
Abstract
Background:
Esophageal squamous cell carcinoma (ESCC) is one of the esophageal cancers known as an aggressive malignant tumor. Long noncoding RNAs (lncRNAs) can be involved in the progression and development of cancers. lncRNA LINC00473 (LINC00473) was reported to exert an oncogenic influence on diverse cancers. However, neither the biological function nor the underlying mechanism of LINC00473 has been explored in ESCC.
Aim of the Study:
The aim of investigation is to explore the role of LINC00473 in ESCC.
Methods:
The expression of LINC00473, miR-497-5p, and protein kinase AMP-activated alpha 1 catalytic subunit (PRKAA1) was detected by reverse-transcription quantitative polymerase chain reaction assay. Cell counting kit-8 and 5-ethynyl-2′-deoxyuridine assays were carried out to measure cell proliferation. Cell migration was tested by transwell assay. Western blot assay was performed to examine the protein expression of PRKAA1, E-cadherin, N-cadherin, and Vimentin. The binding capacity between miR-497-5p and LINC00473 (or PRKAA1) was, respectively, studied by luciferase reporter and RNA immunoprecipitation assays. Pearson correlation analysis was adopted to analyze the correlation between miR-497-5p (or LINC00473) and PRKAA1.
Results:
LINC00473 presented much higher expression and LINC00473 suppression restrained the proliferation, migration, and epithelial–mesenchymal transition (EMT) process in ESCC cells. MiR-497-5p presented lower expression, binding with and negatively regulated by LINC00473 in ESCC. PRKAA1 was confirmed as a downstream target gene for miR-497-5p. PRKAA1 could combine with miR-497-5p, and LINC00473 knockdown or miR-497-5p overexpression downregulated the mRNA and protein expression of PRKAA1. At last, the inhibitory effects of LINC00473 knockdown on proliferation, migration, and EMT process were reversed by PRKAA1 overexpression in vitro and in vivo.
Conclusions:
LINC00473 regulates ESCC progression through miR-497-5p/PRKAA1 axis, which provides a new therapeutic strategy for ESCC patients.
Introduction
Esophageal cancer (EC) has been the seventh most common cancer and the mortality of esophageal squamous cell carcinoma (ESCC) ranks sixth among EC. 1 ESCC is the main histopathological form, occupying 90% of EC. 2 Despite remarkable advances in diagnostic and surgical techniques in ESCC, the overall survival rate of ESCC patients remained ∼10% in 5-year period. 3 With high incidence and mortality, ESCC has led to a vast burden of treatment globally. 4 Therefore, it is significantly important to explore the molecular regulatory mechanism of ESCC as a new way to treat ESCC effectively.
The long noncoding RNAs (lncRNAs) belong to a class of noncoding RNA (ncRNA) with >200 nucleotides. 5 Increasing evidence has confirmed that lncRNAs played important roles in the tumorigenesis and progression of many cancers. For example, AFAP1-AS1 expression was upregulated in lung cancer and drove cell invasion and metastasis. 6 LINC00473 has been proved to function as an oncogene in various cancers. For instance, LINC00473 promotes migration and invasion of gastric cancer cells, predicting poor prognosis. 7 LINC00473 promoted tumorigenesis by sponging microRNA (miRNA) 34a to modulate ILF2 expression in cervical cancer. 8 Upregulation of LINC00473 accelerated cell proliferation and radioresistance of HNSCC cells. 9 Nevertheless, the specific function and molecular mechanism of LINC00473 in ESCC has not been explored.
miRNAs are another type of ncRNAs with <24 nucleotides. 10 Numerous evidences showed that miRNAs frequently regulated the progression of tumorigenesis. 11 –13 Specifically, miRNAs generally bind to complementary sequences of downstream target gene to exert influences on post-transcriptional process. For instance, miR-139-5p suppressed the tumorigenesis and progression by binding to HOXA9 in oral squamous carcinoma. 14 MiR-98 modulated breast cancer cell migration through targeting E2F5. 15 Recently, miR-497-5p was reported to prevent cell proliferation and invasion of angiosarcoma cells by regulating KCa3.1 expression. 16 MiR-497-5p suppresses cell development of osteosarcoma by modulating ADP ribosylation factor-like protein 2. 17 However, the molecular mechanism of miR-497-5p with ESCC is unknown.
This research was aimed to investigate the role of LINC00473 in ESCC progression. The research demonstrated that LINC00473 promoted the progression of ESCC through regulating PRKAA1 expression by functioning as a sponge for miR-497-5p, which may supply a novel therapeutic clue for ESCC patients.
Materials and Methods
Tissue samples
The ESCC samples (n = 64) and adjacent noncancerous tissues (n = 64) were obtained from patients with ESCC at Beijing Friendship Hospital, Capital Medical University. All the obtained specimens were rapidly frozen in liquid nitrogen and preserved at −80°C. None of these patients received other anticancer treatment before operation. Informed consents of this research were signed by patients before surgery, and this exploration was approved by the Ethics Committee of Beijing Friendship Hospital, Capital Medical University.
Xenograft mouse model
Nine nude Balb/c mice (male, 4 weeks of age) of 16–20 g in weight were purchased from the Laboratory Animal Centre of Huazhong Agricultural University. In the assay, the mice were divided into three groups at random (n = 3 per group): sh-NC, sh-LINC00473, and sh-LINC00473 + pcDNA3.1/PRKAA1. KYSE30 cells (1 × 107 cells in 200 μL) were injected hypodermically into the right axillary site of the mice. Either 10 μg of sh-LINC00473 or pcDNA3.1/PRKAA1 was then inoculated into the same hypodermic region where the cancer cells were implanted after hypodermic. Phosphate-buffered saline (PBS) served as a vehicle control. Tumor formation was measured every 4 d and tumor length (L) and width (W) was measured by calipers.
The tumor volume (V) was computed as follows: V = (L × W 2 )/2. All mice were executed by dislocation on day 28, and then the tumors were minced and collected for the following experiments. This investigation was approved by the committee of Beijing Friendship Hospital, Capital Medical University.
Cell culture
ESCC cell lines (KYSE30, TE-1, KYSE150, and KYSE410) were commercially obtained from the Cell Bank of the Chinese Academy of Science (Shanghai, China), and incubated in Dulbecco's modified Eagle's medium (DMEM; Gibco, Grand Island, NY) supplied with 10% fetal bovine serum (FBS; Gibco). Normal esophageal epithelial cells Het-1A were purchased from American Type Culture Collection (Manassas, VA), and cultured in DMEM medium (Gibco) added with 10% FBS (Gibco) as well. All culture was operated in a humidified atmosphere of 5% CO2 at 37°C.
Cell transfection
To downregulate the expression of LINC00473, shRNA sequences targeting LINC00473 (sh-LINC00473#1, sh-LINC00473#2, and sh-LINC00473#3) were obtained from GenePharma (Shanghai, China), with scrambled shRNA (sh-NC) as the scrambled control. MiR-497-5p mimics, negative control (NC mimics), and pcDNA3.1/PRKAA1 for overexpressing PRKAA1 were synthesized by GenePharma. All the transfection was operated by using Lipofectamine 2000 reagent (Invitrogen, Shanghai, China) in line with the manufacturer's instruction.
Real-time reverse-transcription polymerase chain reaction
According to manufacturer's guidelines, TRIzol reagent (Invitrogen, Carlsbad, CA) was utilized to extract total RNA from cultured cells. For lncRNA and mRNA analysis, a Reverse Transcription Kit (Takara, Dalian, China) was adopted to synthesize the first-strand cDNA. For miRNA analysis, a microRNA First-Strand cDNA Synthesis Kit (Sangon Biotech, Shanghai, China) was applied in the process of reverse transcription. The amplification reaction was operated in the SYBR-Green PCR Master Mix kit (Applied Biosystems, Foster City, CA) or TaqMan miRNA assay kit (Applied Biosystems) on a 7900HT Fast Real-Time System (Applied Biosystems). The relative expression of LINC00473, miR-497-5p, and PRKAA1 was computed using the 2−ΔΔCt method with GAPDH or U6 as the endogenous control.
The primer sequences used for quantitative polymerase chain reaction (qPCR) are listed as follows:
LINC00473, forward: 5′-TCA TTT CCC TAC CTG CTC CT-3′ and reverse: 5′-CAG TGT CTG CAC ATC GCT AAT-3′;
MiR-497-5p, forward: 5′-CAG CAG CAC ACU GUG GUU UGU-3′ and reverse: 5′-AAA CCA CAG UGU GCU GCU GUU-3′;
PRKAA1: forward, 5′-TGT AAG AAT GGA AGG CTG GAT GA-3′ and reverse, 5′-GGA CCA CCA TAT GCC TGT GA-3′
GAPDH: forward, 5′-GAA GGT GAA GGT CGG AGT C-3′ and reverse, 5′-GAA GAT GGT GAT GGG ATT TC-3′
U6: forward, 5′-ATT GGA ACG ATA CAG AGA AGA TT-3′ and reverse, 5′-GGA ACG CTT CAC GAA TTT G-3′
Cell-counting kit 8 assays
Cell counting kit-8 (CCK-8; Dojindo, Kumamoto, Japan) was operated to examine the proliferation of ESCC cells. Transfected TE-1 and KYSE30 cells (1 × 103 cells/well) were seeded in a 96-well plate and cultured under normal conditions. Cell proliferation was detected at 0, 24, 48, 72, and 96 h, cells were mixed with 10 μL of CCK-8 solution and further incubated for 4 h. The cell proliferation was examined by microplate reader (Bio-Tek, Winooski, VT) at 450 nm wavelength.
5-Ethynyl-2′-deoxyuridine assay
Cells were cultured in 24-well plates, and 10 μM 5-ethynyl-2′-deoxyuridine (EdU) was added to each well. Then, the cells were fixed with 4% formaldehyde for 20 min after cultured for 2 h at 37°C. After washing with PBS, the incorporated EdU was detected with a Click-iT® EdU kit for 30 min and, subsequently, the cells were stained with Hoechst 33342 for 20 min and were visualized using a fluorescence microscope (Olympus, Tokyo, Japan). The EdU incorporation rate was calculated as the ratio of the number of EdU-positive cells (red cells) to the total number of Hoechst 33342-positive cells (blue cells).
Transwell assay
For the migration examination, 1 × 104 cells in medium with free FBS were supplemented into the upper compartment of the transwell chamber (Millipore, Billerica, MA) at 24 h after transfection, whereas medium with 10% FBS was placed into the lower compartment. After being cultivated for 24 h, the rest cells on the upper surface were cleaned. Cells that had already migrated across the membrane were immobilized and stained with methanol and 0.1% crystal violet and subsequently photographed through an IX71 inverted microscope (Olympus).
Western blot assay
Cells were dissolved by radio immunoprecipitation assay buffer supplied with protease and phosphatase inhibitors (Roche, Shanghai, China). Cell proteins were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. After electrophoresis, proteins were transferred onto polyvinylidene difluoride membrane. The membrane was obstructed in 5% skimmed milk and mixed with primary antibodies (1:1000) overnight at 4°C. Primary antibodies utilized in this assay, including E-cadherin, Vimentin, GAPDH, N-cadherin, and PEKAA1, were purchased from Abcam (Hong Kong, China).
After purifying and incubation, the membranes were incubated with secondary antibodies (1:2000) at room temperature. Protein bands were detected by the ECL Chemiluminescent Detection System (Thermo Fisher Scientific, Rochester, NY) and the levels of expression were quantified by Image J software v1.46r.
Luciferase reporter assay
The 3′-UTR sequences of PRKAA1 containing the predicted miR-497-5p binding site or a mutant variant were subcloned into the pmirGLO vector (Promega, Madison, WI) to produce the wild-type PRKAA1 reporter (PRKAA1-WT) and the mutant-type PRKAA1 reporter (PRKAA1-Mut). Then, wild-type LINC00473 reporter (LINC00473-WT) and the mutant-type LINC00473 reporter (LINC00473-Mut) were also constructed. MiR-497-5p mimics or NC mimics were co-transfected with LINC00473-Wt or LINC00473-Mut into TE-1 and KYSE30 cells by the use of Lipofectamine 2000. NC mimics or miR-497-5p mimics were also co-transfected with PRKAA1-Wt or PRKAA1-Mut into TE-1 or KYSE30 cells. After 48 h of incubation, the luciferase activities were measured by the use of Dual-Luciferase Reporter Assay System (Promega).
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) assay was carried out through the use of Magna RNA-binding protein immunoprecipitation kit (Millipore). In brief, cell lysate (TE-1 and KYSE30 cells) was incubated in RIP buffer containing magnetic beads conjugating human anti-Ago2 antibody. Immunoglobulin G (IgG) was used as internal control. Immunoprecipitated RNA was isolated, purified, and analyzed by reverse-transcription quantitative polymerase chain reaction (RT-qPCR) analysis.
Xenograft mouse model
Nine nude Balb/c mice (male, 4 weeks of age) with 16–20 g in weight were purchased from the Laboratory Animal Centre of Huazhong Agricultural University. In the assay, the mice were divided into three groups at random (n = 3 per group): sh-NC, sh-LINC00473, and sh-LINC00473 + pcDNA3.1/PRKAA1. KYSE30 cells (1 × 107 cells in 200 μL) were injected hypodermically into the right axillary site of the mice. Either 10 μg of sh-LINC00473 or pcDNA3.1/PRKAA1 was then inoculated into the same hypodermic region where the cancer cells were implanted after hypodermic. PBS served as a vehicle control. Tumor formation was measured every 4 d and tumor length (L) and width (W) were measured by calipers.
The tumor volume (V) was computed as follows: V = (L × W 2 )/2. All mice were executed by dislocation on day 28, and then the tumors were minced and collected for the following experiments. This investigation was approved by the committee of Beijing Friendship Hospital, Capital Medical University.
Immunohistochemistry
Ki-67 expression served as a sign of the degree of tumor cell proliferation, which was assessed by immunohistochemistry in tumor tissues. 18,19 The mixture of tumor tissue fragments (5 μm in thickness) and citrate buffer (pH = 6) were cultured for 20 min for antigen retrieval. Subsequently, the tissues were incubated in a humidified chamber with 3% H2O2 for 10 min at room temperature to eliminate endogenous peroxidase activity. The tissues were rinsed for three times with PBS and cultivated with Ki-67 antibody (PAB19919, 1:200; Bioswamp, Wuhan, China) in a humid incubator at 4°C overnight. Next, the tissues were cultured for 30 min at room temperature with MaxVision TM HRP-Polymer anti-mouse/rabbit IHC antibody (PAB160022, 1:200; Bioswamp) in a humid chamber, and then added with diaminobenzidine.
After the core being counterstained with hematoxylin for 3 min, a graded density series (75%/85%/95%/100%) of ethanol and xylene were exploited to dehydrate the slides that were mounted with neutral balsam. The results were analyzed using a bright-field microscope (Olympus).
Statistical analysis
Data were shown as the mean ± standard deviation. Variances among groups are compared by the one-way analysis of variance or Student's t-test. SPSS 20.0 software (SPSS, Chicago, IL) was applied for statistical analysis. Correlation between miR-497-5p (or LINC00473) and PRKAA1 were analyzed with Pearson correlation analysis. All the experiments were operated for three times. Any value of p < 0.05 was considered statistically significant.
Result
LINC00473 presents much higher expression in ESCC cells and drives ESCC progression
LINC00473 expression has been noticed to be upregulated in several cancers, whereas its expression level in ESCC has not been explored. To measure the expression of LINC00473 in human ESCC, RT-qPCR was carried out in ESCC tissues and adjacent normal tissues. ESCC tissues presented higher expression than normal tissues (Fig. 1A). According to Figure 1B, LINC00473 expression was prominently upregulated in ESCC cells (KYSE30, TE-1, KYSE150, and KYSE410) compared with adjacent normal esophageal epithelial cells (Het-1A). The results also revealed that KYSE30 and TE-1 cells showed the higher expression of LINC00473 in five ESCC cell lines. Based on the results, KYSE30 cells and TE-1 cell lines were chosen for further research.
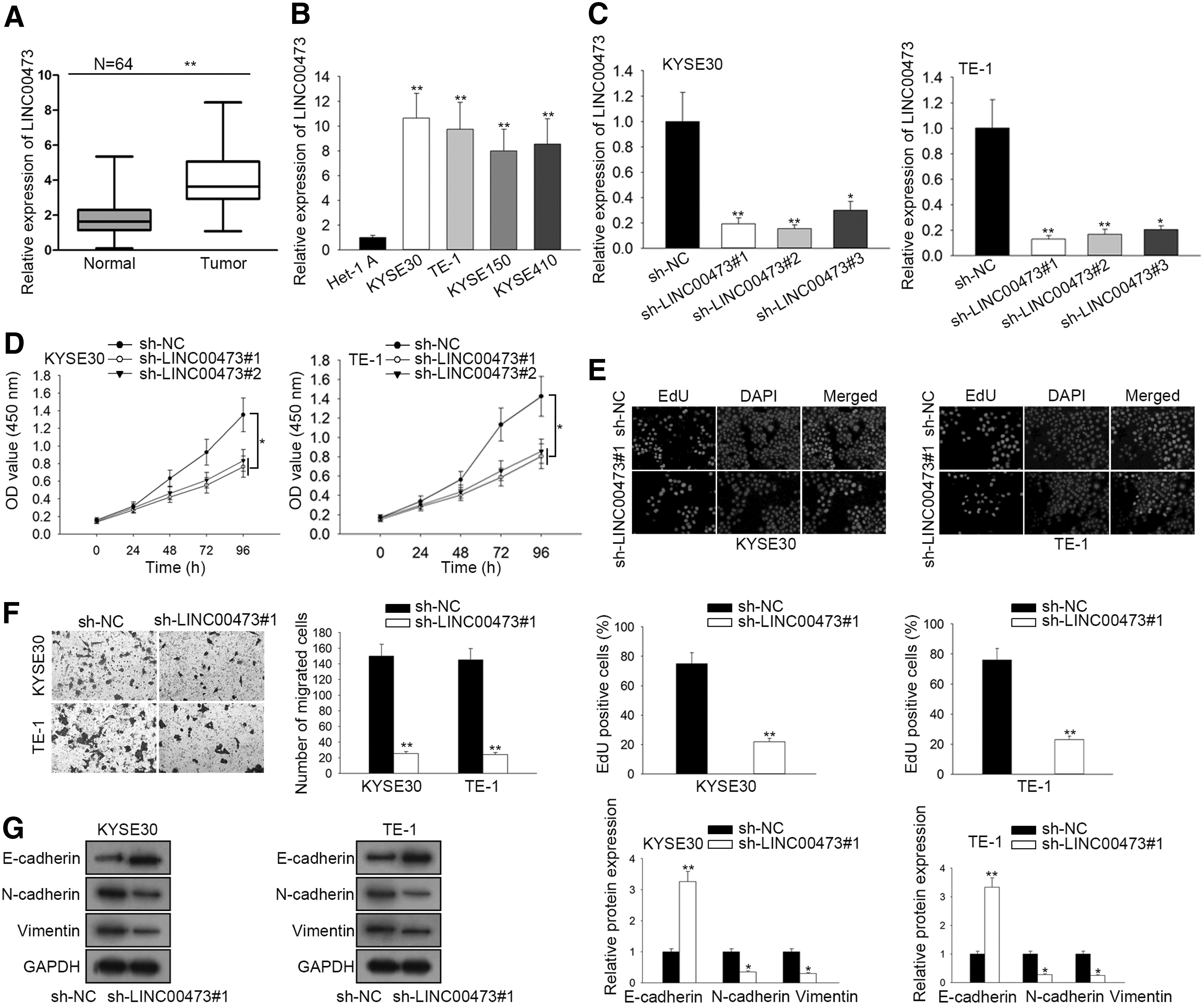
LINC00473 presents much higher expression and LINC00473 knockdown suppresses ESCC progression.
To investigate the biological role of LINC00473 in ESCC, LINC00473 was knocked down in ESCC cells. The transfection of sh-LINC00473#1/2/3 triggered an obvious reduction of LINC00473 expression in KYSE30 and TE-1 cells compared with scramble control (Fig. 1C). In addition, the downregulation in sh-LINC00473#1 and sh-LINC00473#2 groups was more prominent than that in sh-LINC00473#3 group. Hence, sh-LINC00473#1 and sh-LINC00473#2 were selected for the following loss-of-function experiments. CCK-8 assay displayed that LINC00473 deficiency limited cell proliferation (Fig. 1D). In addition, EdU assay further verified that LINC00473 knockdown led to a reduction of EdU positive cells (Fig. 1E). Subsequently, transwell assay indicated that the migratory capacity of ESCC cells was conspicuously prevented by LINC00473 knockdown (Fig. 1F).
Last but not least, the results of Western blot assay demonstrated that LINC00473 deficiency lowered the protein expression of N-cadherin and Vimentin but enhanced the protein expression of E-cadherin in KYSE30 and TE-1 cells (Fig. 1G). Taken together, LINC00473 expression is upregulated and LINC00473 inhibition suppresses cell proliferation, migration, and EMT process in ESCC cells.
MiR-497-5p is sponged by LINC00473 in ESCC
Mounting studies proposed that lncRNAs could bind with miRNAs, thereby sequestering miRNAs away from downstream target gene. Therefore, it is intended to figure out the interaction between LINC00473 and miRNAs in ESCC. According to the starBase website (

MiR-497-5p is sponged by LINC00473 in ESCC.
Finally, RT-qPCR disclosed that miR-497-5p expression was raised by LINC00473 suppression and miR-497-5p overexpression caused a great depletion of LINC00473 expression in KYSE30 and TE-1 cells (Fig. 2F). Overall, LINC00473 negatively regulates miR-497-5p expression by functioning as a molecular sponge.
LINC00473 modulates PRKAA1 expression by competitively binding to miR-497-5p
MiRNAs have been reported to be involved in cancer mostly by modulating target gene. 14,20 starBase was utilized to seek potential target genes of miR-497-5p and PRKAA1 was picked for further study based on the prediction of starBase website (Fig. 3A). First, the expression of PRKAA1 in ESCC tissues and cells was apparently higher than that in normal tissues and cells, which was measured by RT-qPCR assay (Fig. 3B, C). Subsequently, luciferase reporter assay using KYSE30 and TE-1 cells demonstrated that miR-497-5p amplification decreased the luciferase activity of PRKAA1-WT vector, but no obvious alteration was observed in PRKAA1-Mut vector (Fig. 3D). In addition, RIP assay verified that miR-497-5p and PRKAA1 could be immunoprecipitated by Ago2 antibodies instead of IgG antibodies in KYSE30 and TE-1 cells (Fig. 3E).

LINC00473 modulates PRKAA1 expression by competing to bind to miR-497-5p.
To verify the interaction between miR-497-5p and PRKAA1, the mRNA expression and protein expression of PRKAA1 were, respectively, examined by RT-qPCR and Western blot. The results demonstrated that both mRNA and protein expression of PRKAA1 were downregulated in KYSE30 and TE-1 cells transfected with miR-497-5p mimics or sh-LINC00473#1 (Fig. 3F, G). Over and above that, Pearson analysis showed the negative correlation between miR-197-5p and PRKAA1, and the positive correlation between LINC00473 and PRKAA1 (Fig. 3H). In sum, LINC00473 modulates PRKAA1 expression by competitively binding to miR-497-5p.
LINC00473 modulates ESCC development by sponging miR-497-5p and targeting PRKAA1
Rescue assays were carried out to confirm whether LINC00473 aggravated ESCC development through miR-497-5p/PRKAA1 axis. At the beginning, the mRNA and protein level of PRKAA1 expression was significantly upregulated in KYSE30 cells transfected with pcDNA3.1/PRKAA1 (Fig. 4A, B). CCK-8 and EdU assays indicated that PRKAA1 overexpression markedly reversed the LINC00473 silencing-inhibited cell proliferation (Fig. 4C, D). Moreover, according to transwell assay, the restrained effect of LINC00473 deficiency on cell migration was abrogated by PRKAA1 overexpression (Fig. 4E).
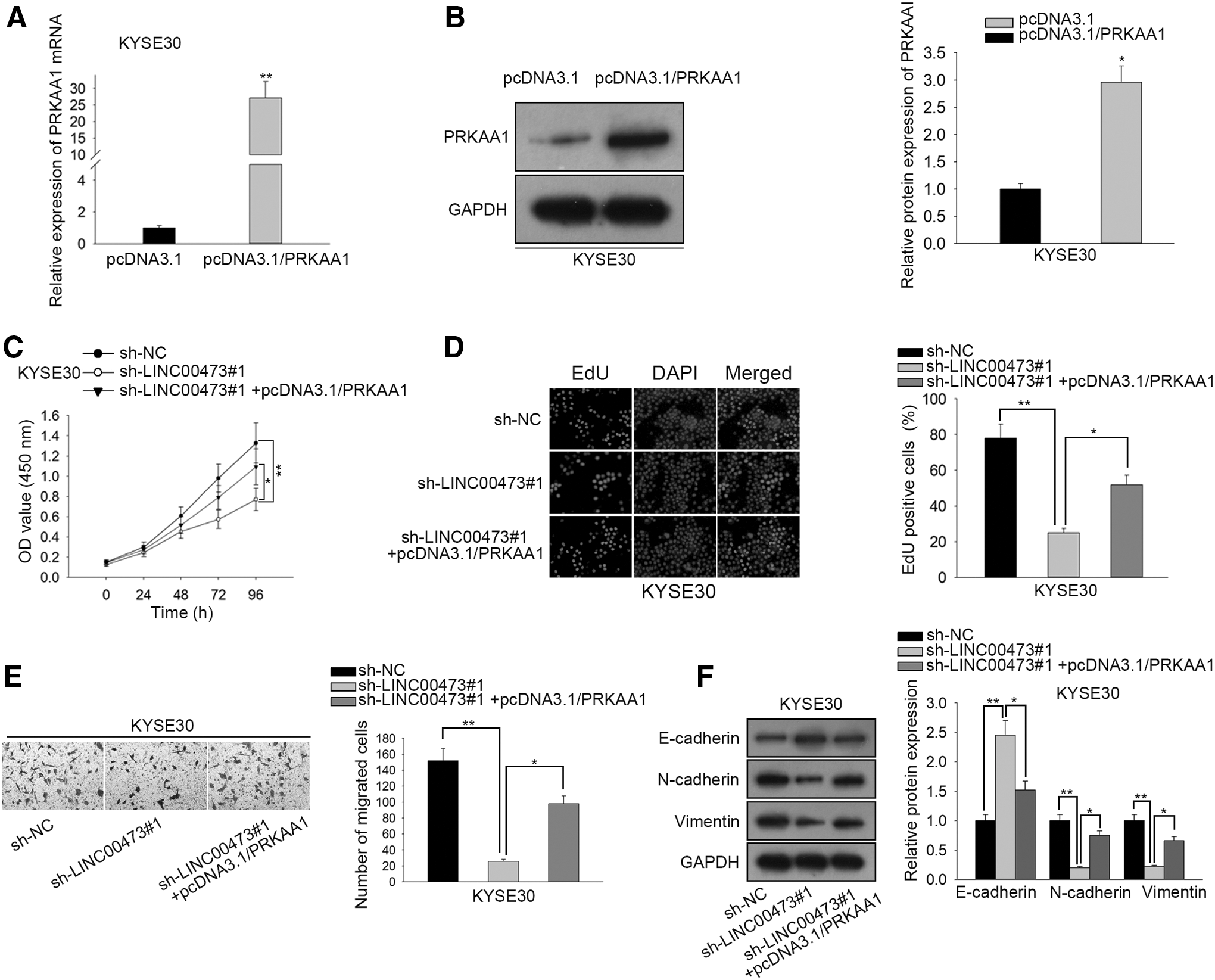
LINC00473 modulates ESCC development by sponging miR-497-5p and targeting PRKAA1.
Similarly, sh-LINC00473#1-mediated rise of E-cadherin expression and reduction of N-cadherin and Vimentin expression were countervailed by PRKAA1 overexpression (Fig. 4F). In a word, LINC00473 modulates ESCC development by sponging miR-497-5p and targeting PRKAA1.
Tumor progression is facilitated by LINC00473 in vivo
The biological function of LINC00473 in vivo was further explored. As depicted in Figure 5A, the tumor volume decreased by LINC00473 silence was reversed by PRKAA1 amplification. Moreover, the inhibitory effect of LINC00473 knockdown on tumor growth and weight was abrogated by transfection of pcDNA3.1/PRKAA1 (Fig. 5B, C). Eventually, the decline of protein expression of Ki67 resulted from LINC00473 deficiency was restored by PRKAA1 overexpression (Fig. 5D). All in all, tumor progression is facilitated by LINC00473 and PRKAA1 in vivo.
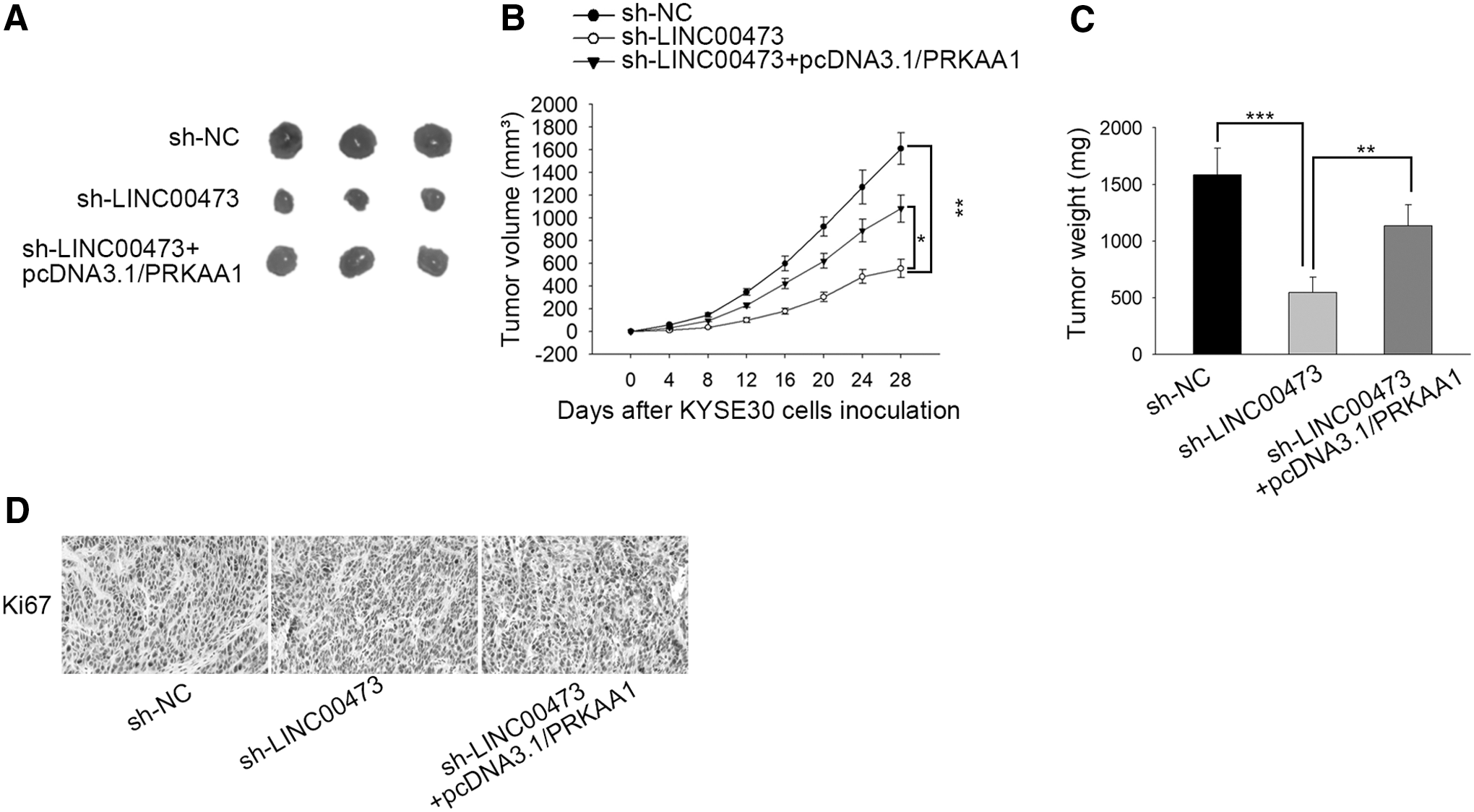
Tumor progression is facilitated by LINC00473 in vivo.
Discussion
Prevenient research has revealed that lncRNAs participate in different types of cancers, such as gastric cancer, breast cancer, and bladder cancer. 21 –23 LINC00473 was reported to indicate a poor prognosis and modulate cell metastasis in gastric cancer. 7 LINC00473 drove taxol resistance in CRC by sponging miR-15a. 24 Although LINC00473 was reported to weaken the effect of radiotherapy in ESCC through miR-347a-5p/SP1N1 axis, 25 the molecular mechanism of LINC00473 remained complicated and the biological function of LINC00473 involved in cellular processes in ESCC deserved a further investigation. In this investigation, it is found that the expression of LINC00473 was obviously higher in ESCC tissues and cell lines. What is more, knockdown of LINC00473 limited the proliferation, migration, and EMT process in ESCC cells.
A large body of evidence indicated that lncRNAs can act as a ceRNA to prevent subsequent miRNAs combining with target mRNAs. For example, LINC01287/miR-298/STAT3 axis modulated growth of hepatocellular carcinoma cells. 26 lncRNA MIR31HG inhibited hepatocellular carcinoma progression by targeting miRNA-575/ST7L axis. 27 In this research, the author predicted and verified LINC00473 can bind with miR-497-5p, which has been reported to inhibit tumorigenesis and promotion of cancer. 28,29 Moreover, LINC00473 negatively regulated miR-497-5p expression by functioning as its sponge in ESCC.
Protein kinase AMP-activated alpha 1 catalytic subunit (PRKAA1), also known as AMPK, plays critical roles in modulating the activities of a series of key metabolic enzymes, and recently has been reported to be oncogenic in various cancers. 30 As example, PRKAA1 played an important role in gastric promotion, 31 and predicts poor prognosis for colorectal cancer patients. 32 Nevertheless, there is no published study on PRKAA1 in ESCC. In this investigation, PRKAA1 was proved to be a downstream target gene of miR-497-5p, inhibited by miR-497-5p but upregulated by LINC00473 in ESCC. Finally, rescue assays indicated that overexpression of PRKAA1 partially rescued LINC00473 knockdown-mediated inhibition on ESCC progression.
In summary, this research was the initial exploration of LINC00473 involved in cellular process of ESCC cells. However, other regulatory mechanism remained to be explored. This study delineated that LINC00473 facilitates cell proliferation, migration, and EMT process in ESCC by modulating miR-497-5p/PRKAA1 axis, indicating the potential LINC00473/miR-497-5p/PRKAA1 axis may offer a new therapeutic target to improve the treatment of ESCC patients.
Footnotes
Acknowledgment
The author thanks all the participants.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This study was supported by Beijing Friendship Hospital Research Launch Fund (yyqdkt2016-17).