
Abstract
Background:
LncRNA LINC00974 participates in oral fibrogenesis, indicating possible involvement in other oral diseases.
Results:
The authors found that LINC00974 was upregulated in oral squamous cell carcinoma (OSCC) and predicted poor survival. In OSCC tissues, LINC00974 was inversely correlated with miR-122 and positively correlated with RhoA. In OSCC cells, LINC00974 overexpression resulted in upregulated, whereas overexpression of miR-122 led to downregulated RhoA. Moreover, downregulated miR-122 was observed after LINC00974 overexpression, whereas LINC00974 expression was not significantly affected by miR-122 overexpression. In invasion and migration assay, miR-122 overexpression resulted in reduced, whereas LINC00974 and RhoA overexpression resulted in increased rate of cancer cell migration and invasion. In addition, miR-122 overexpression reduced the effects of LINC00974 overexpression.
Conclusion:
Therefore, LINC00974 can downregulate miR-122 to upregulate RhoA in OSCC, thereby promoting cell invasion and migration.
Introduction
Incidence of oral squamous cell carcinoma (OSCC) ranks eighth among all malignancies. 1 It is estimated that only <60% patients with OSCC live longer than 5 years after the initial diagnosis. 2 Therefore, OSCC is still a major threat of human health. OSCC mostly originates from thel tongue, which is also the subsite with the worst of prognosis. 3 In recent years, incidence of OSCC showed a decreasing trend but the onset age is also becoming younger. 4 Betel nut chewing, tobacco smoking, and alcohol consumption are the main risk factors for OSCC. 5 However, pathogenesis of this disease is still relatively unknown. 6
RhoA, also known as Ras homolog gene family member A, is a small GTPase with pivotal roles in actomyosin contractility and actin stress fiber formation. 7 RhoA is overexpressed in several types of cancers and promotes cancer cell invasion and migration, thereby promoting cancer metastasis. 8 Some tumor-suppressive miRNAs, such as miR-122, can target RhoA to suppress cancer progression. 9 MiR-122 can interact with transforming growth factor (TGF)-β in many biological processes. 10 In a recent study, Fang et al. reported an lncRNA named LINC00974 with oncogenic role in oral fibrogenesis, which is closely related to OSCC. 11 In oral fibrogenesis, LINC00974 activates TGF-β signaling to promote disease progression. 11 Therefore, the authors speculate that LINC00974 may also have interactions with miR-122 and RhoA in OSCC. This study aimed to explore this possibility.
Materials and Methods
OSCC patients
This study passed review by the Review Board of Ethics Committee of The First Affiliated of Baotou Medical College (#ECBMC20111121357). The present study selected 66 OSCC patients (gender: 30 males and 36 females; age: 32–71 years; mean age: 49.3 ± 5.8 years) from the 122 OSCC patients admitted to and treated in The First Affiliated of Baotou Medical College between January 2012 and January 2014. Inclusion criteria: (1) newly diagnosed OSCC cases; (2) therapies were not initiated. Exclusion criteria: (1) other clinical disorders were also diagnosed; (2) recurrent OSCC; (3) therapies performed. All patients were staged based on AJCC system and clinical findings. The 66 patients included 12, 24, and 30 patients at clinical stage II–IV, respectively. All patients were informed with the principle of the study. All 66 patients signed informed consent.
OSCC tissue samples
Biopsy was performed on all OSCC patients before the initiation of therapies. During biopsy, adjacent nontumor (<1% cancer cells) and OSCC (more than 90% cancer cells) tissue samples were obtained from each patient. All samples were kept in a liquid nitrogen tank.
OSCC cells
SCC090 human OSCC cell line (ATCC) was used in this study. Eagle's minimum essential medium (2 mM
Transient transfections
Negative control (NC) miRNA and miR-122 mimic were synthesized by Sangon (Shanghai, China). LINC00974 and RhoA expression vectors were constructed using pcDNA3.1 vector (Sangon). SCC090 cells were harvested at confluence of 70%–80% and were counted. Lipofectamine 2000 (Sangon) was used to transfect 40 nM miRNA (NC miRNA as NC group) or 12 nM vector (empty pcDNA3.1 vector as NC group) into 5 × 106 cells. Untransfected cells were control (C) cells. All subsequent experiments were performed at 24 h post-transfection.
RNA extractions
Total RNAs in 5 × 105 SCC090 cells of each transfection group were extracted using RNAzol (Sigma-Aldrich). All steps were completed following the instructions from Sigma-Aldrich. To harvest miRNAs, 80% ethanol was used to precipitate and wash RNA samples.
Quantitative polymerase chain reactions
After RNA extractions, DNase I was used to digest RNA samples at 37°C for 2 h to completely remove genomic DNA. AMV Reverse Transcriptase (Promega Corporation) was used to perform reverse transcriptions (RTs) with RNA samples as template. To measure the expression levels of LINC00974 and RhoA mRNA, the QuantiTect SYBR Green PCR Kit (Qiagen) was used to prepare quantitative polymerase chain reaction (qPCR) mixtures with GAPDH as endogenous control.
To measure the expression levels of miR-122, the All-in-One™ miRNA qRT-PCR Detection Kit (Genecopoeia) was used to perform 3′-polyadenylation, RTs, and qPCR reaction mixture preparation. U6 was used as the endogenous control.
All experiments were repeated three times. 2−ΔΔCT was used for data process.
Western blot
Total proteins in 5 × 105 SCC090 cells of each transfection group were extracted using RIPA solution (Sigma-Aldrich). All RNA samples were quantified using the BCA Kit (Sigma-Aldrich). Electrophoresis (10% sodium dodecyl sulfate/polyacrylamide gel electrophoresis gel) was performed to separate protein after denaturing in boiling water for 6 min. After that, gel transfer to polyvinylidene fluoride membranes was performed, followed by blocking in phosphate-buffered saline containing 5% nonfat milk. Following that, membranes were first blotted with rabbit GAPDH (1: 800, ab37168; Abcam) and RhoA (1: 800, ab86297; Abcam) primary antibodies at 4°C for 18 h, followed by blotting with goat horseradish peroxidase (immunoglobulin G) (1:1000, ab6721; Abcam) secondary antibody at 24°C for 2 h. Membranes were finally incubated with ECL Western Blotting Substrate (ab65623; Abcam) to develop signals, which were processed using ImageJ v1.48 software.
Transwell assays
Transwell assays were performed to analyze the effects of the overexpression of LINC00974, miR-122, and RhoA on the invasion and migration of SCC090 cells. SCC090 cells were counted and 3 × 104 cells were mixed with Eagle's minimum essential medium (0.1% FBS) to prepare single cell suspensions. A 96-well Transwell plate was filled with cell suspension (0.l mL) in the upper chamber and Eagle's minimum essential medium (20% FBS) in the lower chamber. It is worth noting that membranes were coated with Matrigel (356234; Millipore) for 6 h at 37°C before invasion assay. The plate was cultivated at 37°C for 12 h, followed by membrane staining for 20 min with 0.5% Crystal Violet (Sigma-Aldrich). Stained cells were observed under an optical microscope.
Statistical analyses
Mean values of three biological replicates of each experiment were calculated and used in all data analyses. Correlations were analyzed by linear regression. Differences between nontumor and OSCC tissues were analyzed by paired t test. Differences among different cell transfection groups were analyzed by one-way analysis of variance and Tukey test. The 66 OSCC patients were divided into high and low LINC00974-level groups (n = 32 and 34, respectively) according to Youden's index. Survival curves were plotted using K–M plotter and were compared by log-rank test. p < 0.05 was statistically significant.
Results
LINC00974 was upregulated in OSCC and predicted poor survival
Expression levels of LINC00974 in both nontumor and OSCC tissues were measured by qPCR and compared by paired t test. Compared with nontumor tissues, expression levels of LINC00974 were significantly higher in OSCC tissues (Fig. 1A, p < 0.05). Survival curves were plotted and compared through aforementioned methods. Compared with patients in low LINC00974-level group, overall survival rate of patients in high LINC00974-level group was significantly lower (Fig. 1B).
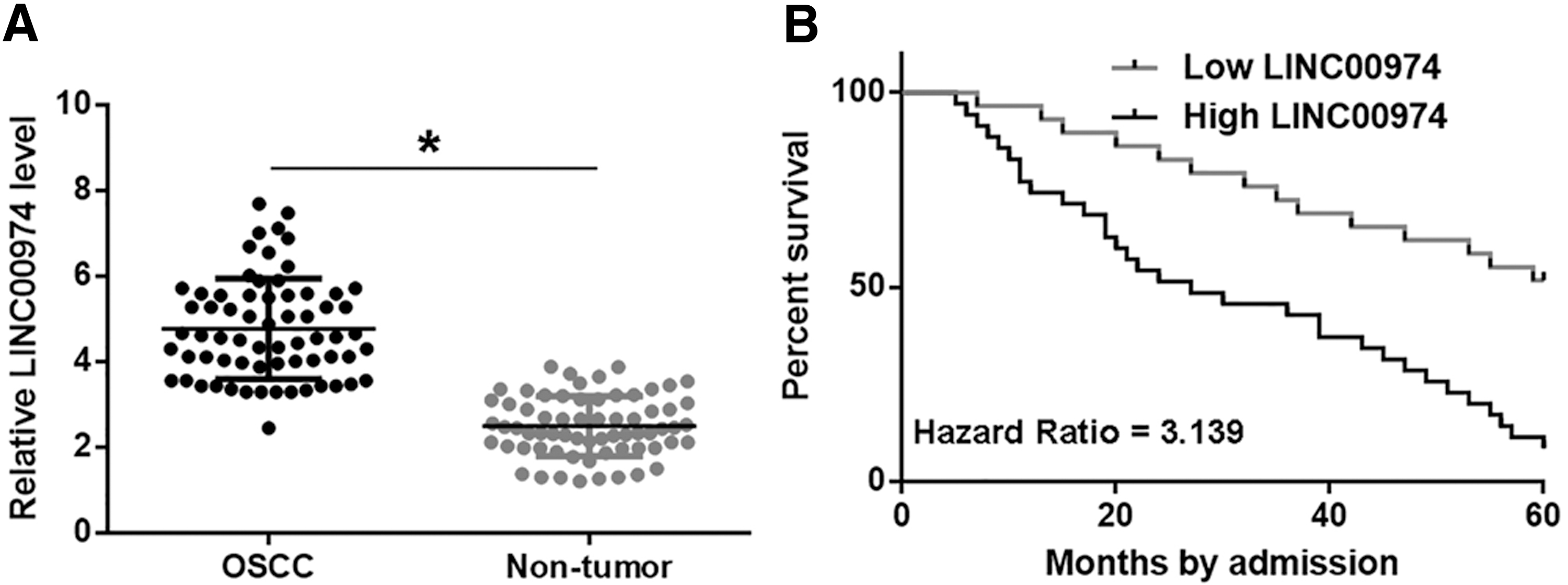
LINC00974 was upregulated in OSCC and predicted poor survival. Expression levels of LINC00974 in both nontumor and OSCC tissues were measured by qPCR and compared by paired t test
LINC00974 was correlated with miR-122 and RhoA in OSCC
Expression levels of miR-122 and RhoA mRNA were also measured by performing qPCR. Correlations between LINC00974 and miR-122/RhoA were analyzed by linear regression. It can be observed that expression level of LINC00974 was significantly and inversely correlated with that of miR-122 (Fig. 2A). However, LINC00974 and RhoA were significantly and positively correlated (Fig. 2B).
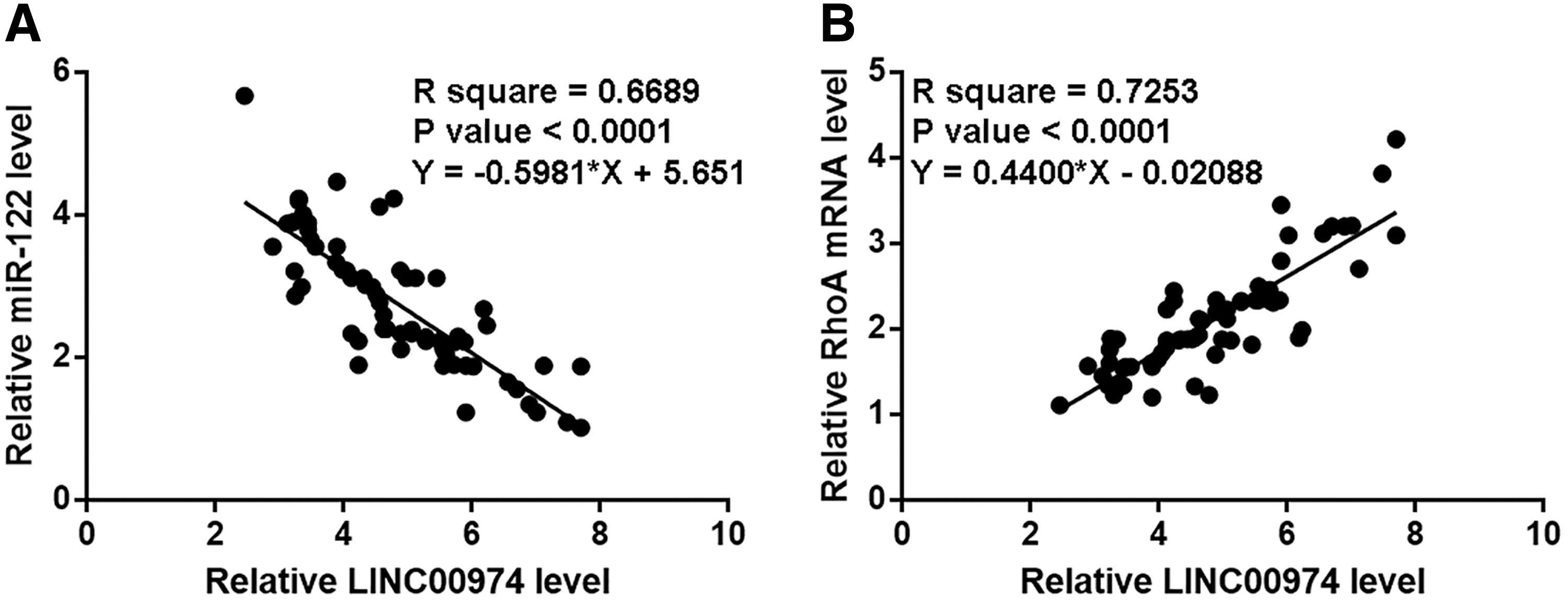
LINC00974 was correlated with miR-122 and RhoA in OSCC. Expression levels of miR-122 and RhoA mRNA were also measured by performing qPCR. Correlations between LINC00974 and miR-122
LINC00974 downregulated miR-122 to upregulate RhoA in SCC090 cells
Expression levels of LINC00974, miR-122, and RhoA were measured at 24 h post-transfections. Compared with NC and C groups, expression levels of LINC00974, miR-122, and RhoA were significantly increased after the transfection of expression vectors or miRNA mimic (Fig. 3A, p < 0.05). Compared with two controls, LINC00974 overexpression resulted in upregulated, whereas miR-122 overexpression resulted in downregulated RhoA, and miR-122 overexpression reduced the effects of LINC00974 overexpression (Fig. 3B, p < 0.05). Moreover, LINC00974 overexpression resulted in downregulated miR-122, whereas LINC00974 expression was not significantly affected by miR-122 overexpression (Fig. 3C, p < 0.05).
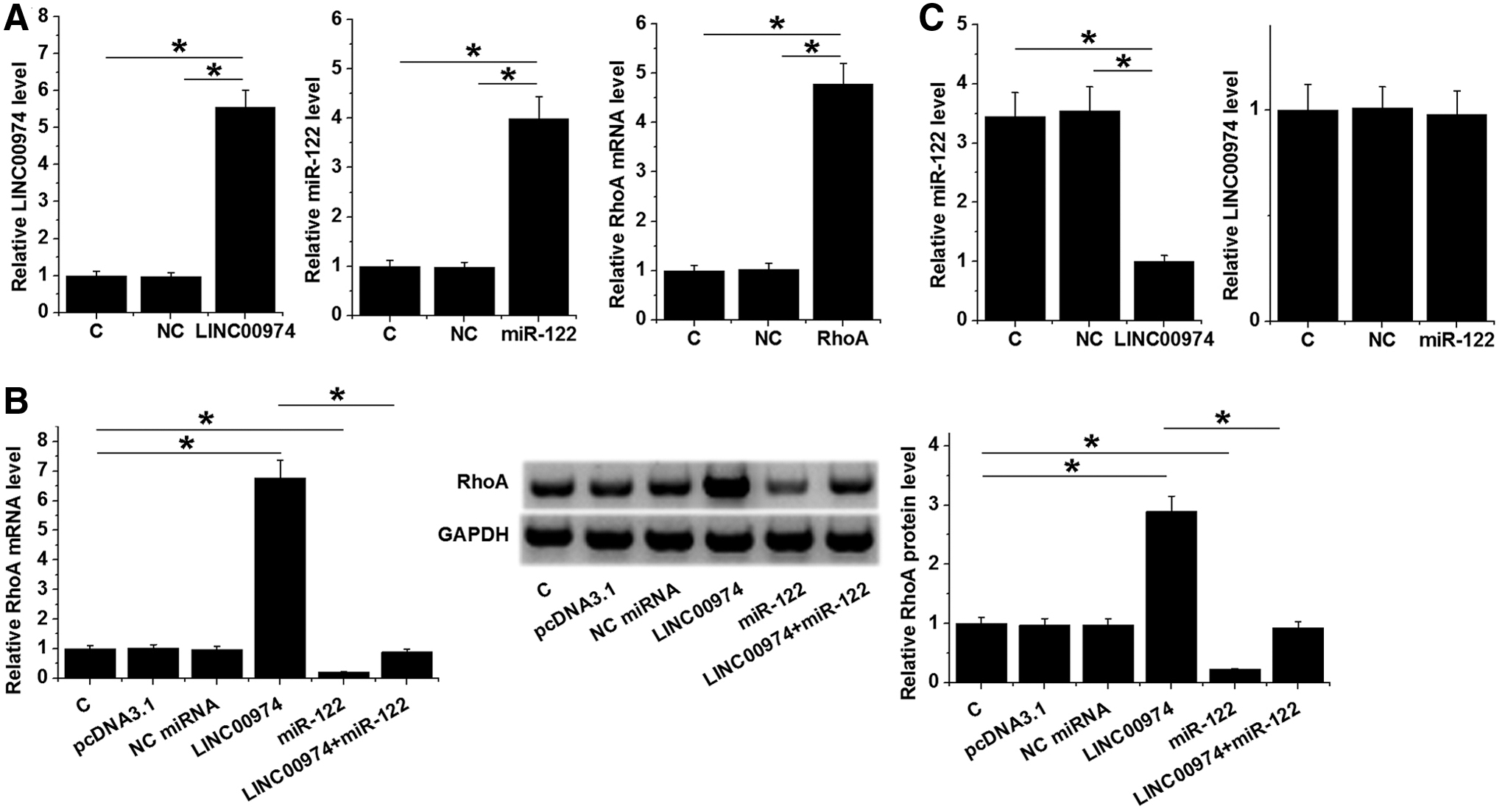
LINC00974 downregulated miR-122 to upregulate RhoA in SCC090 cells. Overexpression of LINC00974, miR-122, and RhoA was confirmed by qPCR at 24h after the transfections of expression vectors or miRNA mimic
LINC00974 promoted SCC090 cell invasion and migration through miR-122 and RhoA
Transwell assays were performed to analyze the effects of the overexpression of LINC00974, miR-122, and RhoA on the invasion and migration of SCC090 cells. Compared with C and NC two control, miR-122 overexpression resulted in reduced, whereas LINC00974 and RhoA overexpression resulted in increased rate of cancer cell invasion (Fig. 4A, p < 0.05) and migration (Fig. 4B, p < 0.05). In addition, miR-122 overexpression reduced the effects of LINC00974 overexpression (p < 0.05).

LINC00974 promoted SCC090 cell invasion and migration through miR-122 and RhoA. Transwell assays were performed to analyze the effects of the overexpression of LINC00974, miR-122, and RhoA on the invasion
Discussion
This study mainly investigated the functions of LINC00974 in OSCC. The authors found that LINC00974 was upregulated in OSCC and predicted the poor survival of OSCC patients. In addition, LINC00974 may downregulate miR-122 to upregulate RhoA, which in turn promote the invasion and migration of OSCC cells.
The expression pattern and functionality of LINC00974 have only been investigated in oral fibrosis and hepatocellular carcinoma (HCC). 11 –13 In oral fibrosis, LINC00974 is overexpressed and promotes disease progression by activating transforming growth factor (TGF)-β/Smad signaling, 11 and inhibition of LINC00974 by arctigenin led to reduced myofibroblast activities, thereby promoting the recovery of oralfibrosis. 12 LINC00974 is also overexpressed in HCC, and it can interact with KRT19 to promote the metastasis and proliferation of HCC cells. 13 This study is the first to report the upregulation of LINC00974 in OSCC. Besides that they also observed the increased rates of OSCC cell invasion and migration after the overexpression of LINC00974. Therefore, their data suggest that LINC00974 is an oncogenic lncRNA in OSCC.
MiR-122 is a well-characterized tumor-suppressive miRNA in many types of cancers, such as breast cancer and HCC. 14,15 However, its involvement in OSCC is unclear. In this study,they found that miR-122 is downregulated in OSCC and it can inhibit the invasion and migration of OSCC cells. Therefore, miR-122 is also likely a tumor-suppressive miRNA in OSCC. It is known that in HCC miR-122 can downregulate RhoA to suppress cancer cell invasion and migration. 9 In the present study they also observed downregulated RhoA after miR-122 overexpression. Therefore, miR-122 may also target RhoA in OSCC.
It is known that the expression of miR-122 can be regulated by certain lncRNAs, such as HULC and ANRIL. 16,17 In this study, they found that LINC00974 can downregulate miR-122 in OSCC to upregulate RhoA, whereas the mechanism is unclear. However, it is known that both LINC00974 and miR-122 have crosstalk with TGF-β. Therefore, TGF-β may mediate the interaction between them.
RhoA as a small GTPase can regulate different cellular processes, such as actomyosin contractility and formation of actin stress fibers. 18 Besides that, RhoA acts on ROCK kinases to induce cell migration and detachment. 19 Consistently, their study also reported the enhancing effects of RhoA on cancer cell invasion and migration. The downstream effectors of RhoA have been well characterized. However, based on their knowledge, the upstream regulation of RhoA has not been well investigated. Their study characterized a novel LINC00974/miR-122/RhoA pathway. Their data provided new insights to the pathogenesis of OSCC. It is worth noting that this study failed to perform in vivo experiments. Future studies are needed to perform animal model experiments to further validate the conclusions in this study.
In conclusion, LINC00974 was upregulated in OSCC and can downregulate miR-122 to upregulate RhoA, thereby promoting cancer cell invasion and migration.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This study is supported by Natural Science Foundation of Inner Mongolia Autonomous Region (Grant No. 2017MS0855).