
Abstract
Background:
Cervical cancer (CC) is regarded as one of the most common gynecological malignancies. LncRNA DLX6-AS1 has been proven vital in various cancers, whereas its exact function is still largely unestablished in CC.
Materials and Methods:
The expression pattern of DLX6-AS1 and miR-16-5p in CC cells was determined by real-time quantitative polymerase chain reaction (RT-qPCR). ARPP19 expression was assessed by RT-qPCR and Western blot assays in CC cells. The precise function of DLX6-AS1 in CC was detected by Cell-Counting Kit-8 (CCK-8), 5-ethynyl-2′-deoxyuridine (EdU), caspase-3 activity, transwell, and Western blot experiments. RNA immunoprecipitation (RIP) and luciferase reporter assays were employed to certify the combination between miR-16-5p and DLX6-AS1 (or ARPP19). Nuclear cytoplasmic segmentation determined the localization of DLX6-AS1 in CC cells. A xenograft mouse model assay studied the influences of DLX6-AS1 silencing on CC progression in vivo.
Results:
Elevated DLX6-AS1 expression was disclosed in CC cells. DLX6-AS1 silence attenuated proliferation, migration, and epithelial–mesenchymal transition program as well as enhanced CC cell apoptosis. DLX6-AS1 was uncovered to sponge and negatively modulate miR-16-5p in CC. Besides, ARPP19 was uncovered as a downstream target gene of miR-16-5p in CC. Rescue experiments indicated that DLX6-AS1 enhanced the cellular process of CC cells through upregulating ARPP19. Moreover, in vivo assay confirmed that DLX6-AS1 promoted CC growth.
Conclusions:
DLX6-AS1 accelerates the progression of CC through sponging miR-16-5p and upregulates ARPP19, which offers a novel insight into prognosis and remedy of CC.
Introduction
Cervical cancer (CC) is known as one of the most lethal malignancies, and the primary reason of cancer-caused mortalities among women in especially less developed countries. Although the treatment of CC has been improved in several aspects including operative treatment and chemotherapy, metastasis and recurrence remain to be the main causes of sorrowfully poor 5-year survival rate of CC patients. 1 In consequence evacuation novel targets to overcome the dreadful CC is extremely urgent.
Possessing >200 nucleotides (nt) in length, long noncoding RNAs (lncRNAs) belong to a kind of important noncoding RNAs with no coding potential. 2,3 Recently, abundant evidence has been shown that lncRNAs have specific biological roles in a variety of cancers. 4,5 For instance, PCAT19 accelerates laryngocarcinoma cells proliferation through modulating the miR-182/PDK4 axis. 6 SBF2-AS1 enhanced CC progression through targeting miR-361-5p/FOXM1 axis. 7 Hepatocellular carcinoma up-regulated EZH2-associated long non-coding RNA aggravates tumorigenesis of colorectal cancer by countervailing miR-939-mediated transcriptional suppression of Bcl-Xl. 8 A recent study verified that DLX6-AS1 promotes hepatocellular carcinoma aggravation through regulating miR-203a/MMP-2 pathway. 9 DLX6-AS1 enhanced cell proliferation and invasion through inhibiting miR-181b in pancreatic cancer. 10 DLX6-AS1 drives the carcinogenesis of renal cell carcinoma through miR-26a/PTEN axis. 11 However, research on the biological impact of DLX6-AS1 in CC is still lacking.
Herein, the research is to uncover the role of DLX6-AS1 in CC progression. First, according to function experiments, DLX6-AS1 expression in normal healthy cervical cells and CC cells was investigated to acquaint with the biological role of DLX6-AS1. After that, the relationship between DLX6-AS1 and miR-16-5p was hypothesized and validated. Furthermore, the relationship between ARPP19 and miR-16-5p was verified. Besides, experiments in vivo were conducted, which further corroborated the function of DLX6-AS1 in CC. Eventually, it was confirmed that DLX6-AS1 accelerated the progression of CC through regulating miR-16-5p/ARPP19 axis, shedding new light on improving therapeutics in CC.
Materials and Methods
Cell culture
American Type Culture Collection (ATCC) provided the normal human cervical cell (End1/E6E7) as well as CC cells (SiHa, HeLa, C-33A, and CaSki). Then the cells were incubated in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) with the addition of 10% fetal bovine serum (FBS; Invitrogen), 100 U/mL penicillin (Invitrogen), and 100 μg/mL streptomycin (Invitrogen). All the cells were conserved at 37°C in a humid air with 5% CO2.
Cell transfection
Sh-DLX6-AS1#1/2/3 was transfected into CC cells to knock down DLX6-AS1, and sh/NC was utilized as negative control. MiR-16-5p mimics were used for miR-16-5p overexpression. PcDNA3.1/ARPP19 was employed for ARPP19 overexpression. Plasmid transfections were conducted by Lipofectamine 2000 (Invitrogen). Synthesized plasmids were all commercially gained from GenePharma (Shanghai, China). After transfection, cells incubated for 48 h were then further purified for the preparation of following experiments.
RNA extraction and quantitative real-time polymerase chain reaction
Trizol reagent (Takara, Otsu, Japan) was applied for total RNA extraction. Target mRNA was reverse-transcribed into complementary DNA (cDNA) with the employment of a reverse transcription kit (Takara), and Taqman Advanced miRNA cDNA Synthesis Kit (Waltham, MA) was applied to reverse transcribe micro-RNA (miRNA) into cDNA. Real-time quantitative polymerase chain reaction (RT-qPCR) was conducted by means of SYBR-Green Real-Time PCR kit. Then the 2−ΔΔCt method was utilized for calculating the relative RNA level of target genes. All the data were, respectively, normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6.
Cell proliferation assay
Cell proliferation was interrogated by adopting Cell-Counting Kit-8 (CCK-8) analysis. The transfected cells (1 × 103 cells per well) were planted in 96-well plates. After culturing for 0, 24, 48, 72, and 96 h, 10 μL of CCK-8 reagent was supplemented into each well. Then cells were continuously cultured for another 4 h and the optical density value was measured at 450 nm applying the microplate reader to assess cell proliferation.
5-Ethynyl-2′-deoxyuridine immunofluorescence assay
Cells were cultivated for 2 h with 5-ethynyl-2′-deoxyuridine (EdU) solution and afterward medium was removed. Thereafter, cells were fixed by PBS supplemented with 4% paraformaldehyde. Cells were then transferred to 70% ethanol and stained by use of Cell-Light™EdU Apollo®488 In Vitro Imaging Kit (RioBio, China).
Caspase-3 activity
Following the manufacturer's guides, the activity of caspase-3 in transfected SiHa and HeLa cells was tested using the caspase activity assay kit (Beyotime, Haimen, China). Samples were monitored by a microplate reader (Tecan Group, Männedorf, Switzerland).
Transwell assay
Transwell assay was utilized to test cellular migration. After transfection, cells were cultivated on the upper chambers that filled with serum-free DMEM. DMEM filled with 10% FBS was added into the bottom chamber. CC cells after 48 h incubation were then fixed by methanol and followed by staining with crystal violet. Thereafter, the number of migrated cells was counted.
Western blot analysis
Employing RNA immunoprecipitation assay (RIPA) lysis buffer (Beyotime Biotechnology, China), total proteins with protease inhibitor were isolated (Roche, China). Then protein concentration was assessed with the BCA™ Protein Assay Kit (Pierce, Appleton). Ten percent sodium dodecyl sulfate–polyacrylamide gel electrophoresis was employed for separating proteins. After electrophoresis, proteins were transferred onto polyvinylidene difluoride membrane. The membrane was then blocked by skim milk and probed with primary antibodies overnight at 4°C and then cultivated with secondary antibodies for >1 h at indoor temperature. The signals were visualized through the chemiluminescent detection system, and GAPDH was chosen for internal control.
RNA immunoprecipitation assay
RIPA was adopted by using Magna RNA-binding protein immunoprecipitation kit (Millipore, Billerica, MA). In brief, cell lysate and magnetic beads conjugating with anti-Ago2 were mixed together with RIP buffer. Anti-IgG was applied for normalization. Immunoprecipitated RNAs were extracted by Proteinase K. Eventually, the expression of purified RNAs was subjected to RT-qPCR.
Luciferase reporter assay
The luciferase reporter vectors pmirGLO (Promega, Madison, WI) were utilized to transfect the DLX6-AS1-Wt or DLX6-AS1-Mut vectors with NC mimics or miR-16-5p mimics, respectively, into cells. The pmirGLO-ARPP19-Wt or pmirGLO-ARPP19-Mut vectors were also cotransfected with miR-16-5p mimics or NC mimics into CC cells. Lipofectamine 2000 was utilized for transfection. After 48 h, Luciferase Reporter Assay System (Promega) examined the relative luciferase signals.
Nuclear–cytoplasmic fractionation
CC cells were segregated with a PARIS Kit (Ambion, Austin, TX). The collected cells were resuspended in cell fractionation buffer, and then cultivated on ice and centrifuged. The supernatant and nuclear pellet were harvested as the cytoplasm fraction and nucleus fraction for RNA extraction with TRIzol and TRIzol LS reagent (Invitrogen).
Xenograft mouse model
The animal study approved by the ethics committee of The 940th Hospital of Joint Logistics Support Force of Chinese People's Liberation Army was carried out. In brief, sh-NC and sh-DLX6-AS1#1 were, respectively, transfected into SiHa cells, followed by subcutaneous injection into 6-week-old BALB/c nude mice.
After injection for 7 d, tumor size was detected by caliper measurements every 4 d. After 28 d, the mice were sacrificed. Harvested tumors were photographed and weighed.
Immunohistochemistry
For immunohistochemistry (IHC) staining, after deparaffinization and rehydration by use of a graded ethanol and xylene, antigen retrieval was carried out by blocking endogenous peroxidase activity using 0.3% hydrogen peroxide. Then samples were treated with anti-Ki67 and anti-ARPP19 (Abcam, Cambridge, MA) at 4°C for one night, followed by secondary antibody incubation (Vector Laboratories, CA). Thereafter, protein was stained by diaminobenzidine (Beyotime, Shanghai, China) and counterstained using hematoxylin (Beyotime).
Statistical analysis
Assays were independently carried out three times, after which all the data were shown as the mean ± standard deviation. Statistical analysis was conducted by applying SPSS 20.0 software (Chicago, IL). Group differences were subjected to the analysis of Student's t-test or the one-way analysis of variance. A value of p < 0.05 was set as the threshold of statistical significance.
Results
DLX6-AS1 expression is extremely upregulated in CC cells and exerts oncogenic functions on the cellular process of CC cells
To probe the role of DLX6-AS1 in CC, the expression of DLX6-AS1 in normal End1/E6E7 and CC cells (SiHa, HeLa, C-33A and CaSki) was first investigated by RT-qPCR. The result revealed that DLX6-AS1 expression in CC cells was distinctly upregulated compared with normal human cervical cells (Fig. 1A). SiHa and HeLa cells expressing higher expression of DLX6-AS1 were selected for subsequent experiments. Afterward, the favorable knockdown efficiency of DLX6-AS1 due to transfection of sh-DLX6-AS1#1/2/3 in CC cells was presented in SiHa and HeLa cells (Fig. 1B). As shown in CCK-8 assay, absence of DLX6-AS1 efficiently inhibited cellular proliferation in CC (Fig. 1C). EdU assay suggested that DLX6-AS1 silence obviously reduced EdU positive cells in CC (Fig. 1D). To deeply analyze the influence of DLX6-AS1 knockdown on apoptosis in SiHa and HeLa cells, caspace-3 viability was investigated, revealing that suppression of DLX6-AS1 increased cell apoptosis (Fig. 1E). Furthermore, transwell assay validated that DLX6-AS1 depletion lowered the number of migrated cells in CC (Fig. 1F). Western blot assay elucidated as a result of repressed expression of DLX6-AS1, epithelial marker (E-cadherin) expression was significantly elevated, whereas mesenchymal markers (N-cadherin and Vimentin) were notably declined in CC cells (Fig. 1G). The result revealed that DLX6-AS1 knockdown was involved in retarding epithelial–mesenchymal transition (EMT) progress in CC. All the findings confirmed that downregulation of DLX6-AS1 inhibits proliferation, migration, and EMT progress as well as promotes apoptosis in CC cells.
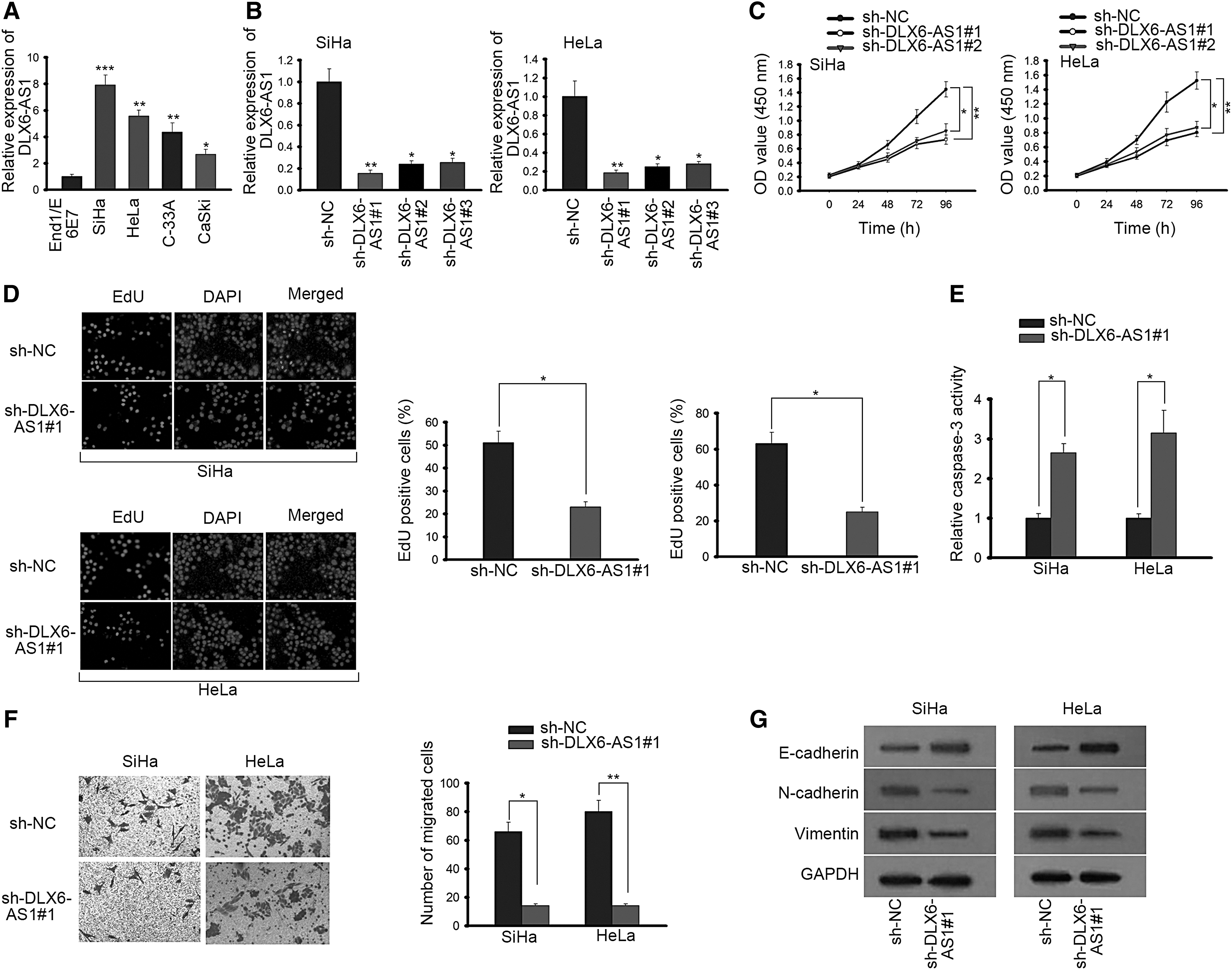
DLX6-AS1 expression is extremely upregulated and exerts oncogenic functions on the cellular process of CC cells.
DLX6-AS1 can bind with miR-16-5p in CC
Growing evidence has reported that lncRNAs could work as competing endogenous RNAs (ceRNAs) to sponge miRNAs and regulate progression of various cancers.
12
DLX6-AS1 was first examined to be located in cytoplasm (Fig. 2A). The cytoplasmic location of DLX6-AS1 prompted us to evaluate the potential ceRNA role of DLX6-AS1 in CC. Therefore, the molecular mechanism of DLX6-AS1 in SiHa and HeLa cells was further investigated. Through starBase v2.0 software (

DLX6-AS1 can bind with miR-16-5p in CC.
DLX6-AS1 regulates ARPP19 expression through competitively binding with miR-16-5p
It is reported that lncRNAs can competitively sponge miRNAs to regulate mRNAs expression. 16 Through searching starBase v2.0 software, ARPP19, the underlying target gene of miR-16-5p, was identified to have binding sites for miR-16-5p (Fig. 3A). Then RT-qPCR and Western blot assays exhibited that ARPP19 expression was prominently elevated in CC cells (Fig. 3B). As presented in Figure 3C and D, miR-16-5p overexpression (or DLX6-AS1 suppression) reduced the mRNA and protein level of ARPP19. Afterward, RIP was carried out to test the relationship among DLX6-AS1, miR-16-5p, and ARPP19. On the basis of the results from RIP, DLX6-AS1, miR-16-5p, and ARPP19 were identified to be all enriched in anti-Ago2-treated groups, but in the anti-IgG group, no obvious expression of the three was observed (Fig. 3E). Moreover, as shown in luciferase assay, luciferase activity of ARPP19-Wt reporter was remarkably alleviated in cells transfected with miR-16-5p mimics, whereas in pcDNA3.1/DLX6-AS1 cotransfected cells, the luciferase activity was enhanced, suggesting that DLX6-AS1 could regulate ARPP19 expression by targeting miR-16-5p (Fig. 3F). Meanwhile, no apparent change was presented in luciferase activity among ARPP19-Mut groups. All these data suggested that DLX6-AS1 could competitively bind with miR-16-5p and regulate ARPP19 expression.
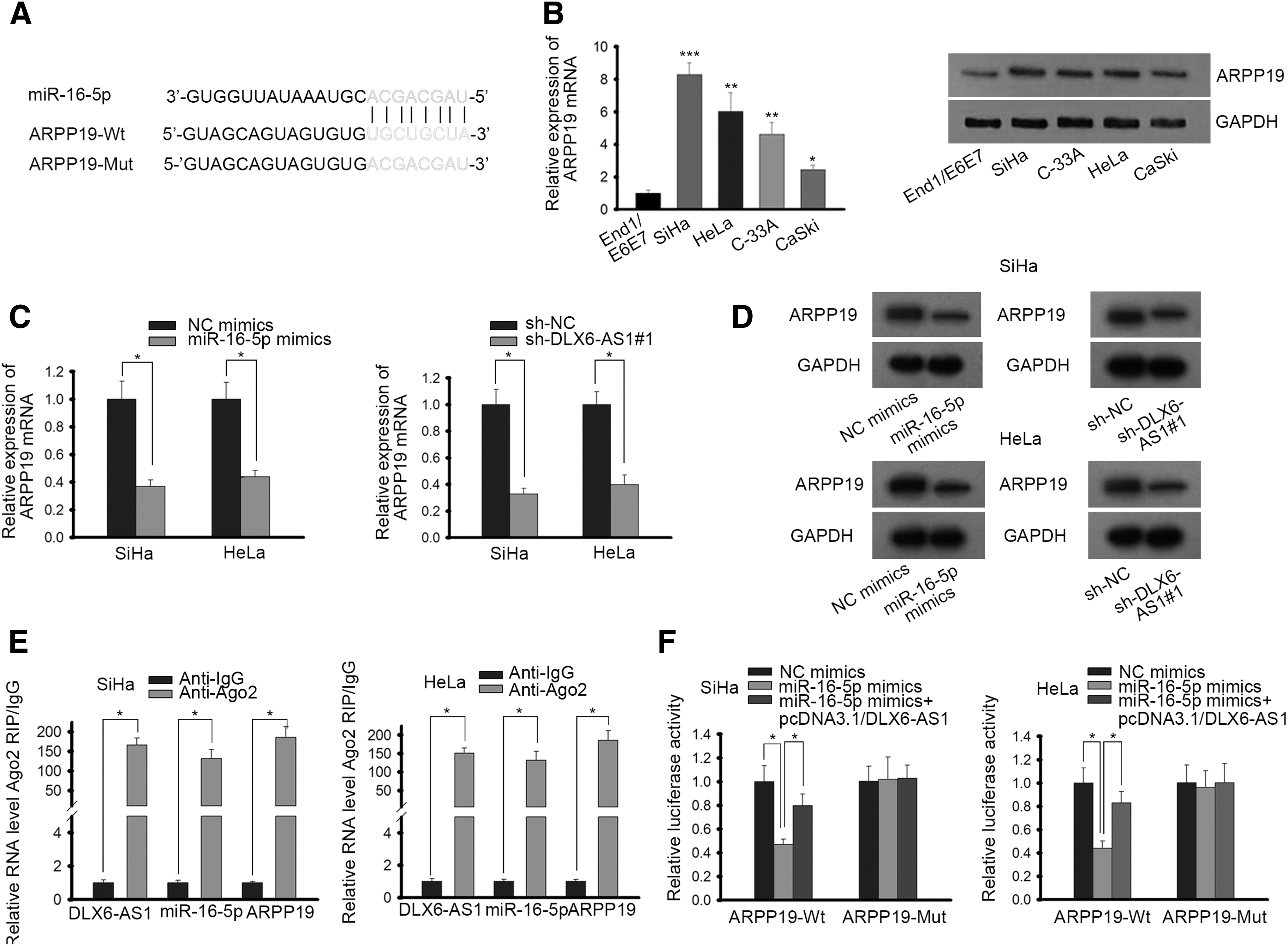
DLX6-AS1 regulates ARPP19 expression through competitively binding with miR-16-5p.
DLX6-AS1 aggravates the progression of CC through miR-16-5p/ARPP19 axis
A series of rescue assays were conducted to prove whether DLX6-AS1 could accelerate CC progression through the modulation of ARPP19 expression. ARPP19 expression was upregulated in pcDNA3.1/ARPP19 transfected cells (Fig. 4A). CCK-8 assay showed that by cotransfection of pcDNA3.1/ARPP19, the inhibitory influences of sh-DLX6-AS1#1 on cell proliferation in CC were abrogated (Fig. 4B). Similarly, EdU assay manifested that knockdown of DLX6-AS1 reduced EdU positive cells in CC, whereas overexpression of ARPP19 partially rescued it (Fig. 4C). As presented in Figure 4D, overexpression of ARPP19 countervailed the accelerating function of DLX6-AS1 silence on apoptosis in SiHa cells. Moreover, overexpression of ARPP19 promoted the cell migration in SiHa cells, which was inhibited by absence of DLX6-AS1 (Fig. 4E). Likewise, Western blot assay also presented that the enhanced expression of E-cadherin and the declined expression of N-cadherin and Vimentin triggered by DLX6-AS1 inhibition were partially reversed by enforced expression of ARPP19 (Fig. 4F). The results disclosed that DLX6-AS1 accelerated EMT progress of CC in an ARPP19-dependent way. To sum up, DLX6-AS1 drove proliferation, migration, and EMT progress as well as restrained cell apoptosis in CC by targeting miR-16-5p/ARPP19 axis.
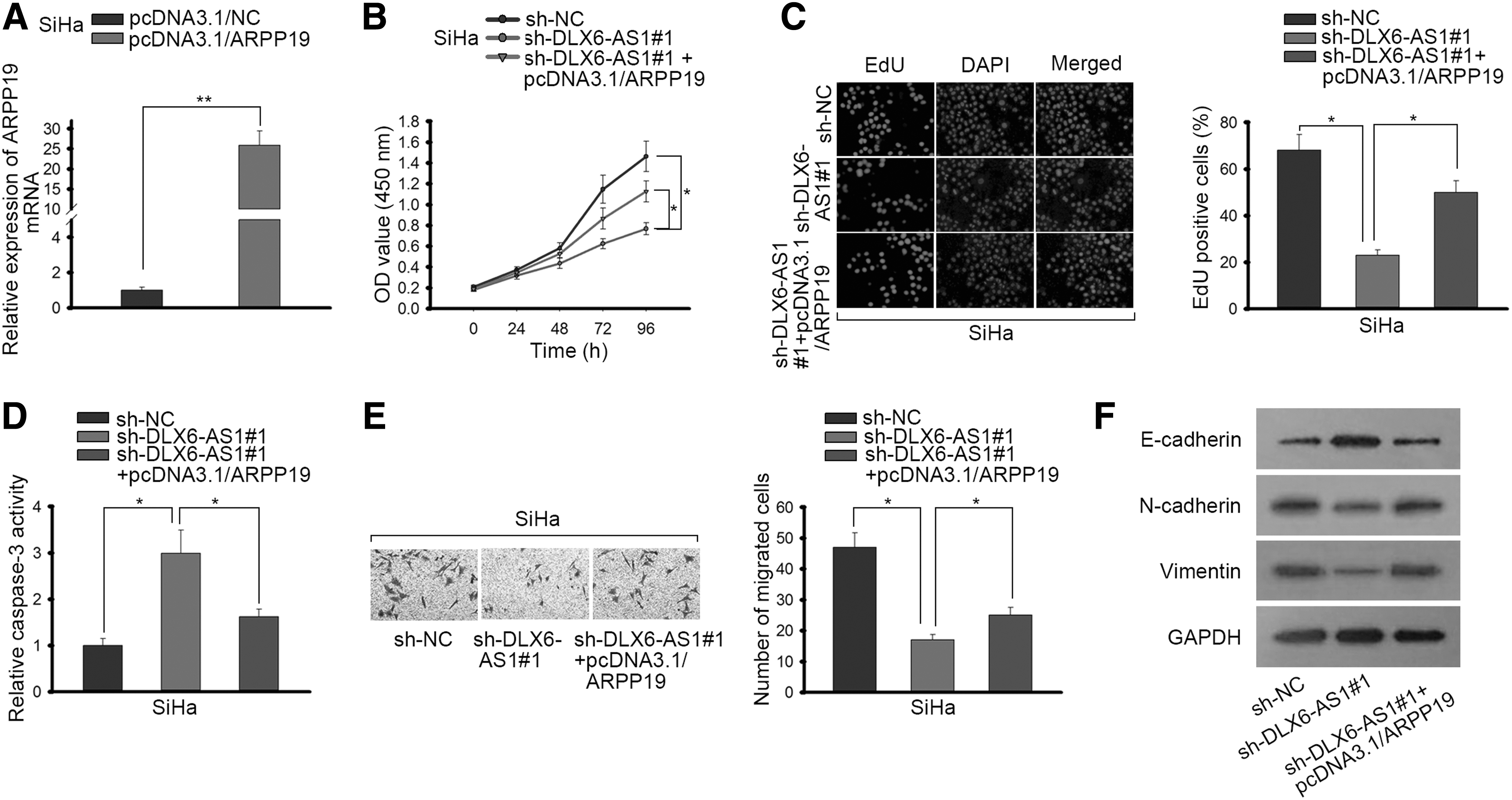
DLX6-AS1 promotes the progression of CC by miR-16-5p/ARPP19 axis.
DLX6-AS1 silencing inhibited CC growth in vivo
Finally, the function role of DLX6-AS1 in vivo was explored by conducting xenograft experiments. DLX6-AS1 knockdown apparently decreased the tumor sizes, volumes, and weights (Supplementary Fig. S1A–C). Subsequently, RT-qPCR analysis uncovered that sh-DLX6-AS1#1 impaired DLX6-AS1 expression and enhanced miR-16-5p expression (Supplementary Fig. S1D). ARPP19 mRNA expression (Supplementary Fig. S1D) and protein expression (Supplementary Fig. S1E) were both decreased based on the results of RT-qPCR and IHC assays. Moreover, IHC also manifested that the positivity of Ki67 in tumor tissue of sh-DLX6-AS1#1 group was noticeably less than that of sh-NC group (Supplementary Fig. S1E), implying that DLX6-AS1 inhibition could diminish CC cell growth in vivo. Hence, the mentioned results attested that DLX6-AS1 silencing inhibited CC growth in vivo.
Discussion
Numerous researches have suggested that lncRNAs exert extremely significant roles in different kinds of cellular biological processes, including CC. 17,18 LncRNA TPT1-AS1 accelerates cell growth and metastasis in CC through sponging miR-324-5p. 19 LncRNA non-coding RNA activated by DNA damage promotes cell proliferation and invasion through upregulating SIP1 expression in CC. 20 Nevertheless, the underlying mechanism of DLX6-AS1 in CC remains obscure. In this study, it was confirmed that DLX6-AS1 expression was much higher in CC cells than in normal cervical cells. Knockdown of DLX6-AS1 alleviated the proliferation, migration, and EMT progress as well as increased the apoptosis in SiHa and HeLa cells. Collectively, DLX6-AS1 drove the progression of CC.
Mounting evidence has suggested that miRNAs belong to small noncoding RNAs that could regulate the expression of downstream targets. 21 Several studies have verified that miRNAs serve as significant parts in the development and progression of some cancers. 22 What is more, recently, many reports indicated that lncRNAs function as ceRNAs to sponge miRNAs and then modulate various diseases. 23,24 For example, lncRNA CCAT1 upregulation enhances hepatocellular carcinoma progression by sponging miR-30c-2-3p. 25 LncRNA TFAP2A-AS1 declines breast cancer cell proliferation and invasion through miR-933/SMAD2. 26 To sequester miRNAs, lncRNA should express steadily at the post-transcriptional level. 27 Evidence from these assays proved that DLX6-AS1 was mainly occupied in the cytoplasm, suggesting the potential ceRNA role of DLX6-AS1 in CC cells. Besides, the existing study has illustrated that DLX6-AS1 could enhance the progression of several cancers such as glioma and nonsmall cell lung cancer through ceRNA regulatory network. 10,28,29 Afterward, it was figured out that miR-16-5p had binding sites for DLX6-AS1 through starBase v2.0 software. MiR-16-5p was found to play an antitumor role in multiple cancers. 30,31 In addition, it was proved that the DLX6-AS1 deficiency could upregulate miR-16-5p and the miR-16-5p overexpression could downregulate DLX6-AS1. Moreover, DLX6-AS1 coexisted with miR-16-5p in RNA-induced silencing complex (RISC). Furthermore, the combination between DLX6-AS1 and miR-16-5p was verified. All the data implied that DLX6-AS1 drove CC progression through binding with miR-16-5p.
It has been reported that ARPP19 expression was upregulated in various cancers. For example, miR-26a repressed thyroid cancer cell proliferation through targeting ARPP19. 32 In this study, miR-16-5p was predicted to bind with ARPP19. Then, ARPP19 level was measured to be promoted in CC cells and decreased by miR-16-5p overexpression or DLX6-AS1 inhibition. More importantly, ARPP19 was also detected to exist in RISC constructed by DLX6-AS1 and miR-16-5p. Besides, the affinity between ARPP19 and miR-16-5p was affirmed. In brief, DLX6-AS1 regulated ARPP19 expression by the way of competitively combining with miR-16-5p. Subsequently, rescue assays revealed that the influences of DLX6-AS1 suppression on CC progression could be partially recovered due to ARPP19 overexpression simultaneously. Finally, in in vivo assays, sh-DLX6-AS1 hindered tumor volume and weight as well as inhibited cell growth in CC. In vivo assay also revealed that DLX6-AS1 silencing downregulated the expression of DLX6-AS1 and ARPP19 together with upregulated miR-16-5p.
In conclusion, this study corroborated that DLX6-AS1 promoted the progression of CC by targeting miR-16-5p/ARPP19 axis, providing a new insight into treating CC.
Footnotes
Acknowledgment
The authors appreciate all people involved in this study.
Ethics Statement
The animal study was approved by the ethics committee of The 940th Hospital of Joint Logistics Support Force of Chinese People's Liberation Army.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.