
Abstract
Objective:
lncRNA HAND2 antisense RNA 1 (HAND2-AS1) is consistently well recognized to suppress multiple tumors, while its function was uncertified in liver cancer.
Materials and Methods:
qRT-PCR analysis and TCGA database discovered the expression in liver cancer. CCK-8 and Transwell migration assay demonstrated the impact of HAND2-AS1 on cell proliferation and migration. Bioinformatic analysis and luciferase reporter assay were utilized to monitor the binding between HAND2-AS1 or SOCS5 mRNA and miR-3118. The function of SOCS5 on inactivating the JAK-STAT pathway was confirmed through Western blot assays. Rescue experiments unmasked that HAND2-AS1-mediated SOCS5 affected cell proliferation and migration through the JAK-STAT pathway in liver cancer.
Results:
The authors discovered the downregulated HAND2-AS1 in liver cancer cells. HAND2-AS1 augmentation apparently impaired the capacity of liver cancer viability, proliferation, and migration. Cytoplasmic HAND2-AS1 directly bound to miR-3118 and released SOCS5, leading to upregulation of SOCS5. Next, the negative regulator role of SOCS5 in the adjusting JAK-STAT pathway was reconfirmed in this study.
Conclusions:
HAND2-AS1 enhanced inactivation of the JAK-STAT pathway through sponging miR-3118 and facilitating SOCS5 to retard cell proliferation and migration in liver cancer.
Introduction
Based on the 2018 Global Cancer Statistics, a larger proportion of diagnosed liver cancer patients are sentenced to death, with the death rate of China remaining alarmingly on top. 1,2 Hepatocellular carcinoma (HCC) characterized by higher aggressiveness and high incidence belongs to liver cancer and ranks as the second prominent cause contributing to tumor-induced death. 3,4 Besides, patients are also in imminent danger of a devastatingly poor, 5-year survival rate. 5 Emphasis placed on unraveling the regulatory mechanism of hepatocarcinogenesis might be conducive to shifting dismal clinical outcomes to favorable ones in liver cancer patients.
Similar to protein-coding genes functioning in tumor initiation and progression, 6 long noncoding RNAs (lncRNAs), defined as greater than 200 nucleotide transcripts, are also qualified as key tumor drivers or inhibitors in the process of cancer development. 7 –9 The instrumental role of lncRNAs in development of liver cancer has been abundantly documented. 10 For instance, MIR210HG predicts poor prognosis and functions as an oncogenic lncRNA in HCC. 11 lncRNA DLX6-AS1 promotes liver cancer by increasing the expression of WEE1 through targeting miR-424-5p. 12 It was obviously concluded that previous research uncovered plenty of oncogenic lncRNAs in liver cancer; however, documents about identified tumor-inhibiting lncRNAs such as LINC01554, 13 LINC01093, 14 and GAS5 15 are limited. In addition, evidence is mounting in recent investigations that lncRNA HAND2 antisense RNA 1 (HAND2-AS1) dominantly impeded varieties of tumors. For example, HAND2-AS1 inhibits esophagus squamous cell carcinoma, 16 osteosarcoma progression, 17 colorectal cancer, 18 and endometrioid endometrial carcinoma. 19 Nevertheless, its role and corresponding mechanism have not been illuminated in liver cancer.
Evidence is accumulating that the Janus kinase/signal transducer and activator of transcription (JAK-STAT) pathway is a master mediator and frequently hyperactivated in tumor promotion, including liver cancer. 20 –23 However, oncogenic signal transduction to various downstream cytokines could also be blocked by a suppressor of the cytokine signaling family such as SOCS5 and SOCS6. 24,25 It is evidently presented that SOCS5 inactivates JAK2/STAT3 pathways to work as a tumor suppressor in human cancers. 24,26 Besides that SOCS5-obstructed liver cancer tumorigenesis was unveiled. 27 However, the current investigation on lncRNA-mediated SOCS5 as the suppressor of JAK2/STAT3 pathways is limited.
Nonprotein-translating RNAs were inclusive of microRNAs (miRNAs) and lncRNAs. Normally, miRNAs represent transcript subtypes with no longer than 25 nucleotides, which is unlike lncRNAs. Targeting mRNAs are demonstrated under the post-transcriptional suppression of miRNAs through binding the mRNA 3’ untranslated region, functioning in cellular biological processes. 28 Moreover, evidence has emerged that lncRNAs endogenously compete against miRNAs to participate in downstream gene regulation. For example, lncRNA AFAP1-AS1 competed with miR-423-5p to accelerate nasopharyngeal carcinoma metastasis through the Rho/Rac pathway. 29 lncRNA LINC00511 competed against miR-185-3p to contribute to breast cancer tumorigenesis through the E2F1/Nanog axis. 30
The present study is designed for unraveling the role of HAND2-AS1 in liver cancer and therefore may potentiate the discovery of new therapies for liver cancer.
Materials and Methods
Identification of differentially expressed mRNAs
Differentially expressed mRNAs were also discovered with the usage of mRNA microarrays in HAND2-AS1-overexpressed Huh-7 cells in comparison with cells transfected with control plasmids. Significantly changed mRNAs between HAND2-AS1-overexpressed and -control groups were identified by the Limma package in R with the aid of the t-test. Fold change >2 and p < 0.05 were used as the standards for differential expression.
Cell lines and culture
Normal liver cell line L02 and cancerous cell lines (Hep3B, BEL-7402, HepG2, and Huh-7), obtained from the Cell Bank of Chinese Academy of Sciences (Shanghai, China), were involved and cultivated as recommended by the manufacturer's protocols. Conventionally, cells were recommended to be incubated at 37°C under a humid incubator with 5% CO2. Dulbecco modified Eagle's medium (DMEM) (Gibco/Brl, Grand Island, NY) was necessary for related cell culture, which also contained 10% fetal bovine serum (10% FBS), 50 U/mL penicillin, and 50 μg/mL streptomycin.
Cell transfection
sh-SOCS5 and relative controls, sh-NC, designed by RiboBio (Guangzhou, China) were applied for SOCS5 downregulation in liver cancer cells. Full length of HAND2-AS1 was inserted into the pcDNA3.1 vector (Invitrogen, Shanghai, China) for the purpose of HAND2-AS1 augmentation with an empty vector as a negative control. MiR-3118 mimics/inhibitor and NC mimics/inhibitor were commercially acquired from Shanghai GenePharma Co., Ltd. (Shanghai, China). sh-HAND2-AS1#1, sh-HAND2-AS1#2, and sh-NC (RiboBio, Guangzhou, China) were utilized for HAND2-AS1 suppression in L02 cells. Twenty-four hours after being planted in the six-well plates, cells were transfected with plasmids using the transfection reagent Lipofectamine 3000 (Invitrogen, Carlsbad, CA) as per manufacturer's instructions.
Real-time quantitative PCR
The expression profiles of HAND2-AS1, SOCS5, and STAT3 mRNA were measured by qRT-PCR analysis and normalized to GAPDH expression. First, the one-step method of total RNA extraction was implemented by applying TRIzol reagent (Invitrogen). Reverse transcriptions of HAND2-AS1 RNA and SOCS5 mRNA into cDNA were carried out following recommendations of the PrimeScript RT reagent (TaKaRa, Otsu, Shiga, Japan) reverse transcription kit. An Mx3000P system and SYBR Green qRT-PCR kit (Biomics, Nantong, China), in accordance with the manufacturers' protocols, were required for all qRT-PCR reactions. The 2-ΔΔCt method was used for calculation of relative changes. GAPDH and U6 were used as reference genes. All assays were repeated three times. The forward primer of SOCS5 is 5′-ATAAGTGGAGATGGTTCTGC-3′ and its reverse primer is 5′-TCCTCCTGTGCAGAGTCC-3′. The forward primer of HAND2-AS1 is 5′-GGGTGTTTACGTAGACCAGAACC-3′ and its reverse primer is 5′-CTTCCAAAAGCCTTCTGCCTTAG-3′. The forward primer of GAPDH is 5′-GTCGATGGCTAGTCGTAGCATCGAT-3′ and its reverse primer is 5′-TGCTAGCTGGCATGCCCGATCGATC-3′.
Western blot analysis
The total cellular protein isolation of liver cancer cells and normal liver cells was executed with RIPA lysis buffer with the protease inhibitor included. The extracted cellular protein was first electrophoresed on 10% sodium dodecyl sulfate–polyacrylamide gel for protein separation, and then transferred to a polyvinylidene fluoride membrane. Next, membrane blotting after 5% skimmed milk blocking was conducted with anti-SOCS5, anti-STAT3, anti-p-STAT3, anti-c-Myc, and anti-PIM1 antibodies, anti-cleaved caspase 3, anti-BCL-2, anti-total caspase 3, anti-BAX, or anti-GAPDH (Abcam, Cambridge, MA) overnight, following which incubation with secondary antibody for 1h was completed and these immunoreactive bands underwent enhanced chemiluminescence exposure and ImageJ software (NIH, Bethesda, MD) analysis. All protein levels were conventionally normalized to GAPDH.
Cell counting kit-8 assay
The examination of proliferation potential of Hep3B and Huh-7 cells together with L02 cells was completed using the cell counting kit-8 (Beyotime, Beijing, China). Suspended 2000 cells were planted into one pore of a 96-well plate for incubation at 24, 48, 72, and 96 h, followed by the addition of 10 μL of cell counting kit-8 (CCK-8) solution into the well, and additionally incubated at 37°C for 1 h. Cell viability of transfected liver cancer cells was evaluated by absorbance at 450 nm recorded by a microplate reader (Thermo Fisher Scientific, Waltham, MA).
Colony formation assay
Twelve-well plates were used for seeding cells (1 × 103 cells/well) following transfection. Every 3 d, the authors replaced the culture medium and 2 weeks later, cells were washed gently by phosphate-buffered saline and underwent 10 min of fixation with methanol. At last, the authors stained cells for 5 min with 0.1% crystal violet. The colonies with more than 50 cells were manually counted.
Transwell migration assay
The migratory capacity of pcDNA3.1/HAND2-AS1-transfected cells was assessed by employing a Transwell migration assay using a 24-well Transwell chamber (Corning, NY); 104 Hep3B and Huh-7 cells after being digested with trypsin were plated into the upper chamber in serum-free medium, with the lower chamber loaded with DMEM containing 10% FBS. After 24 h, transfected cells staying in the upper chamber were wiped out. Conversely, 20% methanol and 0.1% crystal violet were utilized for fixing and staining cells migrating through the membranes. Determination of migratory cell number in five random fields was accomplished with a microscope (Nikon Corporation, Tokyo, Japan).
RNA immunoprecipitation assay
An EZMagna RNA immunoprecipitation (RIP) kit (Millipore, Billerica, MA) and specific antibody (anti-Ago2) (Abcam) were required for RIP assay implementation, and the RIP assay was conducted exactly as recommended by the manufacturer's manuals. In short, Hep3B and Huh-7 cell lysates underwent incubation at 4°C with magnetic beads coupled with anti-Ago2 or IgG antibody overnight. Ago2-RNA complexes after washing were eluted, and coprecipitated RNAs were analyzed by qRT-PCR quantification.
Luciferase reporter assay
pMIR-REPORT luciferase vectors containing wild- and mutated-type HAND2-AS1 full-length (HAND2-AS1-WT and HAND2-AS1-MUT) or 3′-UTR of SOCS5 (SOCS5-WT and SOCS5-MUT) were manufactured and provided by Shanghai GenePharma Co., Ltd. (Shanghai, China). Additionally, miR-3118 mimics and NC mimics were commercially acquired from Shanghai GenePharma Co., Ltd. Cotransfection was performed in Hep3B and Huh-7 cells with the combination of HAND2-AS1-WT, HAND2-AS1-MUT, SOCS5-WT, or SOCS5-MUT and either miR-3118 mimics or NC mimics. The luciferase activities were separated and measured using the dual-luciferase reporter system (Promega).
Statistical analyses
GraphPad Prism software (La Jolla, CA) and SPSS, version 22.0 (IBM, Armonk, NY), were employed for all data analyses, with data reported as mean ± standard deviation (SD). Student's t-test and one-way analysis of variance (ANOVA) were employed for accomplishing the comparisons for different groups. Differences between groups were regarded as statistically significant when a p-value is less than 0.05 (*p < 0.05 and **p < 0.01).
Results
HAND2-AS1 augmentation inhibits liver cancer cell proliferation and migration
The TCGA database implicated the low level of HAND2-AS1 in liver cancer (Fig. 1A). To explore the potential involvement of HAND2-AS1 in cancer cells, the authors analyzed the expression changes of HAND2-AS1 in the normal liver epithelial cell line L02 and four liver cancer cell lines (Hep3B, HepG2, Huh-7, and BEL-7402). The results of qRT-PCR confirmed the differential expression between the two groups, and remarkably repressed HAND2-AS1 expression was observed in four liver cancer cell lines compared with normal cells (Fig. 1B). The authors chose two cell lines (Hep3B and Huh-7) due to the lowest HAND2-AS1 expression to construct stable HAND2-AS1 overexpression cell lines (pcDNA3.1/HAND2-AS1) and negative controls (pcDNA3.1). pcDNA3.1/HAND2-AS1 plasmids transfected into Hep3B and Huh-7 cells effectively reduced the HAND2-AS1 level in both cell lines (Fig. 1C). Overexpression of HAND2-AS1 inhibited cell viability and colony formation ability (Fig. 1D, E). The proapoptotic proteins (cleaved caspase3 and BAX) and antiapoptotic protein (BCL-2) underwent Western blot analysis which indicated that cleaved caspase3 and BAX level were increased upon HAND2-AS1 elevation, while BCL-2 was reduced after HAND2-AS1 elevation (Fig. 1F). Moreover, migration of cells treated with HAND2-AS1-overexpressing plasmids was also suppressed (Fig. 1G). lncLocator prediction demonstrated that HAND2-AS1 was abundant in the cytoplasm (Fig. 1H). Consistent with it, subcellular localization of HAND2-AS1 indicated its enrichment in Hep3B and Huh-7 cytoplasm (Fig. 1I).

HAND2-AS1 augmentation inhibits liver cancer cell proliferation and migration.

To explore the functions of HAND2-AS1 in a normal cell, the authors silenced it in the normal L02 cell line and conducted functional assays. Transfection efficiency for sh/NC, sh/HAND2-AS1#1, and sh/HAND2-AS1#2 was determined by qRT-PCR (Supplementary Fig. S1A). Then, the authors found that silence of HAND2-AS1 had no significant effect on L02 cell proliferation and apoptosis (Supplementary Fig. S1B, C). Altogether, HAND2-AS1 augmentation impaired the proliferative and migratory capacity of liver cancer cells.
HAND2-AS1 promotes SOCS5, a negative modulator of JAK/STAT3 signaling, through miR-3118 interaction
To address the targets of HAND2-AS1, the authors observed the HAND2-AS1-induced differential expression of multiple genes in Huh-7 cells through microarray analysis. It was noted that SOCS5 was one of the remarkably overexpressed genes (Fig. 2A). In addition, based on previous research, the role of SOCS5 has been documented as an inhibitor in HCC
31
and STAT3 signaling pathway inactivation.
32,33
The authors reasonably inferred that HAND2-AS1 mediated the tumor suppressor SOCS5 to exert its tumor-inhibiting role. Given that HAND2-AS1 was located in the cytoplasm, a ceRNA mechanism was further analyzed. The SOCS5 mRNA and protein levels were also determined to be downregulated in liver cancer cells (Fig. 2B, C). To determine whether HAND2-AS1 intervenes in SOCS5 expression in liver cancer cells, Western blot assays were conducted. An increased level of SOCS5 protein was observed in the presence of HAND2-AS1 upregulation (Fig. 2D). StarBase (
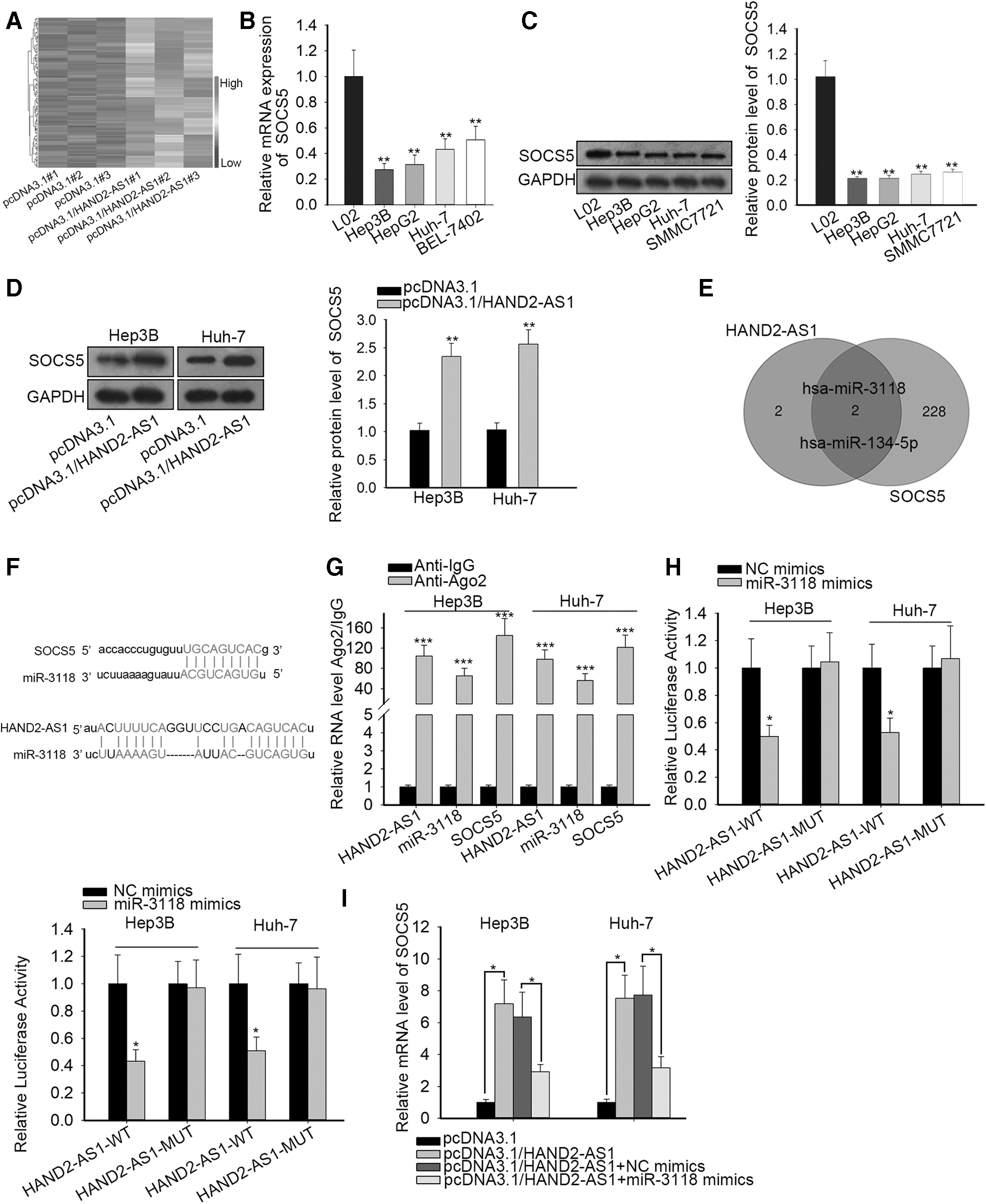
HAND2-AS1 promotes SOCS5, a negative modulator of JAK/STAT3 signaling, through miR-3118 interaction.
HAND2-AS1 facilitates SOCS5 expression to inactivate the JAK-STAT pathway by sponging miR-3118
To elucidate the implication of HAND2-AS1, miR-3118, and SOCS5 in adjusting the JAK-STAT pathway, the authors established stable knockdown SOCS5 cells and confirmed SOCS5 expression by qRT-PCR (Fig. 3A). The impact of SOCS5 on the JAK-STAT pathway was interrogated by Western blot assay. Significantly decreased SOCS5 protein was found after SOCS5 blockade, while phosphorylated STAT3 and downstream gene (c-Myc and PIM1) protein contents of JAK-STAT were enforced in the absence of SOCS5 with total STAT3 protein remaining unchanged (Fig. 3B). Furthermore, HAND2-AS1 overexpression alone suppressed STAT3 phosphorylation and downstream proteins of JAK-STAT. Nevertheless, the effects of HAND2-AS1 were subsequently countervailed when cotransfected with miR-3118 mimics into Hep3B and Huh-7 cells (Fig. 3C). Collectively, HAND2-AS1 augmented inactivation of the JAK-STAT pathway by upregulating SOCS5 through inhibiting miR-3118.
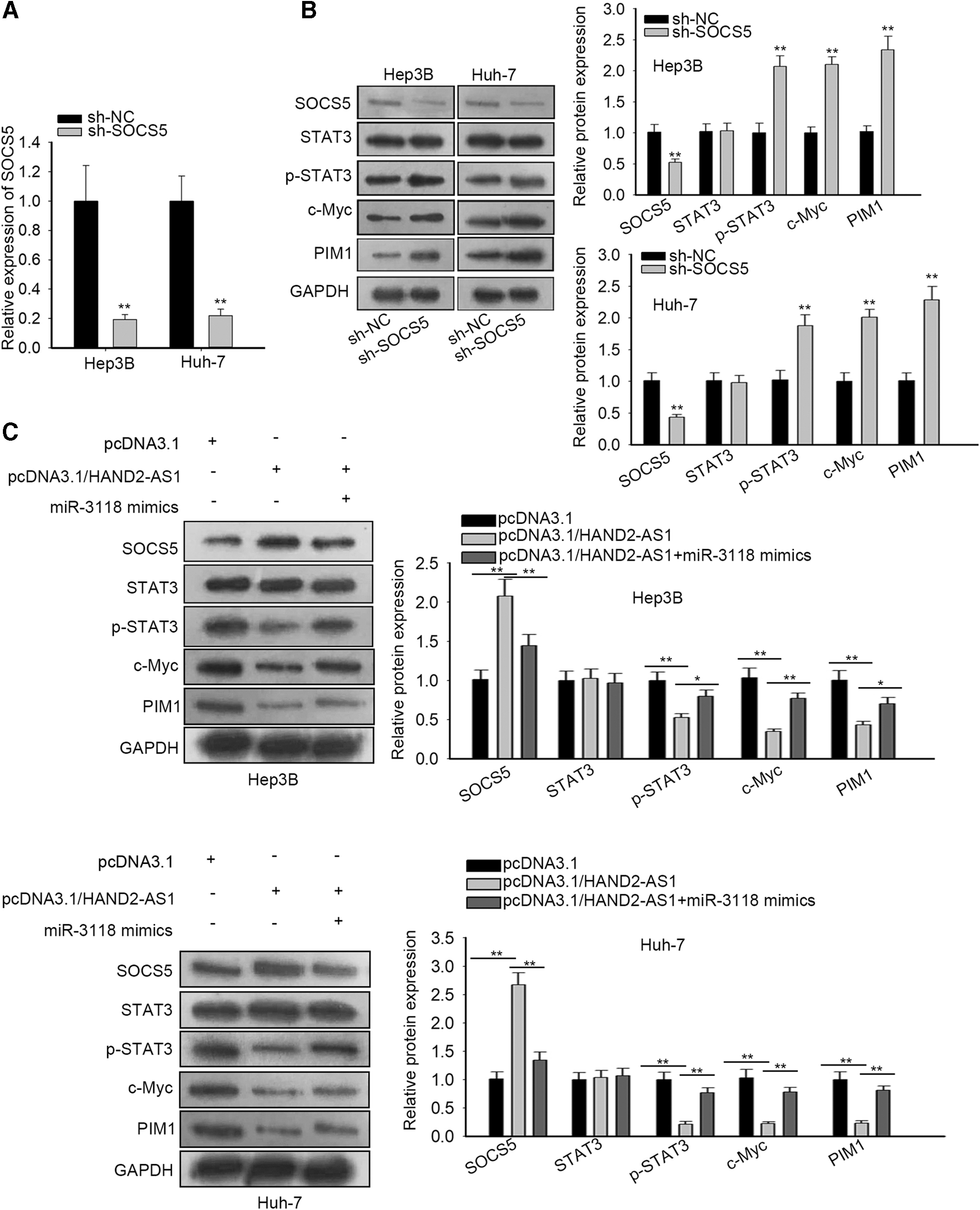
HAND2-AS1 facilitates SOCS5 expression to inactivate the JAK-STAT pathway by sponging miR-3118.
HAND2-AS1 mediates SOCS5 expression to hinder liver cancer cell growth and mobility through the JAK-STAT pathway
To confirm the HAND2-AS1 function and its role in liver cancer cell proliferation and migration through inactivation of the JAK-STAT pathway by upregulating SOCS5, rescue experiments were implemented. The viability of Huh-7 cells was tested, which indicated that SOCS5 inhibition reverted the effect of HAND2-AS1 on Huh-7 cell viability (Fig. 4A). The colony formation assay suggested that enhanced HAND2-AS1 attenuated the colony formation ability of Huh-7 cells and SOCS5 inhibition countervailed the impacts (Fig. 4B). Western blot assay indicated that the promoted Huh-7 cell apoptosis was reduced in the presence of HAND2-AS1 elevation (Fig. 4C). SOCS5 downregulation restored the decrease of Huh-7 cell migration caused by overexpressed HAND2-AS1 (Fig. 4D). All data suggested that HAND2-AS1 hindered liver cancer progression through SOCS5-mediated JAK-STAT pathway inactivation.

HAND2-AS1 mediates SOCS5 expression to hinder liver cancer cell growth and mobility through the JAK-STAT pathway.
Discussion
In spite of the current progress and advances made in the field of liver cancer research, liver cancer still acts as an alarming burden throughout the world and is also prevalent in China due to its high metastasis and high incidence rate. 2 –5 Hence, an in-depth understanding of liver cancer biology and its pathogenesis could be effectively used for diagnostic and prognostic biomarker identification.
lncRNAs induced the abnormal growth and metastasis of a considerable amount of malignant tumors, including liver cancer. 7,9,10 In the fields of liver cancer research, the underlying mechanism of lncRNAs was also unfolded; however, the suppressive effect of lncRNAs has rarely been fully discussed considering the surging reports on their reverse function on tumor progression. According to informed previous work, the oncogenic lncRNAs in liver cancer include MIR210HG, 11 DLX6-AS1, 12 SNHG16, 34 and so on. Antitumor lncRNAs such as LINC01554, LINC01093, and GAS5 have been reported before, which deserve more exploration. lncRNA HAND2 antisense RNA 1 (HAND2-AS1), a bona fide tumor inhibitor in multiple cancers, suppressed the malignant tumor phenotypes in esophagus squamous cell carcinoma, 16 osteosarcoma, 17 colorectal cancer, 18 and endometrioid endometrial carcinoma. 19 However, the tumor-suppressive impact of HAND2-AS1 was first validated in liver cancer in the present study.
Intense investigation about the JAK-STAT pathway demonstrated that its hyperactivation was strongly correlated with diverse tumor promotion processes, including liver cancer. 20,21,23 Consistent with a previous study, this study also unveiled the involvement of the JAK-STAT pathway in liver cancer. This work uncovered that HAND2-AS1 modulated the inactivation of the JAK-STAT pathway to pose an inhibitory effect on liver cancer proliferation and migration. SOCS5 was a recognized tumor suppressor, including in liver cancer, suppressor of the cytokine signaling family, and an inhibitor for JAK/STAT3 pathways. 24,26 The present study further proved the inactivation of JAK/STAT3 pathways caused by SOCS5 in liver cancer. The authors first revealed that HAND2-AS1 positively regulated SOSC5 to hinder the signal transduction of JAK/STAT3 pathways. Mechanically, HAND2-AS1 liberated mRNA of SOCS5 from inhibition of miR-3118. This study also unveiled the interplay between HAND2-AS1 or SOCS5 and miR-3118 in liver cancer for the first time.
In summary, the present work clarified that HAND2-AS1 accelerated the expression of SOCS5, the negative regulator of JAK2/STAT3 pathways, through competing against miR-3118 in liver cancer, which may be developed as a novel therapeutic approach for liver cancer treatment.
Footnotes
Acknowledgment
The authors would like to express great appreciation to all those involved in this research.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.