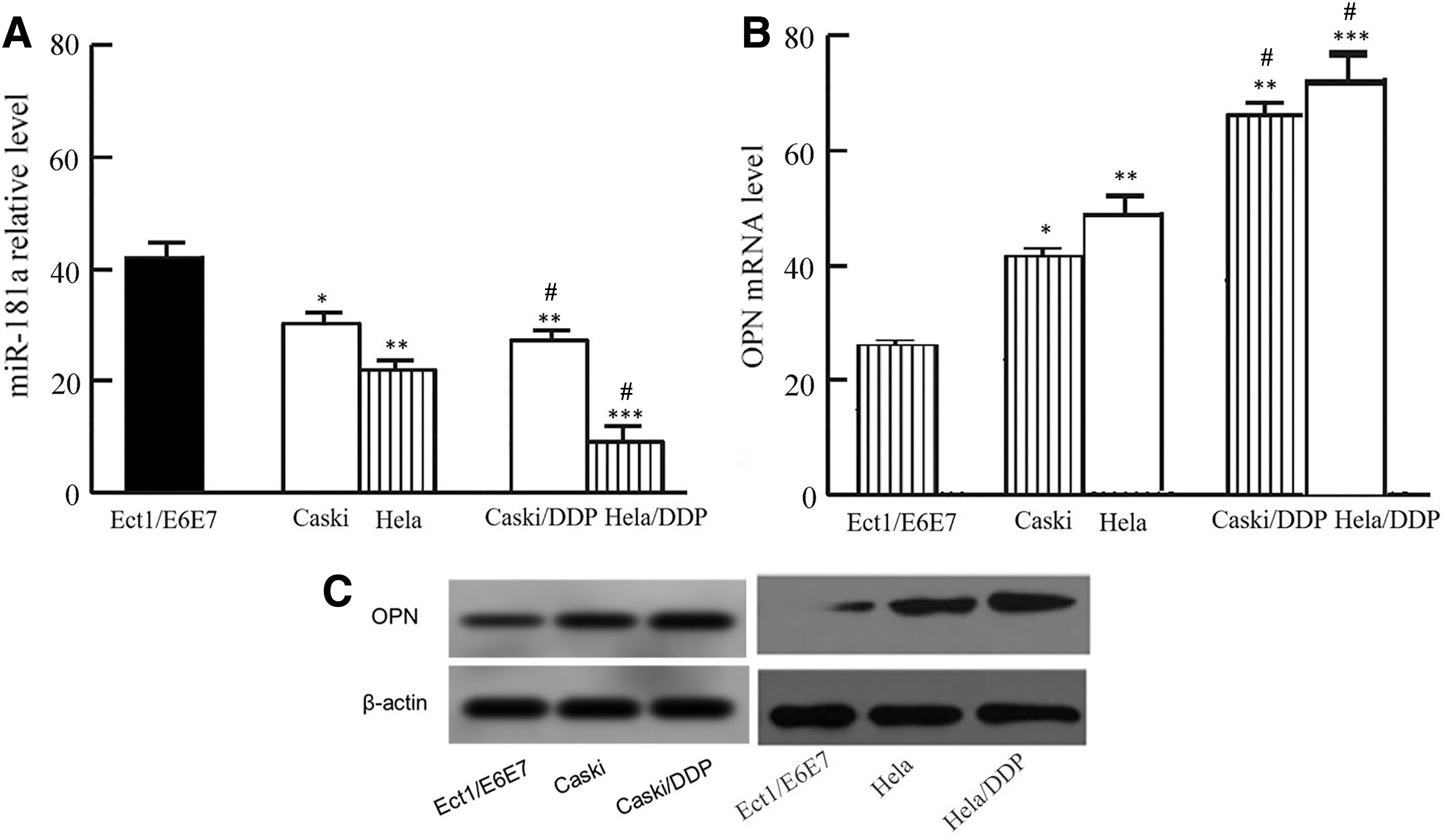
MiR-181a reduced, while OPN upregulated in drug-resistant cells.
MiR-181a overexpression promoted CC cell apoptosis and reduced DDP resistance
qRT-PCR showed that miR-181a mimic transfection significantly upregulated miR-181a expression and declined OPN mRNA level in Caski/DDP or Hela/DDP cells compared with miR-NC group (Fig. 4A, B). Western blot demonstrated that miR-181a mimic transfection significantly elevated OPN protein level in Caski/DDP or Hela/DDP cells (Fig. 4C). Flow cytometry revealed that transfection of miR-181a mimic significantly enhanced cell apoptosis, while inhibiting cell proliferation in Caski/DDP or Hela/DDP cells (Fig. 4D, E).
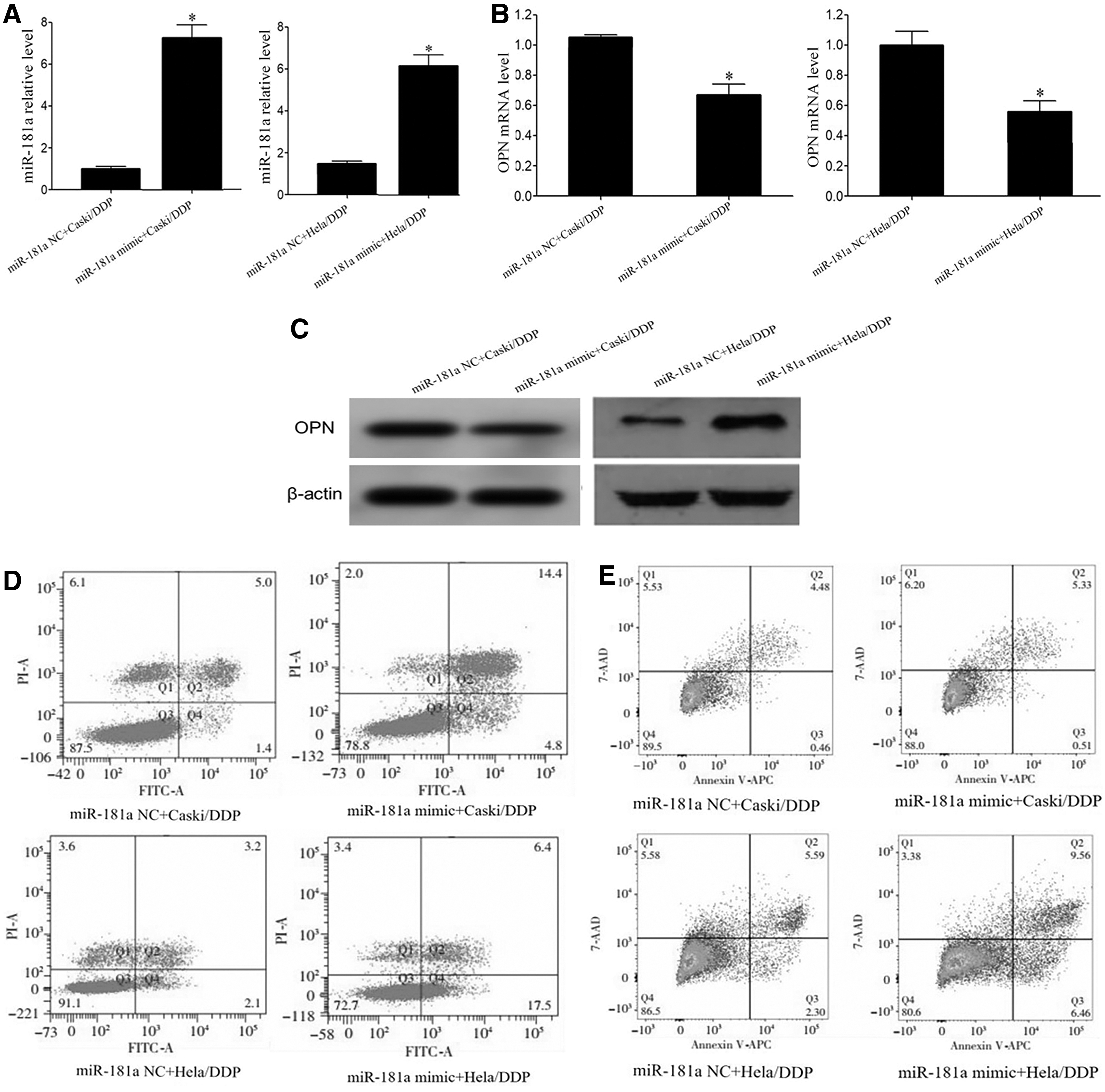
MiR-181a overexpression promoted CC cell apoptosis and reduced DDP resistance.
Discussion
Human papillomavirus (HPV) infection is an important virulence factor in the pathogenesis of cervical cancer. In recent years, with the increase of HPV infection, the incidence of cervical cancer is getting higher and higher. 16,17 DDP is a common drug in neoadjuvant chemotherapy for cervical cancer. 18 –20 However, several patients exhibited reduced sensitivity to DDP chemotherapy and were prone to distant metastasis and recurrence even after chemotherapy. 21 –23 Therefore, it is of great significance to find the abnormal signal molecules and explore the mechanism of cervical cancer resistance, which is helpful to improve cervical cancer chemotherapy efficacy and prognosis.
The human OPN gene is 3143 bp in length and located on chromosome 4q13 containing 7 exons. The OPN gene encodes a protein containing 300 amino acids. 24,25 OPN is a secreted calcium-binding phosphorylated glycoprotein containing an RGD (argin-glyco-aspartate) integrin binding region first found in bone matrix. OPN is widely expressed in various tissues, such as T cells, epithelial cells, bone cells, macrophages, and endothelial cells. 26 –28 OPN can promote tumorigenesis and progression by inducing tumor angiogenesis, facilitating extracellular matrix degradation, accelerating tumor metastasis and invasion, and inhibiting tumor cell apoptosis. 7,24,25,29 Abnormal expression of OPN was associated with the occurrence, progression, and metastasis of various tumors, such as gastric cancer, 4 colorectal cancer, 5 lung cancer, 6 and breast cancer. 7 OPN has been shown to play an important role in the regulation of drug resistance of several cancer cells, such as liver cancer, 8 oral squamous cell carcinoma, 9 and prostate cancer. 10 However, there is still lack of report about the impact of OPN in cervical cancer drug resistance. The abnormal expression of miR-181a can regulate various biological effects, such as cell proliferation and apoptosis; thus it is related to tumorigenesis, progression, metastasis, and drug resistance of multiple cancers, including thyroid cancer, 30 gastric cancer, 31 lung cancer, 32 and breast cancer. 33 Numerous investigations revealed that abnormal expression of miR-181a is also detected in cervical cancer and associated with development, progression, and drug resistance. 13 –15 Bioinformatics analysis demonstrated a target relationship between miR-181a and the 3′-UTR of OPN. In this study, the authors established a cervical cancer DDP drug-resistant cell line to explore the role of miR-181a in the regulation of OPN expression and the proliferation, apoptosis, as well as DDP resistance of cervical cancer cells.
In this study, dual luciferase reporter gene assay exhibited that miR-181a mimic transfection significantly decreased the relative luciferase activity in HEK293T cells, which were transfected with pMIR-OPN-WT, confirming that OPN was the target gene of miR-181a. CCK-8 assay showed that the proliferative activities of parental Caski cells were significantly lower than those of Caski/DDP cells under the treatment of same dose of DDP. The IC50 of Caski/DDP cells was significantly higher compared with Caski cells, indicating successful DDP-resistant Caski/DDP cell line establishment. Gene and protein detections demonstrated that compared with normal cervical epithelial cells, miR-181a expression was significantly decreased, while OPN level was apparently enhanced in cervical cancer cells. Compared with parental Caski cells, MiR-181a was significantly reduced, while OPN mRNA and protein were significantly elevated in Caski/DDP cells. It indicated that decreased miR-181a played a role in upregulating OPN, and the abnormalities of miR-181 and OPN were related to drug resistance in cervical cancer cells. Luo and Qiu 14 reported that miR-181a was significantly declined, while the target gene GRP78 was significantly strengthened in cervical cancer cells, and decreased expression of miR-181a was associated with cervical cancer tumor growth and oxaliplatin resistance. This study exhibited that miR-181a decreased in cervical cancer cells and was related to the drug resistance of cervical cancer, which was similar to Luo and Qiu. 14 In the study of the relationship between OPN and cervical cancer, Leung et al. 34 proved that the serum levels of OPN in cervical cancer patients were significantly higher than those in healthy controls. Song et al. 35 found that the expression of OPN in tumor tissues of patients with cervical cancer was significantly increased, and the increase of OPN expression was positively correlated with the degree of tumor invasion. It showed that abnormal elevation of OPN was associated with cervical cancer, which was similar to these results.
At present, there are few studies on the relationship between miR-181a and drug resistance of cervical cancer cells. Therefore, this study explored whether miR-181a affects DDP resistance in cervical cancer cells by regulating OPN expression. It was observed that transfection of miR-181a mimic significantly reduced the expression of OPN in drug-resistant Caski/DDP cells, which significantly enhanced cell apoptosis, restrained cell proliferative ability, and alleviated DDP drug resistance. Luo and Qiu 14 revealed that elevated expression of miR-181a inhibited cervical cancer cell growth and reduced the oxaliplatin resistance by downregulating GRP78, while increasing the expression of target gene GRP78 promoted cervical cancer cell proliferation and enhanced oxaliplatin drug resistance. This study explored the targeted relationship between miR-181a and OPN, revealing that miR-181a plays a role in targeted inhibiting OPN expression, promoting cervical cancer cell apoptosis, and reducing DDP resistance. However, whether miR-181a regulating OPN is related to the drug resistance of cervical cancer patients is still unclear and requires further investigations. In addition, large cohort clinical studies are required to confirm the findings from this study.
Conclusion
The decrease of miR-181a expression and the upregulation of OPN expression are related to the DDP resistance of cervical cancer cells. Enhancement of miR-181a can inhibit the expression of OPN, induce cell apoptosis cells, restrain cell proliferation, and reduce DDP resistance in cervical cancer cells.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.