
Abstract
Introduction:
Growth differentiation factor 15 (GDF15), a newly identified member of transforming growth factor (GDF) superfamily, is upregulated in ovarian (OV) cancer. Upregulated GDF15 positively correlates with poor prognosis of OV cancer. Thus, elucidation of the mechanism underlying GDF15 overexpression is important.
Method and Results:
PROMO and JASPAR prediction software were used to find transcription factors for GDF15 expression. Data from TCGA database were analyzed to find long noncoding RNAs (lncRNAs) that were also abnormally expressed in OV cancer and had associations with GDF15 expression. Transcription factor CEBPB was predicted as an important regulator of GDF15, confirmed by luciferase reporter assay. However, CEBPB expression was not significantly changed in OV cancer. Data from TCGA database showed that lncRNA GAS5 is downregulated in OV cancer and its expression is negatively correlated with GDF15 expression. RPISeq showed high affinity of GAS5 to CEBPB and this was confirmed by RNA-binding protein immunoprecipitation assay. GAS5 overexpression increased its binding to CEBPB and consequently downregulated GDF15. GAS5 overexpression and GDF15 knockdown decreased viability and increased apoptosis of OV cancer cells, but CEBPB overexpression had opposite effects. However, simultaneous GAS5 and CEBPB overexpression or CEBPB overexpression together with GDF15 knockdown had no effect on cell viability and apoptosis.
Conclusion:
GAS5 functions as decoy of CEBPB, blocking transcription-promoting effect of CEBPB on GDF15.
Introduction
Ovarian (OV) cancer is the most lethal malignant tumor in gynecology with the 5-year survival rate at about 30%. 1,2 There is about 20,000 new cases of OV cancer and about 15,000 deaths from OV cancer in the United States in 2018. 2 As the ovary is located deep in the pelvic cavity, OV cancer is commonly imperceptible. Up to 75% of patients with OV cancer are at the advanced stage at first diagnosis. 3 Although cytoreductive surgery and platinum/paclitaxel combination chemotherapy can temporarily relieve the disease, the recurrence rate in the future is as high as 85%, and the drug resistance of recurrent OV cancer is significantly increased. 3 At present, the pathogenesis of OV cancer is not very clear. Elucidation of the pathogenesis is necessary to improve the treatment regimens and the consequent prognosis of the patients.
Growth differentiation factor 15 (GDF15) belongs to the transforming growth factor superfamily. Recently, the role of GDF15 in the process of tumorigenesis and development of OV cancer has gained increased attention. Staff et al., found that levels of GDF15 in the plasma were elevated in the patients with OV cancer as compared with that in healthy controls, and GDF-15 plasma concentration correlated inversely with survival time after correction for FIGO stage and age. 4 Exogenous stimulation of OV cancer cell lines with recombinant human GDF15 or stable overexpression of GDF15 promoted anchorage-independent growth, increased invasion, and upregulation of matrix metalloproteinases and vascular endothelial growth factor. 5 Conversely, knockdown of endogenous GDF15 or neutralization of secreted GDF15 suppressed invasion and growth of a GDF15-overexpressing OV cancer cell line. 5 In addition, GDF15 has been found to be closely related to platinum resistance of OV cancer in clinical setting. 6 These results suggest that aberrantly upregulated GDF15 plays an important role in the proliferation, invasion, and drug resistance of OV cancer. However, the reason underlying the upregulation of GDF15 in OV cancer remains unclear.
Long noncoding RNAs (lncRNAs) are a class of RNA molecules longer than 200 nucleotides that are not translated into protein. Protein-encoded transcripts in human genome account for only 1%–2%, whereas lncRNA transcripts are 4%–9%. 7,8 lncRNA was first considered to be the noise of gene transcription and that it had no biological function. 7,8 However, more and more studies have shown that lncRNA plays an important role in regulating gene expression at the transcriptional level, post-transcriptional level, and epigenetic level in various physiological and pathological processes. 8 One of the well-studied molecular functions of lncRNAs in gene regulation is they act as ceRNA (competing endogenous RNAs), that is, lncRNAs can sponge miRNA to prevent miRNA-mediated degradation of target genes. 9,10 In addition, lncRNAs can function as a decoy of transcription factors to sequester from their site of action and regulate gene expression. For example, lncRNA PANDA (P21-associated ncRNA DNA damage activated) is very sensitive to DNA damage and favors cell cycle arrest by interacting with the transcription factor NF-YA to limit expression of proapoptotic genes. 11,12 Similarly, the NRON ncRNA can bind to the transcription factor nuclear factor of activated T cells rendering it inactive, because it prevents nuclear accumulation. 13
Studies have found that lncRNA GAS5 was abnormally downregulated in OV cancer, and low expression of lncRNA GAS5 facilitates progression of OV cancer. 14 According to previous research, the anticancer effect of lncRNA GAS5 is associated with the ceRNA mechanisms for miR-196-5p and miR-21. 15,16 Moreover, by using multiple bioinformatics analysis, the authors found that lncRNA GAS5 is a potential regulator of GDF15, acting as a decoy of transcription factor CEBPB. This study aimed to identify this novel mechanism of lncRNA GAS5 with respect to cancer suppression.
Materials and Method
Sample collection
OV cancer tissue samples (n = 18) and normal ovary tissue samples (n = 8) were obtained from patients who underwent operation at the First Affiliated Hospital of Jinzhou Medical University between 2017 and 2018. Informed patient consent was obtained before surgical operations. All the OV cancer diagnoses were re-evaluated and defined by at least 2 expert pathologists. Normal ovary tissue samples were from people who underwent contraceptive operation with ovariectomy, or patients who suffered from tubal ectopic pregnancy that needed ovariectomy. These ovary tissues were confirmed to have no OV cancer by pathological analysis. None of the recruited patients was subjected to preoperative radiotherapy or chemotherapy. This study was approved by the Ethics Committee of Xiangya School of Medicine (Changsha, People's Republic of China).
Cell culture and treatments
Human OV carcinoma tumor cell lines, including Caov-3, OVCAR4, OVCAR5, OVCAR8, EFO-27, MCAS, and SK-OV-3, were obtained from the American Type Culture Collection (ATCC, Manassas, VA). These cells were grown in RPMI 1640 medium (Sigma, St. Louis, MO) supplemented with heat-inactivated 5% fetal bovine serum (Invitrogen, Grand Island, NY) and 1% antibiotic/antimycotic (Invitrogen). Cells were maintained in a 5% CO2 atmosphere at 37°C in a humidified tissue culture incubator.
Cell transfection
OVCAR4 cells were seeded onto six-well plates with a density of 1 × 105 cells/well. OVCAR-4 cells were transfected with siRNA-GDF15 constructed by GenePharma (Shanghai, China) to knock down the targeted genes. To epigenetically overexpress lncRNA GAS5 and CEBPB, their overexpression vectors (GenePharma) were transfected in OVCAR-4 cells using Lipofectamine RNAi Max (Thermo Fisher Scientific, Waltham, MA), according to the manufacturer's protocols. The expression levels of lncRNA GAS5, CEBPB, and GDF15 were assessed 48 h after the transfection using polymerase chain reaction (PCR) assay.
PCR assay
Total RNA was extracted using the mirVana miRNA Isolation Kit (Invitrogen). A High-Capacity RNA-to-cDNA Master Mix (Invitrogen) was used to synthesize cDNA. RT-qPCR was performed using the SYBR ExScript RT-PCR Kit (TaKaRa, Dalian, China) on an ABI 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions. The thermocycling profile was as follows: 95°C for 30 s, followed by 40 cycles of 95°C for 5 s, and 60°C for 30 s. The primer sequences are shown in Table 1.
Premiers Used in Polymerase Chain Reaction Assay
Western blot analysis
Frozen tissues and cell samples were homogenized and lysed in RIPA lysis buffer (Beyotime, Shanghai, China) supplemented with phenylmethanesulfonyl fluoride (Beyotime). The protein concentration of the supernatant was quantitated using the BCA Reagent Kit (Beyotime). Equal amounts of the protein samples (20 μg) were separated by electrophoresis on a 10% or 12% sodium dodecyl sulfate–polyacrylamide gel and transferred to a polyvinylidene fluoride membrane (Bio-Rad, CA). After blocking with nonfat dry milk (Yili Milk Company, Inner Mongolia, China) at room temperature for 2 h, the membranes were hybridized with anti-GDF15 antibody (Dilution 1:1000, ab180754; Abcam, Cambridge, United Kingdom) and anti-GAPDH antibody (Dilution 1:5000, SC-365062; Santa Cruz Biotechnology, Santa Cruz, CA) at 4°C overnight. The primary antibodies were visualized by adding secondary biotin-conjugated antibodies followed by an avidin/biotin/peroxidase complex (VECTASTAIN ABC Elite Kit; Vector Laboratories, Inc., Burlingame, CA) and substrate (Vector NovaRED, VECTASTAIN). Quantification was performed by densitometric analysis using Quantity One software (Bio-Rad).
Luciferase reporter assay
Bioinformatics analysis by PROMO (
RNA immunoprecipitation
RNA immunoprecipitation (RIP) was performed on OVCAR-4 cells using the EZ-Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore) to investigate interactions between lncRNA GAS5 and CEBPB. Briefly, HTR8/SVneo cells were lysed with RIPA-2 buffer and then incubated with Protein-A Dynabeads conjugated with IgG or CEBPB antibodies. The bead/antibody/lysate mixture was incubated at 4°C overnight rotating end over end. Beads were washed with cold NT2 buffer five times. Proteinase K treatment released RNAs from bound proteins and bound RNA was isolated with TRIzol (Invitrogen) and reverse transcribed as described above. Enrichment for lncRNA GAS5 with CEBPB immunoprecipitation was calculated using the comparative Ct method, with samples normalized to input and compared with IgG control. Data are presented as fold enrichment relative to GAPDH enrichment for each sample.
Cell viability measurement
OVCAR-4 cells were seeded in triplicate at 1 × 103 cells/well onto 96-well microplates. After the abovementioned cell treatments, 20 μL (at 5 mg/mL) 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT; Sigma) was added to each well. The medium containing MTT was removed after 4 h and 150 μL dimethylsulfoxide was added subsequently to terminate the reaction. A microplate reader was used to examine absorbance of each well of the plate at 570 nm.
Flow cytometry
OVCAR-4 cells were stained using the Annexin V-FITC/PI Apoptosis Detection Kit I (Kaiji Biological, Inc., Shanghai, China) according to the manufacturer's instructions. The rate of apoptosis was analyzed using a dual laser flow cytometer (Becton Dickinson, San Jose, CA) and estimated using the ModFit LT software (Verity Software House, Topsham, ME).
Statistical analysis
Data were analyzed using SPSS 12.0 software (SPSS, Inc., Chicago, IL). The statistical significance of the differences were evaluated by performing one-way analysis of variance followed by the Scheffe's post-hoc test. p < 0.05 was considered to indicate a statistically significant difference.
Results
CEBPB is an important transcription factor probably responsible for GDF15 expression in OV cancer
We initially analyzed the expression of GDF15 expression in 33 kinds of tumor, based on the data in TCGA database. The results were presented as Manhattan Plot in Figure 1A. GDF15 was upregulated in OV cancer, prostate adenocarcinoma, skin cutaneous melanoma, and so on. However, GDF15 was downregulated in acute myeloid leukemia, lung squamous cell carcinoma, and so on. There were also some kinds of cancers in which the expression level of GDF15 was not different from that in the corresponding normal tissues. These data suggested that the upregulation of GDF15 only exist in some types of cancers. To confirm the upregulation of GDF15 in OV cancer as seen in TCGA database (Fig. 1B, p < 0.05), they performed qPCR and western blot assays to test the expression of GDF15 in 18 OV cancer tissue samples and 8 normal ovary tissue samples. In accordance with the data in TCGA database, GDF15 was overexpressed in the OV cancer at both the mRNA (Fig. 1C, p < 0.01) and protein levels (Fig. 1D, p < 0.05).
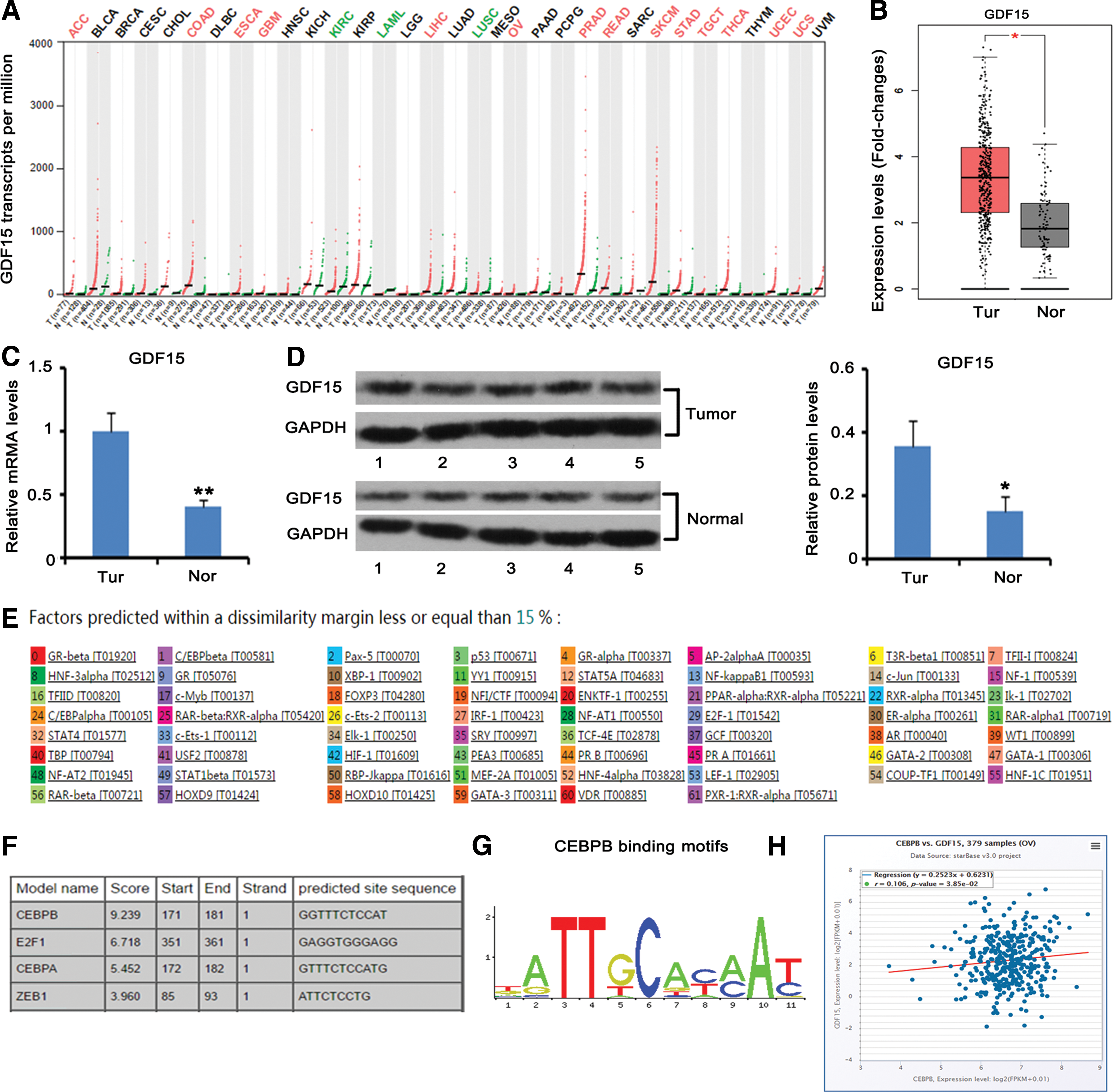
Analysis of transcription factor regulating GDF15 expression in OV cancer.
The authors further analyzed the transcription factors that are responsible for the expression of GDF15. As indicated by the PROMO prediction web-based tool, GDF15 is under the regulation of various transcript factors, such as CEBPB, E2F1, and ZEB1 (Fig. 1E). Another prediction web-based tool, JASPAR, showed that CEBPB had very high score for binding to the promoter region of GDF15 gene (Fig. 1F). Figure 1G shows the specific binding motifs (sequences: TTXCXXXAX, “X” can be G/T, A/T, C/T, A/C, and C/T consecutively) in gene promoters. Furthermore, GDF15 expression was positively correlated to that of CEBPB in OV cancer (p < 0.05), as indicated by available data in TCGA database (Fig. 1H). However, the TCGA database data showed no difference in CEBPB expression between the OV cancer and normal OV tissues (data not shown).
To determine that GDF15 gene is indeed regulated by CEBPB, they performed luciferase reporter assay. Transfection with the CEBPB overexpression vector increased the luciferase activity of the wild-type GDF15 constructs (p < 0.01, Fig. 2A). The core bases of CEBPB-binding sites are “T,” “T,” “C,” and “A,” thus they mutated these bases in the mutant type GDF15 constructs. Results showed that CEBPB overexpression had no effect on the luciferase activity of the mutant type GDF15 constructs. Furthermore, the PCR analysis showed ∼2.5-fold increase in GDF15 expression (p < 0.01, Fig. 2B) with the upregulation of CEBPB (p < 0.01).
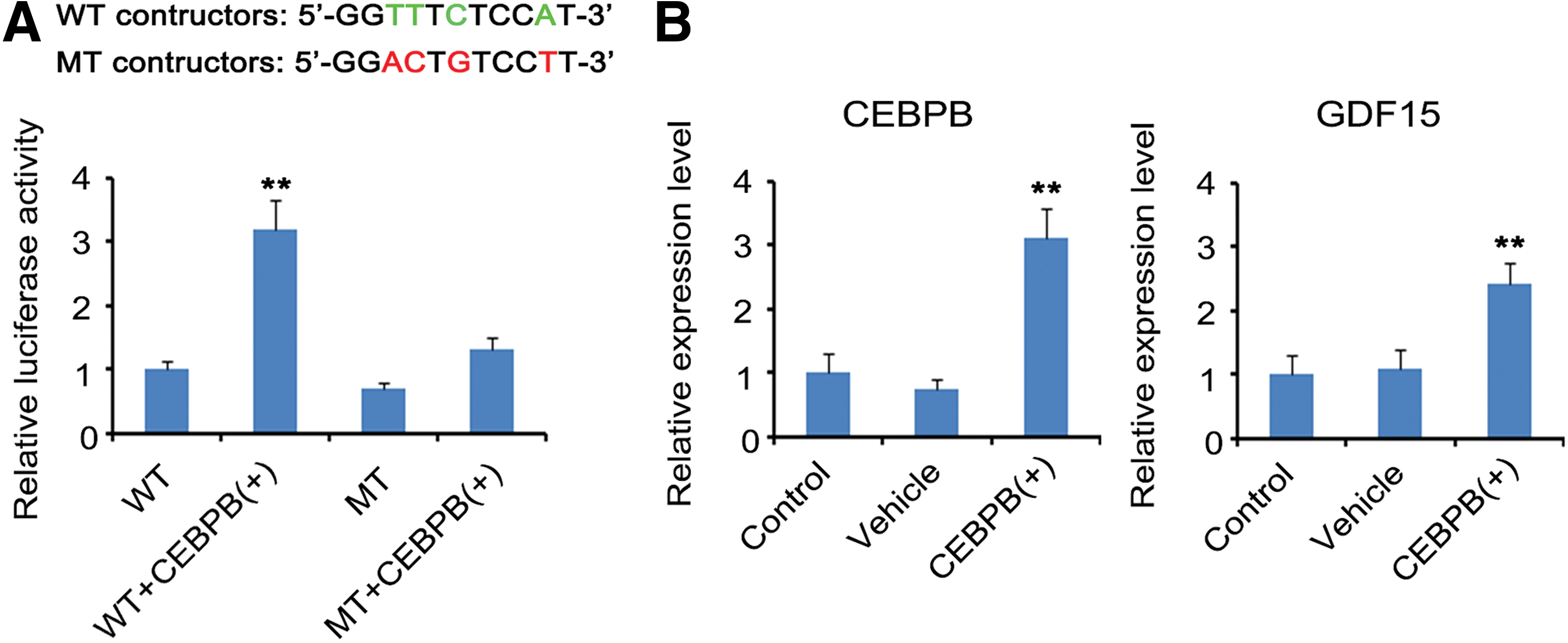
CEBPB positively regulates GDF15 expression in OV cancer cells.
lncRNA-GAS5 expression has negative correlation with that of GDF15 in OV cancer
The authors also analyzed the expression of lncRNA-GAS5 in various types of tumor based on the data in TCGA database. The results are presented as Manhattan Plot in Figure 3A. lncRNA-GAS5 was downregulated in OV cancer and cervical squamous cell carcinoma, but upregulated in many types of cancer, such as lymphoid neoplasm diffuse large B cell lymphoma, glioblastoma multiforme, kidney renal clear cell carcinoma, and so on. To confirm the downregulation of GDF15 in OV cancer in TCGA database (p < 0.05, Fig. 3B), they performed the qPCR assays checking the expression of lncRNA-GAS5 in OV cancer tissue samples and normal ovary tissue samples. Lower expression of lncRNA-GAS5 was observed in OV cancer tissues than that in normal ovary tissues (p < 0.05, Fig. 3C).
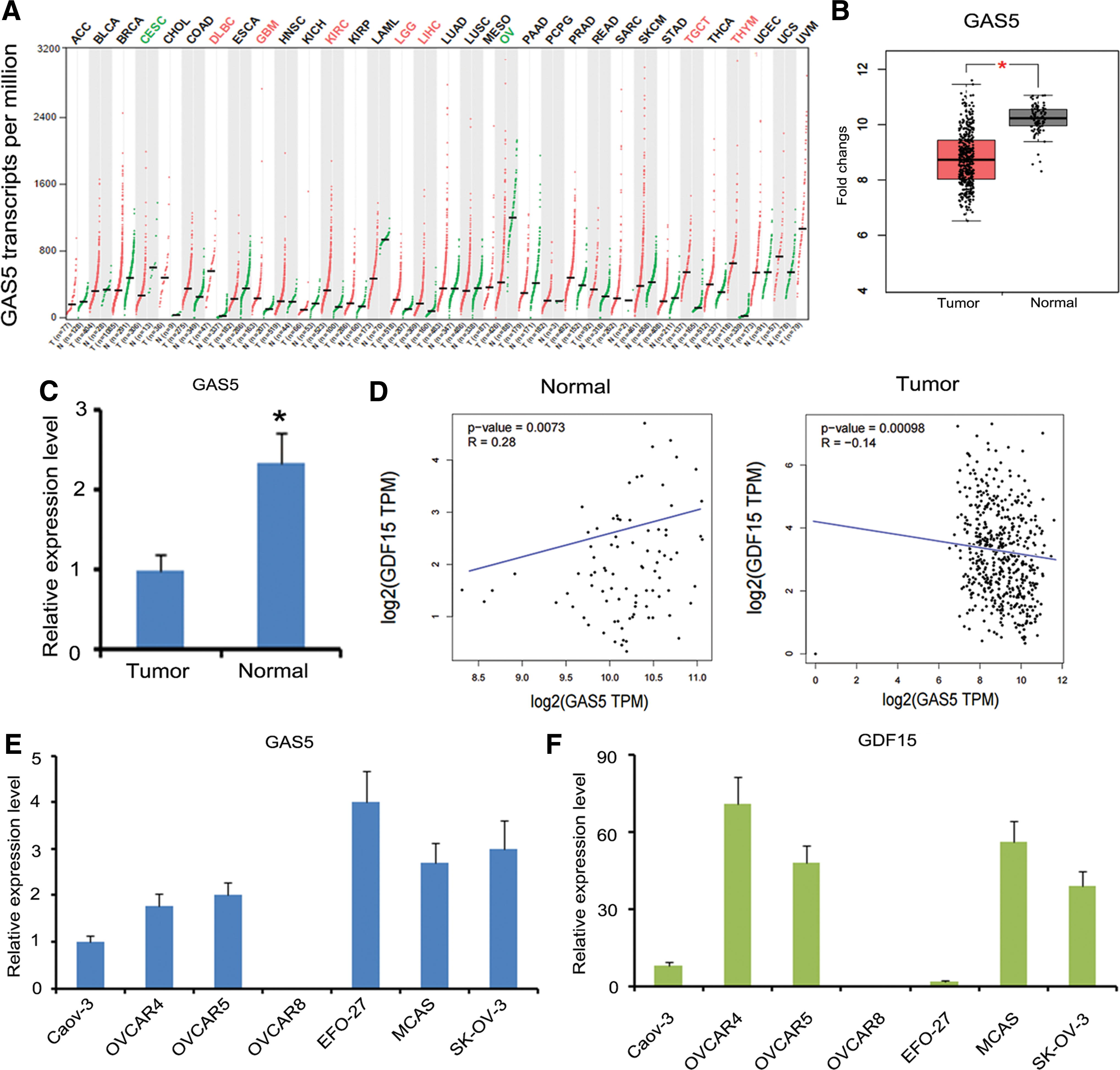
lncRNA-GAS5 expression has negative correlation with that of GDF15 in OV cancer.
This study further analyzed the correlation between lncRNA-GAS5 and GDF15 expression in OV cancer and normal ovary tissues using the data from TCGA database. lncRNA-GAS5 expression had positive correlation with that of GDF15 in normal ovary tissues (p < 0.001, Fig. 3D), however, lncRNA-GAS5 expression was negatively correlated with that of GDF15 in OV cancer tissues (p < 0.001). To further determine the association in their expression, this study detected the expression of lncRNA-GAS5 and GDF15 in seven kinds of OV cancer cells, including Caov-3, OVCAR4, OVCAR5, OVCAR8, EFO-27, MCAS, and SK-OV-3. OVCAR4 expressed relatively low level of lncRNA-GAS5 (Fig. 3E), while showing highest expression of GDF15 among all these cell lines (Fig. 3F). Conversely, EFO-27 cells presented the highest expression of lncRNA-GAS5, but had almost no expression of GDF15. There were some exceptions, like OVCAR8 cells, in which both lncRNA-GAS5 and GDF15 expression were not detected. Based on these data, OVCAR4 cells were used for further study.
The present study further focused on the regulatory effect of lncRNA-GAS5 on GDF15 expression in OV cancer OVCAR-4 cells. The authors epigenetically overexpressed lncRNA-GAS5 in OVCAR-4 cells by transfecting the expression vector. As indicated by PCR assay, lncRNA-GAS5 expression increased by about 2.5-folds 36 h after transfecting the expression vector (p < 0.01, Fig. 4A). The upregulation of lncRNA-GAS5 resulted in the reduction of GDF15 expression in OVCAR-4 cells (p < 0.05, Fig. 4B), but had no impact on the expression of CEBPB.
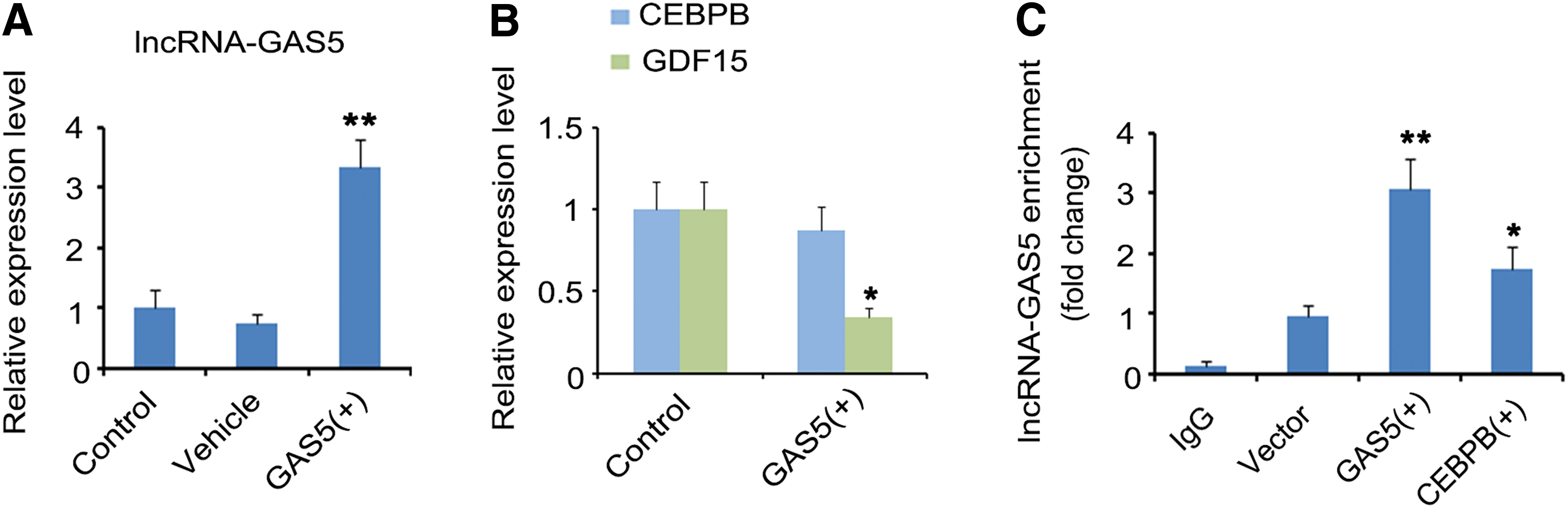
lncRNA-GAS5 negatively regulates GDF15 by binding to CEBPB
Bioinformatics analysis (
The regulatory effect of lncRNA GAS5/CEBPB/GDF15 network on cell proliferation and apoptosis in OV cancer
To determine the regulatory effect of lncRNA GAS5/CEBPB/GDF15 network on cell proliferation and apoptosis in OV cancer, this study artificially changed the expression of lncRNA GAS5, CEBPB, and GDF15 by transfecting corresponding vectors. Transfection with siRNA-GDF15 caused the reduction of GDF15 expression (p < 0.01, Fig. 5A). Overexpression of lncRNA GAS5 and CEBPB decreased and increased GDF15 expression, respectively (p < 0.05, Fig. 5B). Cotransfection with lncRNA GAS5 and CEBPB overexpression vectors had no significant effect on GDF15 expression. CEBPB overexpression together with GDF15 knockdown did not significantly change the GDF15 expression either.
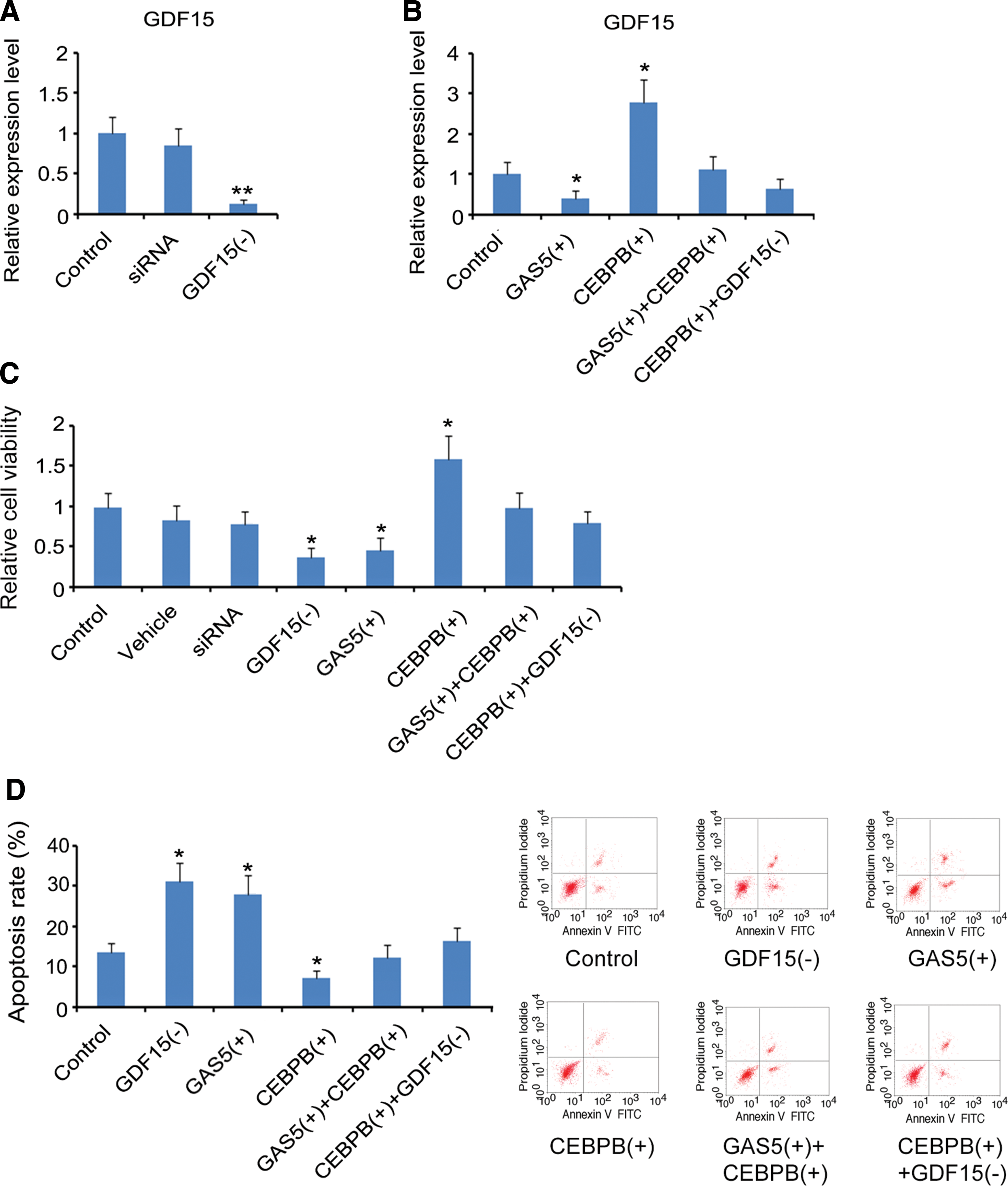
The regulatory effect of lncRNA GAS5/CEBPB/GDF15 network on cell proliferation and apoptosis of OV cancer.
Viability of OVCAR-4 cells was decreased after GDF15 knockdown and lncRNA GAS5 overexpression (p < 0.05, Fig. 5C). However, CEBPB overexpression increased OVCAR-4 cell viability (p < 0.05). Cell viability did not change following the overexpression of both lncRNA GAS5 and CEBPB as well as CEBPB overexpression and GDF15 knockdown. Both GDF15 knockdown and lncRNA GAS5 overexpression increased the apoptosis rate of OVCAR-4 cells (p < 0.05, Fig. 5D). CEBPB overexpression conversely decreased the apoptosis rate (p < 0.05). lncRNA GAS5 overexpression together with CEBPB overexpression as well as CEBPB overexpression with GDF15 knockdown had no effect on apoptosis rate.
We performed protein/protein interaction (PPI) analysis using String (

The potential mechanism underlying the cancer-promoting effect of GDF15.
Discussion
GDF15 is a newly discovered member of transforming growth factor superfamily. Increasing evidence demonstrates that GDF15 is aberrantly upregulated in OV cancer and has a growth-promoting effect. 4 –6 However, the mechanism underlying this upregulation is unclear. Transcription factors play a vital role in the initiation of gene transcription, acting as important regulators in gene expression. PROMO prediction web tool showed that various transcript factors might take part in the regulation of GDF15 gene transcription. The JASPAR prediction web tool marked a very high score in CEBPB among these transcription factors, suggesting the high possibility of involvement of CEBPB in the regulation of GDF15 expression. Moreover, CEBPB expression has positive correlation with GDF15 expression as indicated by the data from TCGA database. Therefore, this study focused on the research of the regulatory effect of CEBPB on GDF15 expression. Luciferase reporter assay identified the binding of CEBPB to GDF15 gene promoter and positive regulation of CEBPB in GDF15 gene expression. Epigenetically, overexpressing CEBPB caused increase in GDF15 expression in OV cancer cells. All these data suggested that CEBPB exerted promoting effect on GDF15 expression in OV cancer cells.
CEBPB belongs to the CCAAT/enhancer-binding protein (CEBP) family. CEBPB protein is not essential for the physiological function of mouse ovary, because it is expressed in a highly restricted temporal and spatial pattern in the mouse ovary and the deletion of CEBPB in mice had no overt effect on OV physiology and reproductive functions. 17 However, higher expression of CEBPB was observed in chemoresistant OV cancer cells than in chemosensitive OV cancer cells. CEBPB is involved in the chemoresistance of OV cancer by regulating COL11A1 expression. 18 Their study detected the expression of CEBPB in normal and cancer tissues of OV using PCR. However, they did not find significant difference in CEBPB expression between the normal and cancer tissues. These data are in accordance to those in TCGA database. Since the expression of CEBPB in OV cancer was not changed, it is likely that the transcription-regulatory effect of CEBPB on GDF15 is enhanced by other molecular mechanisms.
lncRNAs play an important role in regulating gene expression at the transcriptional level, because they can function as decoy transcription factors to sequester from their site of action to regulate gene expression. Therefore, downregulation of lncRNA might, in turn, increase the binding of transcription factors to gene promoter, thus enhancing the function of transcription factors. lncRNA GAS5 has been found be downregulated in OV cancer. 14 Li et al. further found that decreased expression of lncRNA GAS5 promotes cell proliferation, migration, and invasion of OV cancer cells, and is associated with poor prognosis. 14 This study found a negative correlation between lncRNA GAS5 and GDF15 expression in OV cancer through analysis of data from TCGA database and high affinity between lncRNA GAS5 and CEBPB. Therefore, they hypothesized that lncRNA GAS5 was competitively binding to CEBPB with GDF15 gene promoter, thereby attenuating the transcription efficiency of CEBPB. In line with the hypothesis, study through immunocoprecipitation showed that lncRNA GAS5 overexpression increased the amount of lncRNA GAS5 attaching to CEBPB. Moreover, lncRNA GAS5 overexpression decreased GDF15 expression, but lncRNA GAS5 overexpression together with CEBPB overexpression had no effect on GDF15 expression.
TCGA database showed that lncRNA-GAS5 expression had negative correlation with that of GDF15 in OV cancer tissues. The p-value of the correlation is less than 0.001, yet the r value is no more than 0.3. They hypothesized that the expression of GDF15 in OV cancer may also be regulated by other mechanisms in addition to the mechanism reported in their study. For example, previous study suggested that GDF15 expression in cancer is influenced by some types of miRNAs. 19 But, given the p-value of the correlation is very remarkable, GAS5 probably exerted the predominant effect among all the mechanisms underlying the regulation of GDF15 expression. An interesting finding in this study is expression of GDF15 is not elevated in some OV cell lines, such as OVCAR8. OVCAR8 cell line was derived from patients previously treated with intensive chemotherapy. 20 Chemotherapy likely changed the original gene expression profile of OV cancer tissues. As OV cell lines were obtained from patients with different treatment backgrounds, these cell lines show various gene expression profiles.
This study confirmed that lncRNA GAS5/CEBPB/GDF15 regulatory axis is involved in the regulation of proliferation and apoptosis of OV cancer cells. A study performed in cervical cancer cells showed that GDF15 binds to ErbB2 receptor activating the downstream targets such as AKT1 and Erk1/2. This molecular mechanism promotes the proliferation of cervical cancer. 21 In addition, GDF15 has been confirmed to bind to TGF-β receptor to activate Smad2 and Smad3 pathways resulting in the epithelial/mesenchymal transition of colorectal cancer. 22 Their study revealed that GDF15 could bind to many kinds of proteins. Some of them play key roles in the generation and development of tumor, such as p53, CXCL8, EGF, and EGR2. Their study further identified that GDF15 impacts the functions of these proteins, thus promoting OV cancer progression. In addition, bioinformatics analysis showed that GDF15 is most likely involved in the biological processes, including myoblast fusion, cell development, MAPK, SMAD, BMP, and transforming growth factor receptor cascades. Most of these processes are important for cancer growth and metastasis.
To conclude, this study revealed an important mechanism underlying the abnormal upregulation of GDF15. This mechanism was associated with the downregulation of lncRNA GAS5, which increased the amount of CEBPB binding to the promoter region of GDF15, thus elevating the transcriptional efficiency. Consequently, lncRNA GAS5/CEBPB/GDF15 regulatory axis has been confirmed to be associated with the regulation of proliferation and apoptosis of OV cancer cell.
Footnotes
Disclosure Statement
No competing financial interests exist.