
Abstract
Objective:
Long non-coding RNAs (lncRNAs) are a new class of noncoding RNAs, which serve as crucial regulators in tumor progression, including osteosarcoma (OS). The objective of this study is to uncover the clinical significance and biological role of lncRNA CDKN2B antisense RNA 1 (CDKN2B-AS1) in OS.
Methods:
Quantitative real time PCR analysis was used to determine the expression of CDKN2B-AS1 in OS tissues and cell lines. The associations between CDKN2B-AS1 expression and clinicopathological characteristics of OS patients were assessed by Chi-squared test. CCK-8, colony formation, flow cytometry and transwell assay were utilized to evaluate the effects of CDKN2B-AS1 knockdown on cell proliferation, cell cycle, migration and invasion. The protein expression associated with cell cycle and epithelial-mesenchymal transition (EMT) was measured by western blotting.
Results:
CDKN2B-AS1 was found to be markedly up-regulated in OS tissues and cell lines. Clinical data further demonstrated highly expressed CDKN2B-AS1 tended to be strongly positively correlated with tumor size, distant metastasis and TNM stage. Loss-of-function of CDKN2B-AS1 leaded to inhibited cell proliferation and induced cell cycle G0/G1 phase arrest. In addition, CDKN2B-AS1 knockdown significantly suppressed OS cells migration and invasion. Mechanistically, CDKN2B-AS1 knockdown in OS cells suppressed the expression of CDK4 and Cyclin D1, as well as EMT, as demonstrated by elevated levels of epithelial markers (E-cadherin) and downregulation of mesenchymal markers (vimentin and N-cadherin).
Conclusions:
Taken together, our findings suggest that CDKN2B-AS1 represents a potential therapeutic target for OS.
Introduction
Long noncoding RNAs (lncRNAs) belong to a subset of long (>200 nucleotides) RNA molecules, which play important roles in chromatin organization, transcription, and translational modulation. 1 Since the serendipitous discovery of the first lncRNA, the field of lncRNAs involved in the etiology and pathophysiology of diseases has been expanded greatly. 2 LncRNAs have been shown in some previous studies to trigger malignant phenotypes of cancer cells through crosstalk with macromolecular substances, such as DNA, RNA, and protein. 3 On the other hand, deficiency or amplification of lncRNAs in malignant cells can lead to oncogenesis via downregulating tumor suppressors and/or upregulating oncogenes. 4 For instance, downregulation of GAS5, a newly identified lncRNA, could reduce p21 protein level and promote cell cycle progression in stomach cancer. 5 Enhanced expression of lncRNA H19 can induce the inhibitor of DNA binding/differentiation 2, thus contributing to bladder cancer cell proliferation. 6 It is generally believed that lncRNAs, not only mediates the cancer cell abilities for tumor growth and metastasis, but can also provide diagnostic and prognostic values for cancer patients. 7 For example, overexpression of HULC, 8 TUG1, 9 and UCA1 10 in osteosarcoma (OS) patients are highly predictive of progression to advanced clinical stage and poor prognosis. However, the biological function and diagnostic/prognostic potential of only a few lncRNA in OS, has been studied.
OS is a rare but devastating disease that originates from primitive mesenchymal cells formed by bone. There has been an age-specific incidence rates of OS with the first peak in adolescence between 10 and 14 years of age and the second peak among adults aged >65 years. 11 This tumor predominantly occurs in the long bone of the limbs, such as femur and the tibia. 11 Surgery resection in combination with multi-agent, dose-intensive chemotherapy achieves a satisfactory long-term prognosis in most patients with localized OS. 12 However, pulmonary metastasis is observed in 50% of OS patients, which often led to very poor outcome, with no more than 20% 5-year survival. 13 Hence, exploration of the molecular mechanisms underlying disease progression and clinical applications for suppressing metastasis is currently hot research topics in the control of OS development.
CDKN2B antisense RNA 1 (shortened to CDKN2B-AS1, also known as ANRIL) is a recently discovered lncRNA that is located on chromosome 9q21 region. 14 In 2011, Burdon et al. 14 reported a genome-wide analysis of open-angle glaucoma blindness in 590 individuals and 3956 controls, and discovered two susceptibility locus at CDKN2B-AS1 and TMCO1 in the Australia population. Besides, intronic polymorphisms in the CDKN2B-AS1 for myocardial infraction and coronary artery disease were revealed by AbdulAzeez et al. 15 Recently, a growing number of research showed that CDKN2B-AS1 levels were upregulated in different cancer cells, including thyroid cancer, 16 epithelial ovarian cancer, 17 cervical cancer, 18 and prostate cancer. 19 CDKN2B-AS1 was proposed to have oncogenic properties, due to its ability to promote tumorigenicity. 20 Given the clinicopathological significance of CDKN2B-AS1 in several types of malignancies, detecting the expression level of CDKN2B-AS1 might help us evaluate the progression stage of cancer and predict the survival time in individual patients. 20 However, the functional and clinicopathologic significance of CDKN2B-AS1 in OS remains unclear.
In this study, we sought to investigate lncRNA CDKN2B-AS1 expression and clinicopathologic features in OS to improve our understanding of OS etiology and its diagnosis. Here, we determined the expression levels of CDKN2B-AS1 in OS tissues and cell lines. Then, the association between CDKN2B-AS1 expression and clinicopathological characteristics in OS patients was analyzed. Finally, cell growth, cell cycle progression, migration, and invasion following knockdown of CDKN2B-AS1 were explored in OS cells.
Materials and Methods
Patient sample collection
Fresh OS tumors and matched adjacent normal tissues from 56 patients were collected from the Department of Orthopaedics, Renmin Hospital of Wuhan University (Hubei, China). Meanwhile, the basic clinical characteristics, including gender, age, and histological type were also obtained from each patient and summarized in Table 1. All specimen tissues were snap frozen in liquid nitrogen instantly following surgical resection and stored at −80°C until further analysis. All participants signed written informed consent documentation. This trial was conducted in accordance with the Institutional Review Board of Renmin Hospital of Wuhan University.
The Association Between Clinicopathological Characteristics and the Expression of CDKN2B-AS1 in Osteosarcoma Patients
Low or high CDKN2B-AS1 expression group was classified by the median expression of all specimens. p-Values less than 0.05 are shown in bold.
OS, osteosarcoma; TNM, tumor-node-metastasis.
RNA extraction and quantitative real-time PCR
Total RNA was isolated from frozen tissues using TRIzol reagent (Sigma-Aldrich, SF, CA). For RNA transcription to cDNA, the PrimeScriptTM RT Reagent Kit (Takara Bio, Inc., Otsu, Japan) was used according to the manufacturer's instructions. Then, the expression of CDKN2B-AS1 was determined using Takara SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) kit on an Applied Biosystems 7300 Fast Real-time PCR System. The thermocycling conditions were 95°C for 10 min, followed by 40 cycles at 95°C for 15 s, 60°C for 40 s. Each assay above was performed in triplicate. Experiments were performed at least three times. Relative gene expression was analyzed by the 2−ΔΔCt method. Quantification of CDKN2B-AS1 expression was normalized to GAPDH. Real-time PCR primer sequences are as follows: CDKN2B-AS1 forward: 5′-CTATCCGCCAATCAGGAGGC-3′; reverse: 5′-AAAAGGGACACTAGTCCGGC-3′; GAPDH forward: 5′-ACTAGGCGCTCACTGTTCTC-3′; reverse: 5′-ATCCGTTGACTCCGACCTTC-3′.
Cell culture
Four strains of human OS (SW1353, U2OS, MG-63, and Saos-2) and a normal osteoblast cell line hFOB1.19 were provided by American Type Culture Collection (ATCC, Manassas, VA). Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum was used as culture medium for all cell lines. These cells were grown in a humidified incubator containing 5% CO2 at 37°C.
Transient transfection
The small interfering RNAs that specifically target human CDKN2B-AS1 (si-CDKN2B-AS1: 5′-CAUUACCUCCUUUGGGAAUTT-3′) and negative control (si-NC: 5′-AUACCAAAUCAGGUAGGUGTT-3′) plasmids were purchased from GenePharma, Shanghai, China. On the day of transfection, U2OS or MG-63 cells at 5 × 105 per well were seeded in six-well culture dishes and subsequently transfected with 50 nM si-CDKN2B-AS1 or si-NC for 48 h. For delivering siRNA into cells, Lipofectamine 2000 (Invitrogen) was used per the manufacturer's protocol.
Cell counting kit-8 assay
Proliferation ability of OS cells was measured by cell counting kit-8 (CCK-8) assay (Dojindo, Japan). In brief, transfected cells were dispensed into 96-well plates at 3000 per well with five replicate wells. Aliquots of 10 μL CCK-8 reagent were transferred into each well and plates were incubated for 2 h at 37°C. Absorbance intensities at 450 nm were determined using a microplate reader (Bio-Tek, Winooski, VT).
Colony formation assay
For colony formation assay, transfected OS cells were added to six-well plate at a density of 500 cells per well and cultured for consecutive 2 weeks. The naturally formed colonies (at least 50 cells per colony) were fixed for 30 min with 4% paraformaldehyde and stained for 30 min with 0.5% crystal violet at room temperature. Five fields were randomly selected for observation and counting of cell colonies.
Cell cycle assay
For cell cycle, transfected OS cells were seeded into six-well plates, washed with PBS, and fixed with 70% ice-cold ethanol in PBS overnight at 4°C. After washing with PBS three times, cells were incubated with 50 μg/mL RNAase and 50 μg/mL propidium iodide for 30 min at room temperature and in darkness. The percentage of cells in G0/G1, S, and G2/M phases of the cell cycle was analyzed using the BD Biosciences FACSCalibur flow cytometer (San Jose, CA) equipped with ModFit LT software (Verity Software House, ME).
Cell migration and invasion assay
For cell migration assay, transfected cells (1 × 106 cells) were trypsinized, resuspended in 2 mL of serum-free media, and seeded into the upper transwell chambers (8 μm pores; Corning, Inc., Corning, NY). Total 2 mL of media containing 10% fetal bovine serum was added in the lower chambers of the transwell. After 24 h incubation, the cells in the upper chamber were scraped gently. Then, the cells that migrated to the lower membrane were counted in five randomly selected fields under a microscope after fixation in methanol and staining with 0.1% crystal violet. For cell invasion assay, the protocols were similar to the transwell migration assay, except for the upper surface of chambers that were coated with Matrigel (BD Biosciences).
Western blotting
Total protein was extracted using iced RIPA buffer (Beyotime Biotechnology, Jiangsu, China) and determination of protein concentration was performed using a BCA protein assay kit (Beyotime Biotechnology Co.,) following the manufacturer's instructions. Subsequently, a total of 30 μg protein was separated by 10% SDS-PAGE and transferred to nitrocellulose membrane (Millipore, MA). After blocking, membranes were incubated with primary antibodies against CDK4, Cyclin D1, E-cadherin, N-cadherin, Vimentin, and GAPDH overnight at 4°C, and then peroxidase-conjugated secondary antibodies were added and incubated at room temperature for 2 h. Then, the protein signals were visualized with ECL reagents (Sigma-Aldrich; Merck KGaA).
Statistical analysis
All statistical analyses were performed using the SPSS version 17.0 for windows (SPSS, Inc., Chicago, IL). Comparison of CDKN2B-AS1 expression between tumor tissues and adjacent tissues were performed using the Wilcoxon Signed Rank test. Besides, the associations between CDKN2B-AS1 expression and clinicopathological characteristics of OS patients were assessed by chi-squared test. The quantitative data were expressed as mean ± SD. Student's t-test was performed to determinate differences between two groups, and one-way ANOVA analysis was used between three or more groups. The p-value <0.05 was considered as statistically significant.
Results
CDKN2B-AS1 was expressed at high levels in OS tissues and cell lines
To investigate whether CDKN2B-AS1 plays an important role in OS, the expression level of CDKN2B-AS1 in 58 pairs of OS tissues and adjacent noncancerous tissues was analyzed by quantitative real-time PCR. From Figure 1A, we can see that CDKN2B-AS1 was markedly upregulated in OS tissues in comparison with adjacent noncancerous tissues (p < 0.001). Moreover, the expression levels of CDKN2B-AS1 were all significantly upregulated in four OS cell lines, compared with the normal osteoblastic cell line hFOB1.19 (Fig. 1B). Among these analyzed OS cell lines, U2OS and MG-63 cells showed much higher levels of CDKN2B-AS1 expression and thus were chosen for further experiments.

Osteosarcoma specimens and cell lines exhibit higher CDKN2B-AS1 expression level.
CDKN2B-AS1 was correlated with clinical progression in OS patients
The relationship between CDKN2B-AS1 expression and the clinicopathological characteristics in OS patients were then determined. The OS patients were divided into high CDKN2B-AS1 group (n = 35) and low CDKN2B-AS1 group (n = 23) according to the median CDKN2B-AS1 expression levels as a cutoff value. Subsequently, associations between CDKN2B-AS1 expression (high and low) and the clinical data of patients with OS were analyzed by chi-squared test. As shown in Table 1, high-level expression of CDKN2B-AS1 was significantly associated with tumor-node-metastasis (TNM) stage (p = 0.016), tumor size (p = 0.036), and the risk of distant metastasis (p = 0.023), but had no statistical association with gender (p = 0.667), age (p = 0.625), anatomic location (p = 0.075), and histological type (p = 0.364).
Knockdown of CDKN2B-AS1 suppressed cell proliferation and triggered G0/G1 cell cycle arrest in OS cells
To confirm whether CDKN2B-AS1 plays a positive role in OS in vitro, stable CDKN2B-AS1 knockdown was established in U2OS and MG-63 cells using siRNA transfection, followed by quantitative real-time PCR validation in these two cell lines (Fig. 2A, B). The results of CCK-8 assay are shown in Figure 2C and D and it is clear that knockdown of CDKN2B-AS1 could strongly suppress U2OS and MG-63 cells proliferation. Moreover, colony formation assay further demonstrated that the U2OS and MG-63 cells in the si-CDKN2B-AS1 group had reduced numbers of colonies compared with the si-NC group (Fig. 2E). We next analyzed whether impaired cell proliferation was correlated with cell cycle alteration by CDKN2B-AS1 knockdown. As expected, si-CDKN2B-AS1 caused a significant alteration in cell cycle distribution: the fraction of U2OS and MG-63 cells in G0/G1 phase increased, while the fraction of two types of OS cells in S and G2/M phase decreased compared with si-NC transfection (Fig. 3A, B). These results indicated CDKN2B-AS1 played positive role in OS cell growth and proliferation.
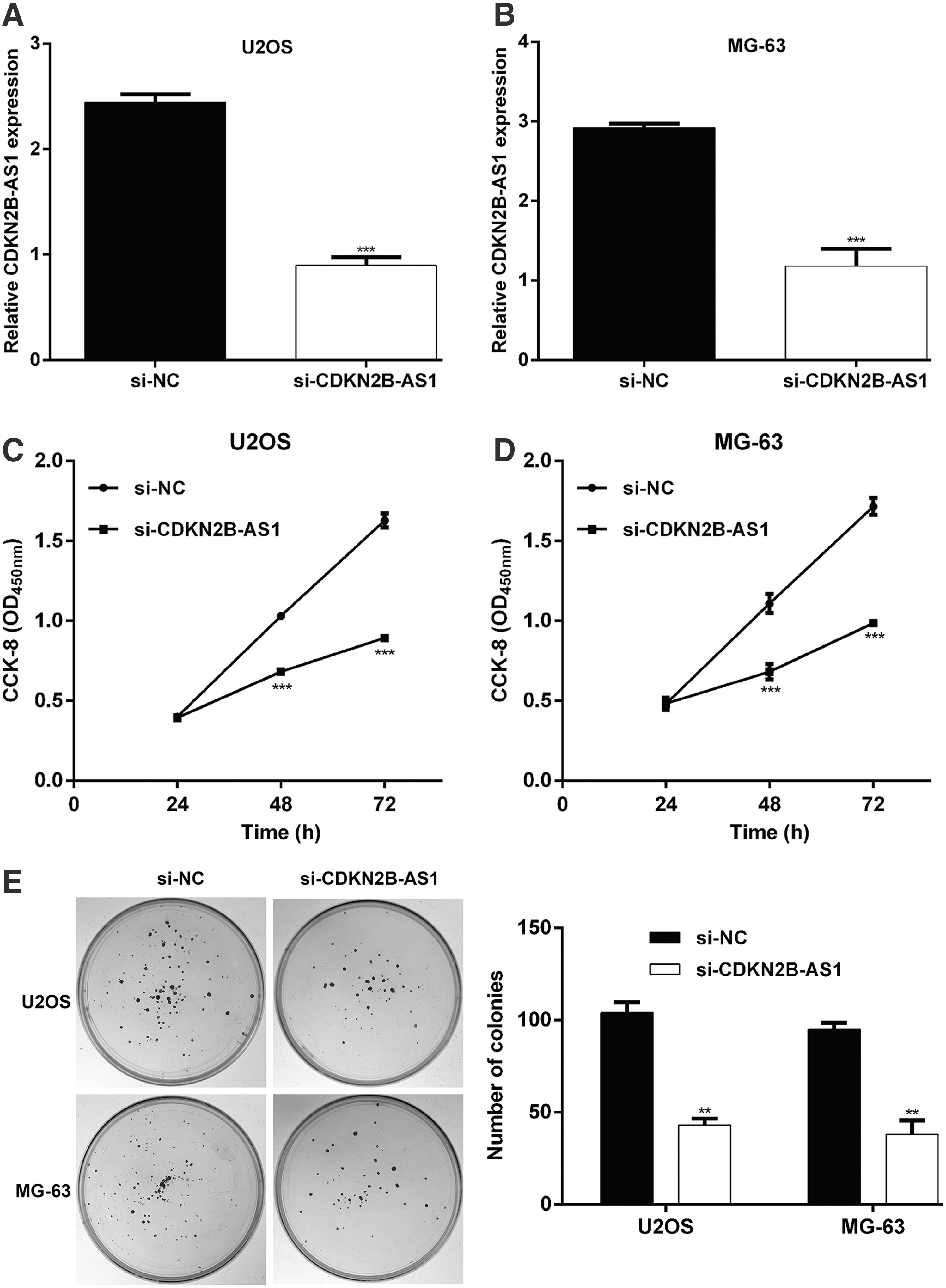
CDKN2B-AS1 knockdown suppressed cell proliferation in OS cells.

CDKN2B-AS1 knockdown induced G0/G1 phase arrest in OS cells. U2OS and MG-63 cells were transfected with si-CDKN2B-AS1 or si-NC, respectively. Flow cytometry was used to explore cell cycle distribution in U2OS
Knockdown of CDKN2B-AS1 leads to diminished migration and invasion of OS cells
In addition, changes in the migratory capability of OS cells with CDKN2B-AS1 knockdown were measured with a cell migration assay using transwell inserts. As depicted in Figure 4A, the migrated number of U2OS and MG-63 cells was decreased (39.3 ± 3.1 and 53.3 ± 7.4, respectively) in si-CDKN2B-AS1 group compared with the si-NC group (102.7 ± 7.0 and 124.0 ± 5.6, respectively). Furthermore, CDKN2B-AS1 knockdown also impaired cell invasion (Fig. 4B), as reflected by decreased invasive cell number in si-CDKN2B-AS1 group compared with control group (U2OS: 25.3 ± 5.5 vs. 75.0 ± 6.0; MG-63: 22.0 ± 3.0 vs. 58.7 ± 4.5).
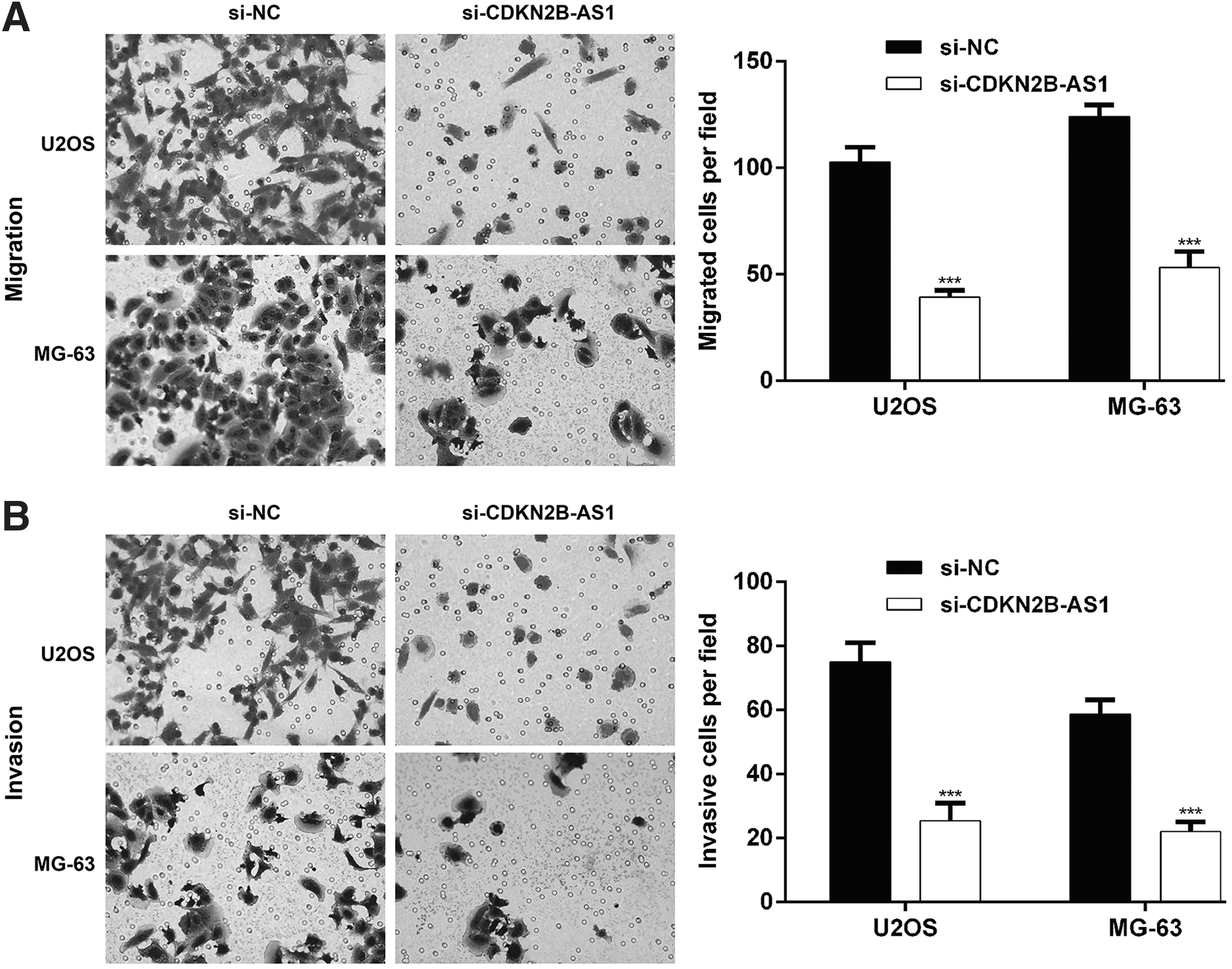
Migration and invasion of OS cells after CDKN2B-AS1 knockdown. U2OS and MG-63 cells were transfected with si-CDKN2B-AS1 or si-NC, respectively.
CDKN2B-AS1 knockdown regulated the molecular levels associated with cell cycle progression and epithelial-mesenchymal transition
To reveal the molecular mechanisms underlying CDKN2B-AS1 depletion affecting cell proliferation and invasion we used Western blot to analyze the expression of downstream molecules associated with cell cycle and epithelial-mesenchymal transition (EMT). The results indicated that CDK4 and Cyclin D1, associated with G0-S transition, were significantly downregulated in U2OS (Fig. 5A) and MG-63 cells (Fig. 5B) after knockdown of CDKN2B-AS1. Additionally, we found CDKN2B-AS1 knockdown suppressed the expression of EMT in both U2OS and MG-63 cells, as reflected as increased E-cadherin, decreased N-cadherin and Vimentin.

CDKN2B-AS1 knockdown regulated the molecular levels associated with cell cycle progression and EMT. U2OS and MG-63 cells were transfected with si-CDKN2B-AS1 or si-NC, respectively. Western blot analysis showed CDKN2B-AS1 knockdown downregulated CDK4, Cyclin D1, N-cadherin, Vimentin, but upregulated E-cadherin in U2OS
Discussion
In recent 10 years, accumulating evidence indicates that deregulation of lncRNA contributes to the initiation and progression of most, if not all, human malignancies. 21 LncRNAs have an important clinical application in identifying and characterizing the disease since their earliest discovery. 22 To date, several studies have explored the association between polymorphisms in the lncRNA CDKN2B-AS1 and diseases such as glaucoma, myocardial infarction, and coronary artery. 14,15 Multitudinous literatures have demonstrated that there is abnormal expression of CDKN2B-AS1 in human cancers. For example, CDKN2B-AS1 was found to be abnormally high expressed in cervical cancer and closely linked to advanced clinical stage, lymph node metastasis, and poor outcome. 23 Qiu et al. 24 have found that patients with high CDKN2B-AS1 had serous ovarian cancer with a higher histological grade and a poorer prognosis compared with those with low CDKN2B-AS1. In addition, high levels of CDKN2B-AS1 is seen in surgical specimens in colorectal cancer compared with normal colorectal mucosae, and the presence of CDKN2B-AS1 amplification was correlated with TNM staging, Duke staging, and lymphatic metastasis. 25 Even though previous investigations have revealed that CDKN2B-AS1 acts as an important player in multiple types of cancer the role of CDKN2B-AS1 in OS has not been yet well characterized.
In this study, we characterized the expression and clinical significance of CDKN2B-AS1 in OS. We first observed that the expression of CDKN2B-AS1 was statistically significant elevated in OS tissues and cell lines to a greater extent than matched nontumor adjacent tissues and normal osteoblast cell line hFOB1.19. Moreover, we found that high levels of CDKN2B-AS1 mRNA showed significant correlation with tumor size, distant metastasis, and TNM stage. Our results indicated that CDKN2B-AS1 expression might have an important clinical utility that can be used as a differential diagnostic biomarker for metastatic and advanced OS.
Previous studies have shown that CDKN2B-AS1 implicate in multiple cellular processes associated with tumorigenesis such as those impacting proliferation, apoptosis, invasion, metastasis, and cell cycle progression of the cells. 26 To determine the influence of CDKN2B-AS1 on the biological behavior of the OS cells, the in vitro experiments were carried out. Our data showed knockdown of CDKN2B-AS1 could depress OS cells viability, impair migration and invasion capabilities, and induce G0/G1 cell cycle arrest. Furthermore, western blot revealed that CDKN2B-AS1 knockdown acts to diminish CDK4 and Cyclin D1 protein levels. The complex formed by CDK4 and Cyclin D1 is indispensable for the G1-to-S phase transition. 27 We suggest that CDKN2B-AS1 depletion triggered cell cycle arrest at G0/G1 phase through depressing CDK4-Cyclin D1 complex formation. Our results also confirmed that knockdown of CDKN2B-AS1 attenuated the OS cells EMT by upregulating the E-cadherin expression and downregulating the Vimentin and N-cadherin expression. These data imply that the impaired EMT process is the main reason for the lack of invasion and migration of CDKN2B-AS1-knockdown OS cells.
Mechanically, CDKN2B-AS1 appears to function in the regulation of epigenetic through at least three different mechanisms: (1) recruits SUZ12 in the Polycomb repressive complex 2 (RPC2) and depressing the expression of the CDKN2B locus 28 ; (2) binds directly to CBX7, a member of Polycomb repressive complex1 (RPC1), and inhibits the CDKN2A/B loci 29 ; (3) participates in DNA methylation of some loci in differentiated cells. 30 It is noteworthy that three tumor suppressors, p15INK4b, p16INK4a, and ARF are encoded by CDKN2A/B (INK4-ARF). 31 Importantly, the active CDK/Cyclin complex can be suppressed by several CDK inhibitors, including p15INK4b and p16INK4a. 32 In colorectal cancer, p16INK4a deficiency can lead to worse outcomes in terms of tumor progression, lymphatic invasion, and survival. 25,33 Additionally, CDKN2B-AS1 has been reported to promote tumorigenesis through silencing KLF2, P21, and miR-99a/miR-499a, along with regulating PI3K/Akt and TGF-β/Smad signaling pathways. 16,23,34,35 In fact, further investigation is still needed that will clarify whether these mechanisms correlate with the functional role of CDKN2B-AS1 in OS.
In conclusion, our study examined and verified that CDKN2B-AS1 was highly expressed in OS tissues and cells. Amplification of CDKN2B-AS1 is linked to tumor size, distant metastasis, and TNM stage. Functional studies indicate a key role of CDKN2B-AS1 in the promotion of tumor growth, aggressive migratory and invasive phenotypes, and cell cycle progression through regulation of CDK4, Cyclin D1, and EMT biomarkers. Together, increased knowledge of the clinicopathological significance and molecular mechanisms of CDKN2B-AS1 in OS will help us to understand the tumor etiology and could provide a prospective biomarker for diagnosis and therapeutic targets in OS.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.