
Abstract
Background:
To investigate the effects of curcumin on miR-34a and proliferation of gastric cancer cells.
Materials and Methods:
Human gastric cancer cell line SGC-7901 was divided into control, curcumin, miR-34a agomir (miR-34a), miR-34a agomir negative control, and curcumin combined miR-34a antagomir (combine) groups. 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide assay, scratch damage test, and transwell assay were used to detect cell proliferation, migration, and invasion. The cell apoptosis and cell cycle were detected by flow cytometry. Western blot was used to detect the expression of B-cell lymphoma-2 (Bcl-2), cyclin-dependent kinase 4 (CDK4), and cyclin D1. Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling was used to detect apoptosis of tumors and Western blot was used to detect the expression of Bcl-2, CDK4, and cyclin D1 in tumors.
Results:
The results showed that curcumin markedly increased the content of miR-34a microRNA (mRNA) in SGC-7901 cells, inhibited proliferation, migration, and invasion of SGC-7901 cells, when compared to control group (p < 0.05). Compared with control group, curcumin significantly inhibited cell cycle progression in G0/G1-S phase, increased the cell number of G0/G1 phase, and downregulated the Bcl-2, CDK4, and cyclin D1 protein expression in cells and tissues (p < 0.05). After transfection of miR-34a agomir or antagomir into cells it was found that miR-34a agomir and curcumin had similar effects on resisting malignant biological behavior. Curcumin combined with miR-34a antagomir could weaken or reverse the above results.
Conclusions:
Curcumin could inhibit the proliferation and induce apoptosis of SGC-7901 cells. Its mechanism might be related to the miR-34a expression in cells, thus affecting the expression of Bcl-2, CDK4, and cyclin D1.
Introduction
In recent years, gastric cancer has become one of the most common malignant tumors of digestive tract in China. 1 The occurrence of gastric cancer is the result of stimulation by multiple factors, such as diet, infection, hormones, heredity, and so on. 2 In particular, the blocked apoptosis of heteromorphic cells is closely related to the occurrence of gastric cancer. Because of the lack of clinical manifestations, most patients with early gastric cancer have reached advanced stage when they are diagnosed. Radical gastrectomy is still the first choice for treatment at present. 3 However, chemotherapy can bring intolerable pain to patients, especially bone marrow suppression, gastrointestinal reactions, liver and kidney function damage, and other toxic side effects. 4 Therefore, searching for new therapeutic drugs has become an urgent problem for medical workers.
Curcumin, a natural polyphenol pigment compound, is widely found in Curcuma, Calamus, and other traditional Chinese medicines. 5 Curcumin has been used for hundreds of years in southeast Asia and India. It is widely used in the treatment of digestive tract diseases, respiratory diseases, rheumatism, immune diseases, diabetes, and other diseases. 6,7 It has been widely recognized in anti-immunity, antiangiogenesis, anti-inflammatory, lipid-lowering, antioxidation, and anticoagulation. 8 Recent studies have shown that curcumin alone or in combination with other anticancer drugs could effectively induce apoptosis in cancer cells, such as melanoma, mantle cell lymphoma, liver cancer, prostate cancer, ovarian cancer, and pancreatic cancer. 9
mRNA is a single-stranded noncoding RNA that has been widely concerned and used in recent years, which can be used as a potential drug target for the treatment of human diseases. 10 Previous studies have shown that miR-34a has biological activities such as regulating adipocyte differentiation, inhibiting cancer growth, and regulating inflammation. 11,12 In addition, previous studies have proved that miR-34a played an important role in gastric cancer. 11 Curcumin could significantly regulate the expression of miR-34a in colorectal cancer and breast cancer. 12 However, as far as we know, no studies have reported the effect of curcumin on miR-34a in gastric cancer.
Materials and Methods
Cell culture
Human gastric cancer cell line SGC-7901 (BNCC100674) was cultured in RPMI 1640 (Gibco, Rockville, MD) medium containing 10% fetal bovine serum (FBS) (Sigma-Aldrich, St. Louis, MO) in a humidified cell incubator (Thermo Fisher, Waltham) of 5% CO2 at 37°C for static culture. The fourth generation cells were selected for experiment. About 80% of the cells were collected for detection.
Cells were divided into five groups: Control (phosphate-buffered saline [PBS] treatment of gastric cancer cells; curcumin (50 μM curcumin interfered with gastric cancer cells, doses obtained from Tian et al. 8 ); miR-34a agomir (transfection of miR-34a agomir [miR41275141400-4-5]); miR-34a agomir negative control (NC) group (miR-34a agomir NC reagent [miR4N0000001-4-5]); and curcumin combined miR-34a antagomir groups (combine) (50 μM curcumin combined with miR-34a antagomir treatment [miR30000255-4-5]).
SGC-7901 cells were digested and passaged by 0.25% trypsin (Invitrogen, Carlsbad, CA). About 2 × 105 growth cells were inoculated into six-well plates. After 24 h, cell growth was observed by inverted microscope. When cell density reached 30%–50%, Lipofectamine™ 2000 transfection reagent (Invitrogen) was used according to the instructions. miR-34a agomir, miR-34a agomir NC reagent, and miR-34a antagomir reagent (purchased from Guangzhou Ruibo Biotechnology Co., Ltd., China) were transfected into SGC-7901 cell with Lipofectamine 2000 respectively to construct the overexpressed and silenced miR-34a cell lines. Curcumin (CAS 458-37-7; Sigma-Aldrich) was prepared with dimethyl sulfoxide (DMSO) into 1 M solution, frozen at −20°C for storage. Curcumin was thawed and preserved at 4°C. The concentration of 50 μM curcumin was prepared with RPMI-1640 medium containing 5% FBS and 1% penicillin-streptomycin.
Real-time fluorescence quantitative polymerase chain reaction
Total RNA were extracted by total RNA extraction kit (A27828, MagMAX™ mirVana™ Total RNA Isolation Kit; Thermo Fisher Scientific), and then transcribed into cDNA. Real-time polymerase chain reaction (RT-PCR) reaction system was constructed using 2 μL cDNA as template. The reaction was carried out under the following conditions: 95°C for 10 min; 95°C for 15 s; 60°C for 60 s, and 40 cycles were amplified. The relative expression of miR-34a was calculated by using U6 gene as internal reference and 2−▵▵Ct method. miR-34a primer was designed (Shanghai Biotechnology Service Co., Ltd.) as followed: F: 5′-ACACTCCAGCTGGGTGGCAGTGTCTTAG-3′; R: 5′-CTCAACTGGTGTCGTG-GAGTCG-3′. U6 was designed as followed: F: 5′-GATTATCGGACCATTCCACTG-3′; R: 5′-GATCTGTTCCATGACTGTG-3′.
MTT assay for cell proliferation
The cells of each group were digested by 0.25% trypsin-ethylenediamine tetraacetic acid (EDTA), and the cell concentration was adjusted to 1 × 104/mL. The cells were inoculated on 96-well flat-bottomed culture plate, incubated in incubator of 5% CO2 and 37°C. 20 μL 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide (MTT) (5 mg/mL) was added after 24, 48, 72, and 96 h of incubation, respectively. The cells were cultured in incubator of 37°C. After 4 h, the supernatant was discarded carefully and 200 μL DMSO (Sigma-Aldrich) was added. The absorbance optical density (OD) value of each hole was measured at 490 nm of the wavelength of Bio-Rad Laboratories.
Scratch test for cell migration
Cells were digested with 0.25% trypsin-EDTA and centrifuged at 1000 rpm for 5 min. Cells were collected and cell density was adjusted to 3 × 105 cells/mL. It is advisable to add 1 mL cell suspension into each hole of the six-well plate and confluence 100% within 24 h. The cells were scribed in sterile environments with a 10 μL tips and the suspended cells were removed by PBS washing twice. Serum-free medium was added slowly and cultured in 5% CO2 incubator at 37°C. Photographs were collected at 0 and 24 h respectively.
Transwell assay for cell invasion
Cells were collected with 0.25% trypsin-EDTA in sterile environment. The cells were removed and suspended in serum-free DMEM (Gibco) medium. The cell density was 2 × 105 cells/mL. A mixed solution of 50 μL matrix gel was added to the upper chamber in advance, and solidified in 37°C incubator for 30 min. About 600 μL complete medium containing 10% FBS was added to the 24-well plate. Then, 100 μL cell suspension was taken from the upper chamber and cultured in 5% CO2 incubator at 37°C. Membranes were collected after 24 h, fixed with methanol for 30 min, stained with 0.5% crystal violet for 20 min, and observed under optical microscope (Olympus, Japan). Nine visual fields were randomly selected for photography, and the number of invasive cells on the membrane was calculated.
Flow cytometry to detect apoptosis and cell cycle
Cells were collected by centrifugation (1000 rpm for 5 min) after digestion with 0.25% trypsin-EDTA. Cells were washed twice with sterile PBS solution precooled at 4°C. The total number of cells was about 1 × 106 cells/mL. Cells were resuspended with 250 μL 1 × binding buffer. Then, 5 μL FITC-labeled ANNEXIN-V were added to 195 μL of cell suspension. After gently mixing, the cells were placed for 3 min. Then, 10 μL Propidium Iodid (20 μg/mL) was mixed in cells. The incubation time was 10 min at room temperature. Added 400 μL 1 × binding buffer, gently mixed, and then did flow cytometry (Gallios; Beckman Coulter, Inc., Brea, CA) for detection. The results were analyzed by Cell Quest software (BD Biosicences, San Diego, CA).
Cells were inoculated into six-well plates and were 80% to 90% confluent, then they were digested with 0.25% trypsin-EDTA and centrifuged at 1000 rpm for 5 min. Cells were collected and washed twice with cold 1 × PBS. After centrifugation (1000 rpm for 6 min), 70% precooled ethanol (−20°C) was fixed for 12 h (4°C). Then, centrifugated cells with 1500 rpm for 5 min and washed with 1 × PBS. Cells were resuspended with 400 μL PI/RNA enzyme staining buffer (BD Pharmingen, San Diego, CA). Cells were detected by flow cytometry after 30 min exposure to light at 4°C.
Western blot to detect protein expression
The expression levels of B cell lymphoma-2 (Bcl-2), cyclin-dependent kinase 4 (CDK4), and cyclin D1 were detected by Western blot. Cells in each group were collected and the total protein was extracted according to the instructions of the kit. The concentration of protein samples was detected by BCA protein quantitative kit (23225, Pierce BCA Protein Assay Kit; Thermo Fisher Science). Protein was separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) electrophoresis (Mini Protein 3; Bio Rad, Hercules, CA) and transferred to polyvinylidene fluoride membrane (Millipore, MA), then blooked with 5% evaporated milk for 1 h. 5% bovine serum albumin was added to dilute the first antibody of each protein: Rabbit anti human Bcl-2 antibody (1:1000, ab196495; Abcam, UK); anti-CDK4 antibody (1:1000, ab137675; Abcam); anti-cyclin D1 antibody (1:2000, ab226977; Abcam); and β-actin polyclonal antibodies (1:1000, ab8227; Abcam) and then incubated with goat anti-rabbit IgG (1:2000, ab6721; Abcam) for 1 h. Emitter coupled logic chemiluminescence was detected by image J software (National Institute of Health [NIH]) for grayscale scanning and quantification. The protein expression level was standardized according to β-actin.
Construction and grouping of xenograft tumor model in nude mice
Thirty-six BALB/c male nude mice (7–8 weeks old, 20 ± 2 g) were selected (purchased from Jinan Pengyue Laboratory Animal Breeding Co., Ltd., license No. SCXK (Lu) 2014-0007). All animals were housed in a controlled environment at 22°C–24°C and relative humidity of 50%–60% with free access to food and water. Animal experiments follow the NIH guidelines (NIH Pub. No. 85-23, revised 1996) and have been approved by the Animal Protection and Use Committee of Weihai Central Hospital.
Cells in logarithmic phase were collected from each group and digested into cell suspension by trypsin and the concentration was adjusted to 1.0 × 107 cells/mL. Thirty-six BALB/c male nude mice (n = 6) were selected and randomly selected 6 mice as normal control. The remaining nude mice were inoculated with 200 μL stable transfected cell suspension in the subcutaneous tissue of the right lower extremity to construct the nude mice model of transplanted tumors.
The survival condition of nude mice was observed every day, and the growth of tumors was observed every week, and the volume of tumors was measured by vernier caliper. The growth curve of tumors was drawn within 4 weeks with the volume: V = a*b 2 /2 (a: tumor length; b: tumor short diameter). After 4 weeks, nude mice were anesthetized by intraperitoneal injection of pentobarbital sodium (70 mg/kg). Cervical vertebrae dislocated after anesthesia, and tumors were taken for weighing. Partial tumor tissues were immersed and immobilized in 4% paraformaldehyde solution for subsequent terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labeling (TUNEL) experiments, a method for detecting apoptosis in cells or tissues. Some tumor tissues were put into liquid nitrogen and then transferred to −80°C refrigerator for Western blot detection.
TUNEL detection of apoptotic level of tumor cells
Tumor tissues fixed in 4% paraformaldehyde solution were routinely embedded in paraffin and coronal section (4 μm) (paraffin embedding machine; paraffin section machine/Leica company, Germany). Five slices of each specimen were prepared. After routine xylene dewaxing and gradient ethanol dehydration, cell apoptosis detection kit (batch number: ZK-8005; Beijing Zhongshanjinqiao Biotechnology, China) was used. TUNEL assay was used to quantitatively detect apoptotic tumor cells. Five visual fields were randomly selected under 400 × optical microscope. The nuclei of normal cells were blue and the apoptotic positive cells were brown. The numbers of positive cells were counted under the microscope.
Statistical methods
SPSS19.0 statistical software was used to analyze the statistical data. The results of data analysis were expressed as mean ± standard deviation. One-way analysis of variance (ANOVA) was used for data analysis among multiple groups, and Fisher's least significant difference test was used for subsequent analysis. The difference was statistically significant with p < 0.05.
Results
Curcumin increased miR-34a mRNA levels and decreased cell proliferation ability
The content of miR-34a mRNA in cells of each group was detected by RT-quantitative PCR (qRT-PCR). As shown in Figure 1A. Compared to the control group, the expression of miR-34a mRNA increased significantly in curcumin, miR-34a, and combine groups (p < 0.05). Compared to curcumin and miR-34a groups, the content of miR-34a mRNA in combine group significantly decreased (p < 0.05). MTT assay was used to detect the proliferation activity of cells in each group after 24, 48, 72 and 96 h respectively. As shown in Figure 1B, compared with control group, the cell proliferation ability in curcumin, miR-34a, and combine groups was decreased significantly (p < 0.05). Compared with curcumin and miR-34a groups, the proliferation level of cells in combine group was reversely increased (p < 0.05).

Curcumin increased miR-34a mRNA levels and decreased cell proliferation ability.
Administration of curcumin or miR-34a decreased cells migration and invasion
Cell scratch test and transwell test were used to detect the migration and invasion ability of SGC-7901 cells. As shown in Figure 2, compared with control group, the wound healing rate and the number of penetrating cells were obviously decreased in curcumin, miR-34a, and combine groups (p < 0.05). Compared with curcumin and miR-34a groups, the wound healing rate and the number of penetrating cells in combine group were reversely increased (p < 0.05).
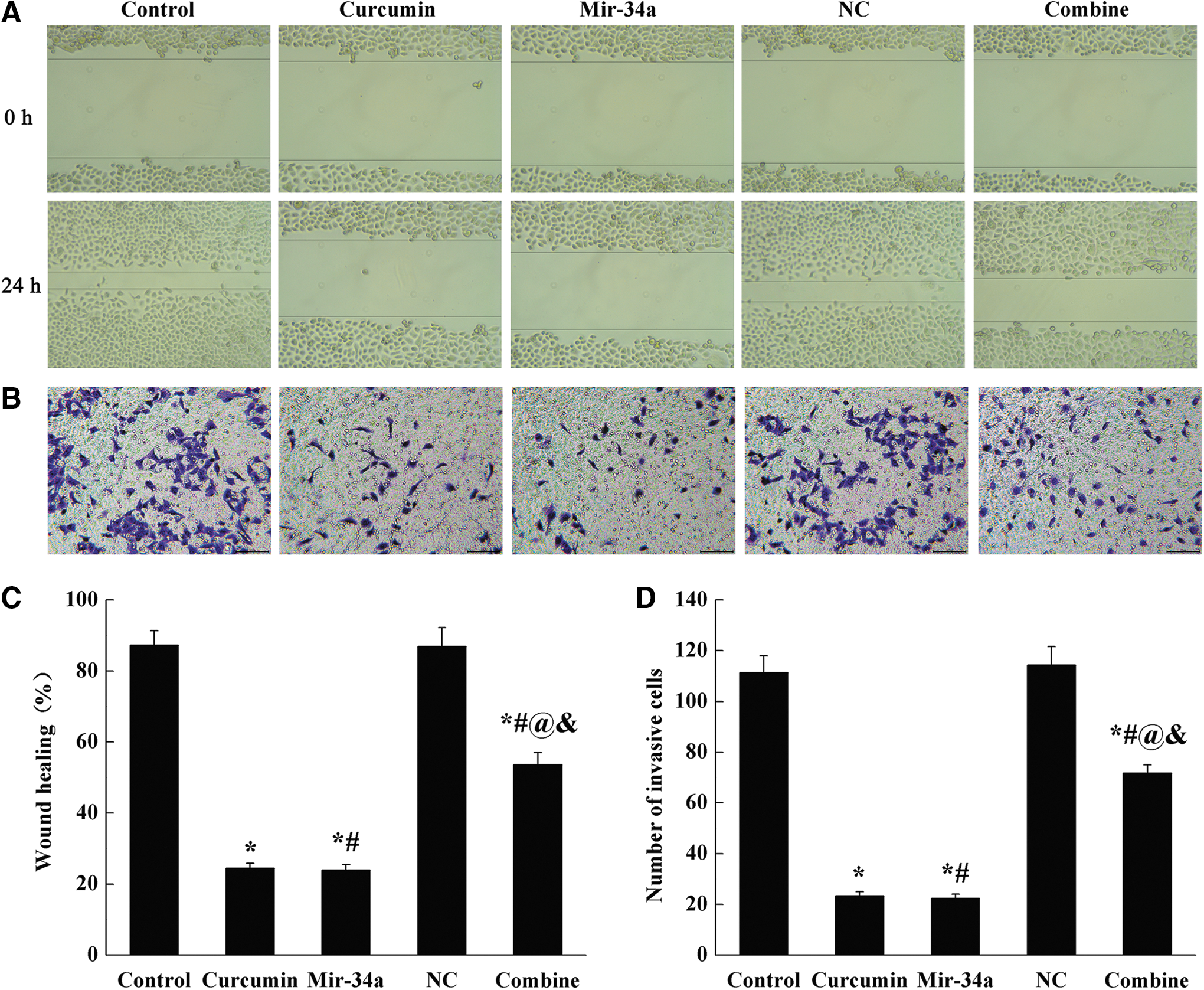
Administration of curcumin or miR-34a decreased cell migration and invasion.
Administration of curcumin or miR-34a promoted cell apoptosis
The apoptosis of SGC-7901 cells was detected by flow cytometry (Fig. 3A, C). Compared with control group, the apoptotic cells in curcumin, miR-34a and combine groups were all increased significantly (p < 0.05), and there was no significant difference between curcumin and miR-34a groups. Compared with curcumin and miR-34a groups, the apoptotic cells in combine group decreased significantly (p < 0.05).
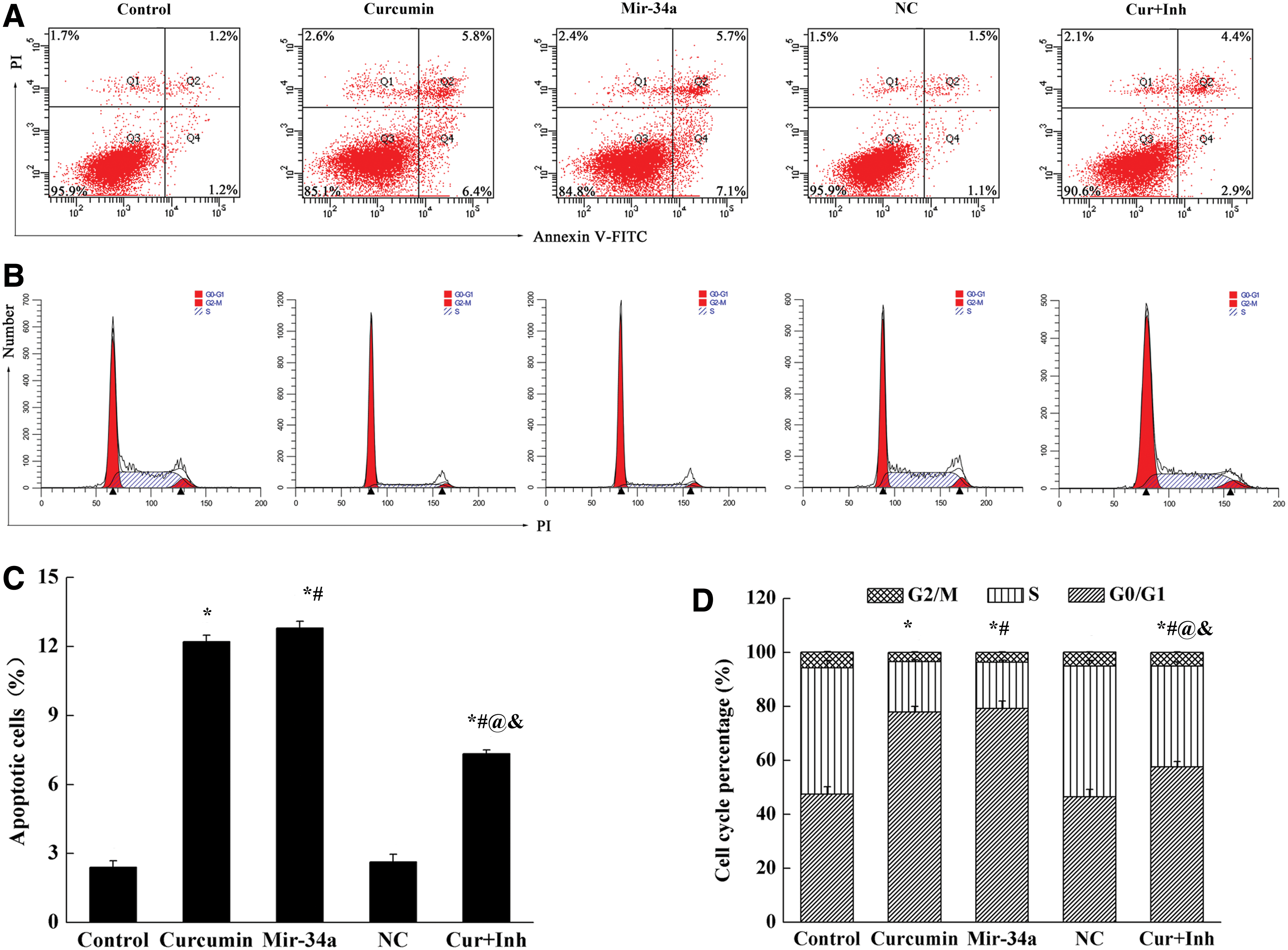
Administration of curcumin or miR-34a promoted cell apoptosis.
Further, flow cytometry was used to detect cell cycle (Fig. 3B, D). Compared with control group, the number of G0/G1 phase cells increased significantly in curcumin, miR-34a, and combine groups, while the number of S phase cells decreased (p < 0.05). Compared with curcumin and miR-34a groups, the numbers of G0/G1 phase cells in combine group were significantly lower (p < 0.05).
Administration of curcumin or miR-34a decreased Bcl-2, CDK4, and cyclin D1 expression
As shown in Figure 4, the expression levels of Bcl-2, CDK4, and cyclin D1 were detected by Western blot. Compared with control group, the expression levels of Bcl-2, CDK4, and cyclin D1 in curcumin, miR-34a, and combine groups were significantly decreased (p < 0.05), and there was no significant difference between curcumin and miR-34a groups. Compared with curcumin and miR-34a groups, the expression levels of these three proteins in combine group were reversely increased (p < 0.05).
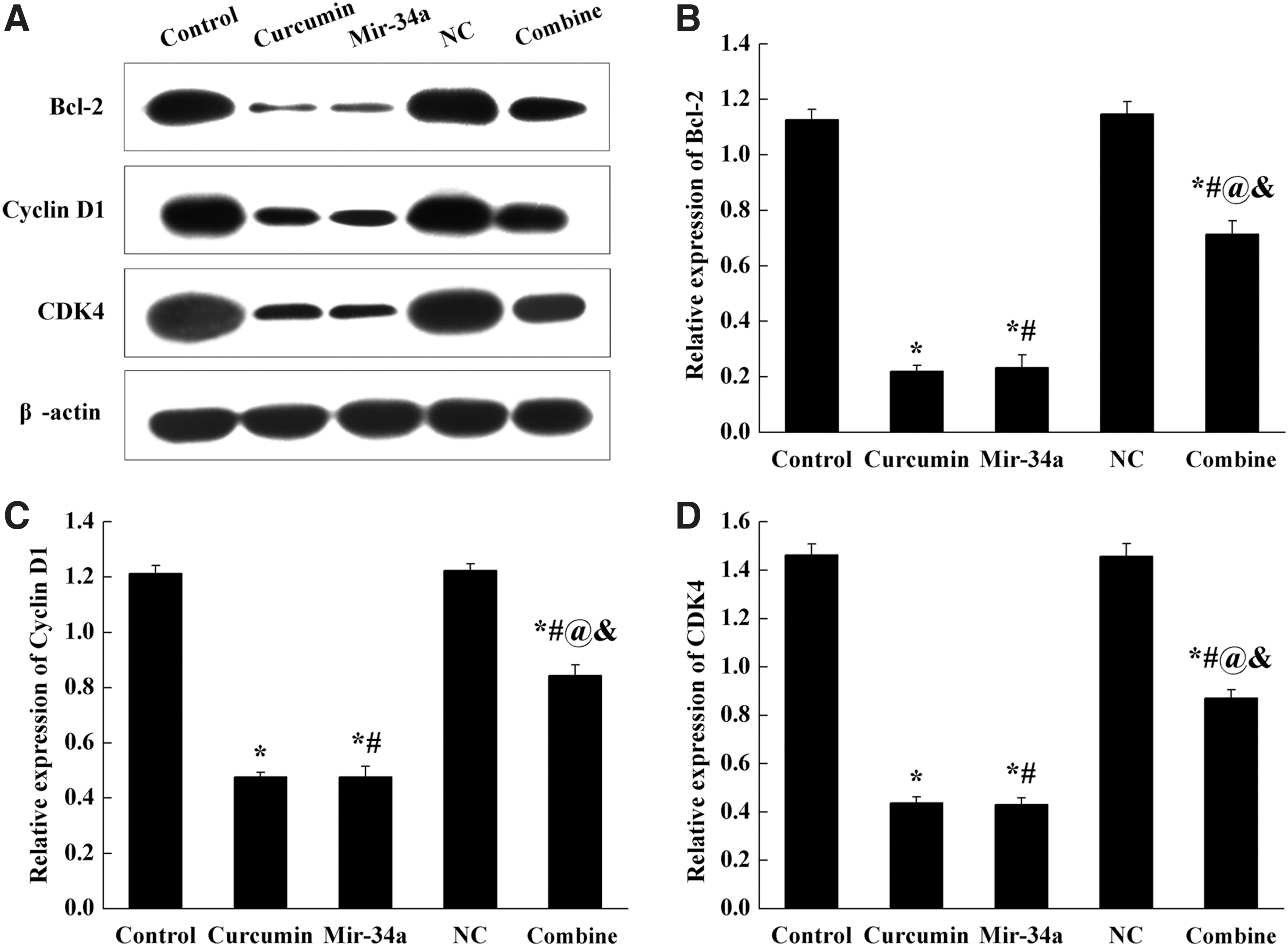
Administration of curcumin or miR-34a decreased Bcl-2, CDK4, and cyclin D1 expression in SGC-7901 cells.
Administration of curcumin or miR-34a suppressed the growth of transplanted tumors in nude mice
We established a subcutaneous transplantation tumor model in nude mice (Fig. 5). After subcutaneous inoculation of SGC-7901 cells, the tumor formation rate was 100% in control and NC groups, and the tumors grew rapidly. The tumor size and tumor weight were significantly reduced in curcumin, miR-34a, and combine groups (Fig. 5A, B, p < 0.05). Compared with curcumin and miR-34a groups, the growth rate and size of tumor in combine group were increased (Fig. 5A, B, p < 0.05). The experimental results were consistent with the trend of in vitro experiments.

Administration of curcumin or miR-34a suppressed the growth of transplanted tumors in nude mice.
TUNEL results showed that tumor cells grew vigorously and there were few apoptotic cells in the control group (Fig. 5C, D). Compared with control group, the apoptotic levels of tumor cells in curcumin, miR-34a, and combine groups were significantly increased (p < 0.05). The number of positive apoptotic cells in combine group was decreased, compared with curcumin and miR-34a groups (p < 0.05).
Administration of curcumin or miR-34a decreased Bcl-2, CDK4, and cyclin D1 in tumor tissues
As shown in Figure 6, the levels of Bcl-2, CDK4, and cyclin D1 proteins in curcumin, miR-34a, and combine groups were significantly decreased in tumor tissues (p < 0.05). While, compared with curcumin and miR-34a groups, the expression of Bcl-2, CDK4, and cyclin D1 in combine group were notably upregulated (p < 0.05).

Administration of curcumin or miR-34a decreased Bcl-2, CDK4, and cyclin D1 in tumor tissues.
Discussion
In this study, we found that curcumin could upregulate miR-34a expression and inhibit the proliferation of cells in gastric cancer. Furthermore, treatment of curcumin could decrease the levels of Bcl-2, CDK4, and cyclin D1 in gastric cancer.
Curcumin has many pharmacological effects such as antioxidation, antitumor, and so on. 13 Previous study showed that the proliferative activity of SGC-7901 cells decreased gradually with the increase of curcumin concentration and the prolongation of action time. 14 Curcumin is a lipophilic molecule that can penetrate the cell membrane rapidly and induce apoptosis. 15 These results were consistent with our study, and showed that curcumin administration suppressed the proliferation of SGC-7901 cells and reduced cell apoptosis.
We further demonstrated curcumin inhibited the proliferation of SGC-7901 cells by inhibiting the G0/G1-S phase of cell cycle, suppressing the levels of Bcl-2, CDK4, and cyclin D1 in SGC-7901 cells. Cyclin D1 can regulate the transition of cells from G1 to S phase. 16,17
CDK4 is a serine/threonine kinase that binds to cyclin D and regulates cell transition from G1 phase to S phase. 18 In many tumors, abnormal cyclinD-CDK4-INK4-Rb pathway exists. The change of this pathway speeds up the G1 phase process and enables the growth of cancer cells to accelerate to gain survival advantage. 19 Apoptosis suppressor protein can inhibit apoptosis of many malignant tumors, induce metastasis of cancer cells, and increase the invasiveness of tumors. Bcl-2 is an oncogene, which can inhibit apoptosis. 20,21 In our study, curcumin could weaken the growth of transplanted tumors and reduce the weight of tumors via downregulating the expression of Bcl-2, CDK4, and cyclin D1 in tumor tissues.
Recent studies have shown that mRNAs played an important regulatory role in cell growth, differentiation, proliferation, and apoptosis, and were associated with the occurrence and development of many tumors. 22 miR-34a played the role of tumor suppressor gene and regulates many important biological behaviors of cancer cells. 23 In this study, after transfection of miR-34a agomir into cells, it was found that miR-34a agomir treatment had similar effects on resisting malignant biological behavior. Curcumin inhibited the proliferation of SGC-7901 cells, which might be closely related to upregulating miR-34a.
In conclusion, curcumin could inhibit proliferation and induce apoptosis of gastric cancer SGC-7901 cells. Its mechanism may be related to the upregulation of miR-34a expression in gastric cancer cells.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.