
Abstract
Background:
NRAGE (neurotrophin receptor-interacting melanoma antigen-encoding gene homolog) has a complex role and regulates cell growth in different tumor cells. Although NRAGE was been discovered for more than 10 years ago, the function of NRAGE in hepatoblastoma (HB) cells is currently unknown.
Materials and Methods:
The expression of NRAGE was detected by reverse transcription-quantitative polymerase chain reaction assay or western blotting assay. Cellular apoptosis was analyzed to estimate the effect of NRAGE under radiation. The ability of clonogenic capacity was evaluated to confirm the influence of proliferation for NRAGE by radiation. The immunofluorescence assay was used to further study the expression of NRAGE under radiation. A nude mouse tumor xenograft model was constructed to confirm the effect of NRAGE deficiency under radiation conditions in vivo.
Results:
The authors determined that deletion of NRAGE significantly inhibited HB cell proliferation in vitro and in vivo, and NRAGE knockdown apparently sensitized HB cells to ionizing radiation (IR). Further mechanistic studies revealed that NRAGE plays a critical role in homologous recombination by inhibiting the expression of RNF8 (ring finger protein 8) and BARD1 (BRCA1 associated RING domain 1) and the recruitment of RAD51.
Conclusions:
The authors demonstrated that downregulation of NRAGE sensitizes HB cell lines to IR in vitro and in vivo. It provides a promising therapeutic strategy for HB patients by specifically targeting NRAGE.
Introduction
Hepatoblastoma (HB) represents 80% of hepatic malignant tumors in children and is usually diagnosed before 3 years of life. 1 The therapeutic strategies for HB patients mainly include complete surgical resection and neoadjuvant chemotherapy. 2,3 However, due to unresectable features of some HB patients and chemo-resistance, the eventual survival of HB remains poor. 4 Molecular targeting therapy is a powerful strategy for the treatment of cancer. Therefore, it is of virtual importance that the authors reveal the relevant mechanisms in HB.
DNA damaging therapeutic strategies, including ionizing radiation (IR), are frequently used in the treatment of solid tumors in the clinic. 5 However, the development of chemo-resistance or radio-resistance, which is partially induced by the abnormal DNA damage repair (DDR), limits the therapeutic effects of a DNA damaging therapeutic strategy. 6,7 Thus, targeting abnormal DDR pathways is thought to be a promising way to circumvent chemo-resistance or radio-resistance. 8,9 DNA lesions mainly include single-strand breaks and double-strand breaks (DSBs), among which DSBs are the most serious lesions. Homologous recombination (HR) and nonhomologous end-joining (NHEJ) are two critical DSB repair pathways in eukaryotes. 10 NHEJ restores the majority of IR-induced DSBs, but is an error-prone pathway and functions throughout the cell cycle. 11 HR preferentially repairs replication fork collapse-induced DSBs, and it is an error-free pathway that is restricted to the S/G2 phase of the cell cycle. 12 Upon DSB induction, a cluster of protein expression, relocalization, and modification is triggered, such as the early DNA damage sensor complex MRN (Mre11-Rad50-NBS1) which rapidly discriminates damage site through autophosphorylation upon genomic insult and accumulates ataxia telangiectasia mutated (ATM) to DSBs. 13 –16 This interaction causes the ATM dimer to separate into active monomers and phosphorylate the Histone's H2A variant (H2AX), then numerous downstream effectors, including ring finger protein 8 (RNF8), are recruited to DSBs and lead to the formation of K63-linked polyubiquitin chains on histones at DSBs. 17 This ubiquitination cascade is responsible for the localization of repair mediators, including BRCA1 associated RING domain 1 (BARD1) 18 and recruitment of recombination protein A (RAD51) (Fig. 1). 19
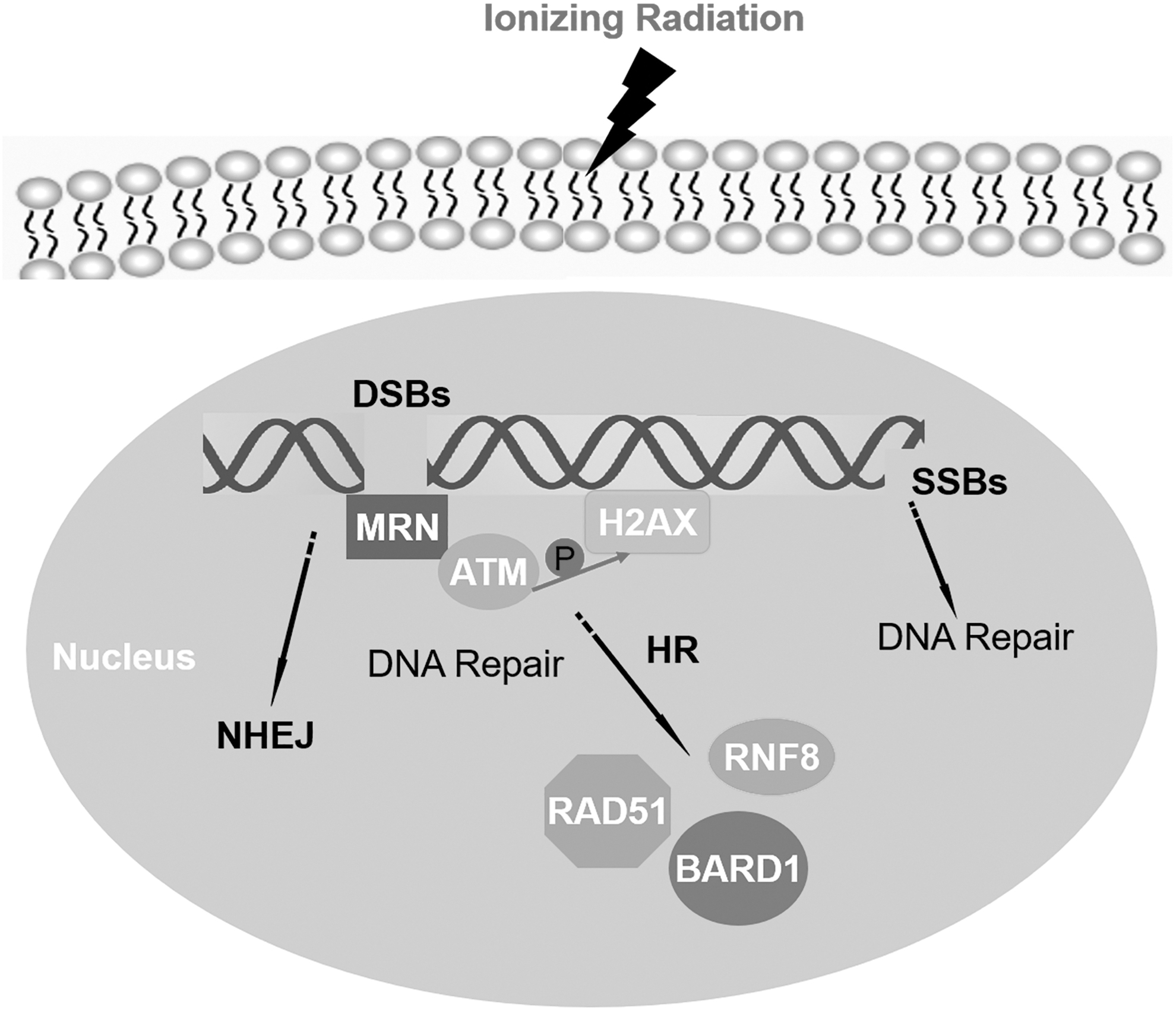
Schematic diagram of DNA damage repair process.
NRAGE (neurotrophin receptor-interacting melanoma antigen-encoding gene homolog), a critical adaptor protein, is characterized by a unique region of about 200 amino acids and plays an important role in numerous life activities. 20,21 NRAGE (MAGED1), as a member of the MAGE gene family, different from other recognized MAGE members, is a ubiquitous expressed gene. 22 NRAGE is reported to interact with transcription factors (Dlx/Msx, Che-1), nucleoprotein (PCNA, TBX2), and cell surface receptors (p75NTR, UNC5H1, ROR2, and TrkA) to participate in cell differentiation, apoptosis, cell cycle and tumorigenesis, and metastasis. 23 It has been reported that NRAGE positively regulates esophageal cell proliferation through binding with proliferating cell nuclear antigen (PCNA) using its DNAPIII domain, 24 and the authors also found that NRAGE has a crucial effect on HR and enhances esophageal cancer cell chemoresistance both in vivo and in vitro. Mechanistically, NRAGE regulates the stability of RNF8 and BARD1 by interacting with the RING domains of these two proteins to form a new ternary complex to participate in DDR. 18
Prior research increasingly determined that NRAGE has a complex role in different tumors and has a close relationship with the prognosis of tumor patients. 23 However, the function of NRAGE in HB cells has never been reported up to now. In present study, the authors preliminarily revealed that RNF8 and BARD1 mediated NRAGE-dependent HR in HB cells, and NRAGE is required for RAD51-mediated HR in HB cells. What's more, the authors also revealed that downregulation of NRAGE sensitizes HuH6 cells to IR in vitro and in vivo experiments. These results provide a potential molecular targeting therapeutic strategy for HB patients.
Materials and Methods
Cell culture
HB cell lines HepG2 and HuH6 cells obtained from American type culture collection (ATCC, MD) and the Cell Bank of Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China) were maintained in Dulbecco's modified Eagle's medium or minimum essential medium (MEM) at 37°C in a humidified atmosphere with 5% CO2. Media were supplemented with 1% MEM Nonessential Amino Acid Solution (Gibco, Grand Island, NY), 10% fetal bovine serum (Gemini Bio-Products, Calabasas, CA), 100 U/mL penicillin, and 100 U/mL streptomycin.
Plasmid construction and transfection
The human NRAGE interference plasmid is a 19-nt fragment inserted into the psuper-egfp vector under the control of the H1 promoter to disrupt the targeted AAGATGAAAGTGCTGAGATTC sequence. Lentivirus plasmids carrying the small hairpin NRAGE interference (NRAGE-shRNA) were constructed according to the previous study. 18 Lentivirus-mediated shRNA was packaged in 293T cells and infected HepG2 and HuH6 cells under Polybrene (Santa Cruz Biotechnology, Santa Cruz) for 24 h. In the end, positive transfected HepG2 and HuH6 cells were selected using 2 μg/mL puromycin (Invitrogen, Carlsbad, CA) for 7–10 d.
Apoptosis assay
Cellular apoptosis was analyzed by flow cytometry. FITC-conjugated Annexin V (eBioscience, CA) and propidium iodide (eBioscience) were used to stain HuH6 cells, which were irradiated with 0, 2, 4, and 8 Gy and incubated for 48 h at 37°C with 5% CO2. Each sample was quantified by flow cytometry (FCM), and the data were analyzed by FACS Vantage SE system (BD Biosciences, San Jose, CA).
Colony survival assay
Cell survival following different radiation exposure was used to detect the ability of clonogenic capacity and colony formation of HB cells. Briefly, HuH6 cells were diluted into single cell suspensions to seed into six-well plates according to the appropriate number of cells of different doses and irradiated with 0, 2, 4, and 8 Gy IR after being incubated for 6–12 h at 37°C with 5% CO2, respectively. After incubation intervals of 2 weeks, the cells in six-well plate were stained with 1% crystal violet and manually counted.
In vivo tumorigenicity
Ten male 4-week-old nude mice purchased from Shanghai Super-B&K Laboratory Animal Corp. Ltd. (Shanghai, China) were used for evaluating the inhibitory function of the deletion of NRAGE in vivo experiment. All animal experiments were approved by the animal center of Shanghai Tenth People's Hospital of Tongji University and were performed following the Guidelines for the Care and Use of Animals for Scientific Research.
HepG2 cells (1 × 107) stably transfected with Vector or NRAGE were subcutaneously injected into the left or the right gluteal region of the male nude mice (n = 10), respectively. Subsequently, the nude mice were randomly divided into two equivalent groups, which were radiation group and nonradiation group. A total radiation dose of 14 Gy (2 Gy/fraction every 3 d for 21 d) was delivered locally to nude mice using a linear accelerator. The tumor volume was measured every 3 d, and tumor size was calculated according to the following formula: Volume (mm3) = width 2 (mm2) × length (mm)/2. All nude mice were euthanized after 21 d, and tumors were weighted after ethical dissection and photography.
Immunofluorescence assay
After the treatment, HuH6 cells were incubated with 4% paraformaldehyde for 15 min and blocked with 5% bovine serum albumin (BSA) for 1 h, then stained with relevant primary antibodies overnight at 4°C. The next day, the cells were stained with the secondary antibodies (DyLight™ 488- or 555-conjugated AffiniPure Donkey Anti-Mouse IgG) at 37°C for 1 h. Finally, the cells were stained with 4,6-diamidino-2-phenylindole (Invitrogen). All images were observed and pictured using confocal fluorescence microscopy (Carl Zeiss, Jena, Germany). The main primary antibodies used in Confocal immunofluorescence (IF) are as follows: NRAGE (SC-136552; Santa Cruz Biotechnology), RAD51 (ab133534; Abcam, Cambridge, United Kingdom), and γH2AX (ab26350; Abcam).
Reverse transcription-quantitative polymerase chain reaction
The expression of NRAGE mRNA was detected by a reverse transcription-quantitative polymerase chain reaction (RT-qPCR) assay. The total RNA was isolated with TRIzol Reagent (Invitrogen) from the HB cell lines following the manufacturer's protocol. RT-qPCR was performed with PrimeScript RT Reagent Kit (DRR0037A; Takara, Dalian, China) and Fast SYBR Green Master Mix (Applied Biosystems, Foster City, CA). For standardization, GAPDH mRNA was quantified in each sample. Relative gene expression was analyzed using the comparative 2−ΔΔCt method. The forward primer sequence for human NRAGE was 5′-CATCGTTGCTGCCTTCTTCG-3′, and the reverse sequence was 5′-ATCTCATTGGCAGTTGGGGG-3′. The forward primer sequence for human GAPDH was 5′-CGGAGTCAACGGATTTGGTCGTAT-3′, and the reverse sequence was 5′-TGCTAAGCAGTTGGTGGTGCAGGA-3′.
Western blotting assay
Cells were lysed on ice with strong lysate RIPA (Beyotime, Jiangsu, China) containing protease inhibitor for 15 min and then were centrifuged at 12,000 rpm in a 4°C centrifuge. After quantification using Pierce BCA Protein Assay Kit (Thermo Scientific), 80 μg of total protein was loaded in each lane for separation and electrophoretically transferred to a nitrocellulose membrane (GE Healthcare), followed by the block with 5% BSA for an h and indication with primary antibodies overnight at 4°C. The membranes were washed with Phosphate Buffered Saline with 0.1% Tween-20 (PBST) thrice, and fluorescence-conjugated secondary antibodies with appropriate dilution were against primary antibodies for an h at 37°C. The membranes were visualized using the Odyssey Infrared Imaging System (LI-COR, Nebraska, NE). Then, final analysis of the protein expression was carried out using ImageJ. 25 The main antibodies used in the western blot assay are as follows: GAPDH (#5174; Cell Signaling Technology, Boston, MA), NRAGE (SC-136552; Santa Cruz Biotechnology), RNF8 (ab105362; Abcam, Cambridge, United Kingdom), and BARD1 (SC-11438; Santa Cruz Biotechnology).
Statistics
In this study, quantitative data from three independent assays are presented as mean ± standard deviation. Two-tailed Student's t-test or one/two-way analysis of variance (ANOVA) was performed as appropriate. The p-values less than 0.05 were defined as *, while less than 0.01 were deemed as ** and less than 0.0001 were deemed as ****, all were considered significantly.
Results
The expression of NRAGE mRNA and protein was effectively inhibited by Lentivirus-mediated shRNA in HB cell lines
HB cell lines (HepG2 and HuH6) were transfected with lentiviral expressing control shRNA and NRAGE shRNA. The stable cells transfected with NRAGE shRNA and control shRNA were selected under puromycin (1 μg/mL). Then, RT-qPCR assay was used to detect the expression of NRAGE mRNA in NRAGE-shRNA group and NC (negative control)-shRNA group. As shown in Figure 2A, NRAGE in NRAGE-shRNA group was significantly lower compared with the NC-shRNA group. What's more, western blotting assay showed that NRAGE protein level was also inhibited in NRAGE-shRNA group compared with NC-shRNA group (Fig. 2B). To further confirm their results, the IF assay was performed for authentication (Fig. 2C, D), and the fluorescence intensity of NRAGE was also correspondingly decreased consistent with western blot assay. All above results revealed that both mRNA and protein levels of NRAGE were effectively inhibited by NRAGE-shRNA in the HB cells.

NRAGE-shRNA significantly inhibits the expression of NRAGE in HB cells.
Knockdown of NRAGE sensitizes HuH6 cells to IR in vitro
IR is a commonly used DNA-damaging induced cancer therapeutic strategy. 26 To examine the response of NRAGE knockdown HB cells to IR, colony formation analysis was used to detect for HuH6 cell proliferation exposed to IR. From the results, the authors found that the colony number in NRAGE-shRNA group was significantly reduced compared with NC-shRNA group, indicating that NRAGE plays a potential role in HuH6 cell survival (Fig. 3A, B). In addition, the average lethal dose (D0) and survival curve shoulder zone (Dq) of the NRAGE-shRNA group were decreased, while the sensitive enhancing ratio (SER = 1.103) was increased under the NRAGE deficiency (Fig. 3C). This result indicated that HuH6 cells in NRAGE-shRNA group were more sensitive to IR therapy. From previous reports, the authors know that cells frequently end up with apoptosis if DNA damage could not be repaired in time. 27 Therefore, to examine whether deletion of NRAGE can induce apoptosis, HuH6 cells transfected with NRAGE-shRNA or NC-shRNA were exposed to the irradiation of 0, 2, 4, and 8 Gy, respectively, and the apoptosis rate was detected by flow cytometry after 48 h of radiation therapy. NRAGE deficiency markedly enhanced cell apoptosis induced by radiation; interestingly, IR also significantly induced HuH6 cell apoptosis in a dose-dependent manner (Fig. 3D, E), and the apoptosis of NRAGE-shRNA/HuH6 cells exposed to the irradiation of 8 Gy was especially aggravated in comparison with NC-shRNA/HuH6 cells (Fig. 3E). All above results determined that deletion of NRAGE highly improved the radiosensitivity.
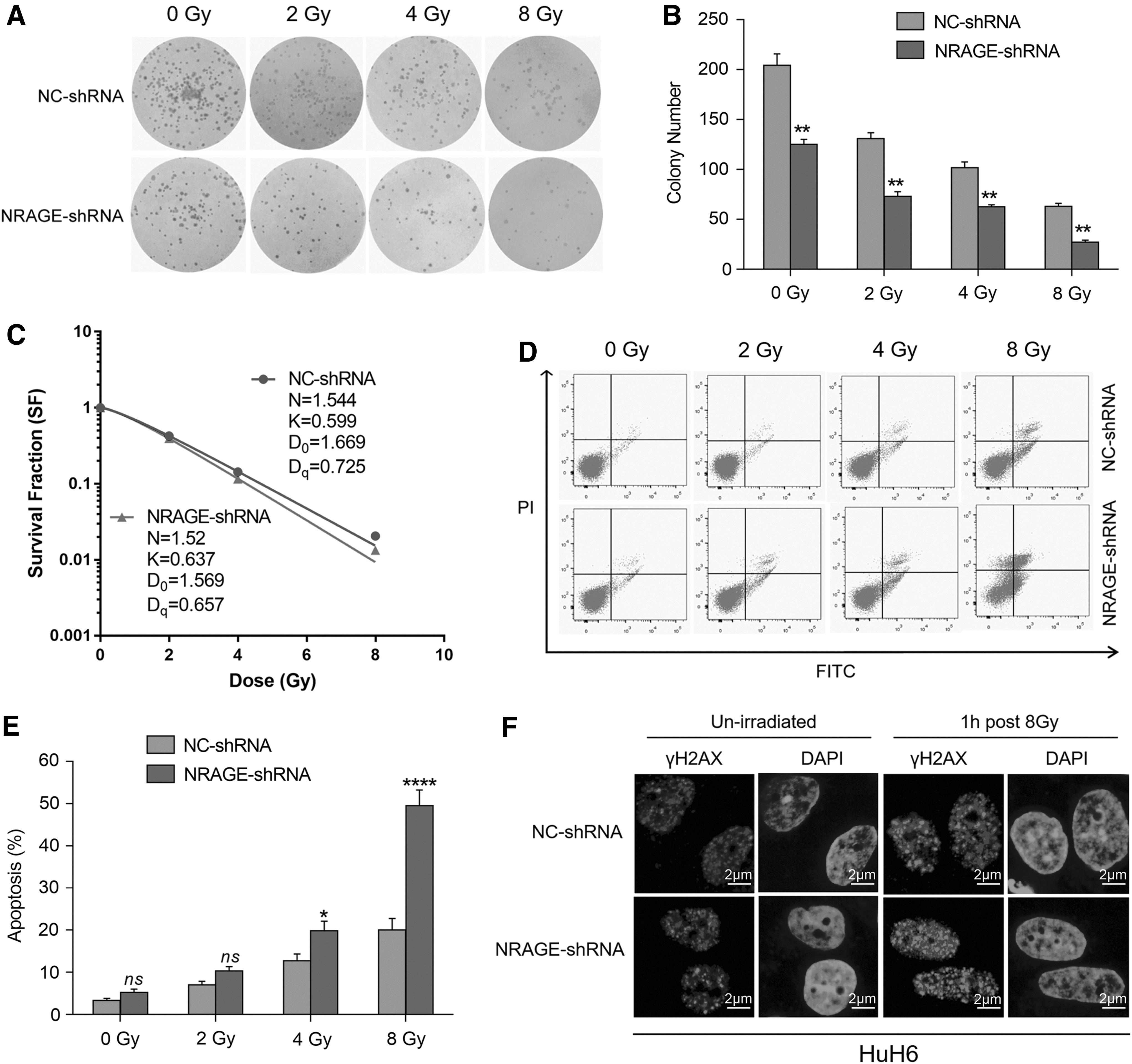
Knockdown of NRAGE enhances HuH6 cell sensitivity to radiation in vitro.
Knockdown of NRAGE enhances HB cells' sensitivity exposed to radiation and inhibits tumor proliferation in vivo
To further confirm the in vivo effect of NRAGE deficient under radiation conditions, the authors constructed a nude mouse tumor xenograft model injected by HepG2 cells, which was infected with lentivirus expressing shRNA against NRAGE (shRNA-NRAGE) or its NC-shRNA. The authors measured the volume of the nodules every 3 d to determine whether the deletion of NRAGE could enhance HepG2 cell radiosensitivity. Their results displayed that either interference with NRAGE or radiation alone significantly lessened the tumor volume compared to the untreated groups. In addition, the tumor volumes and weight in NRAGE-shRNA group, especially when exposed to irradiation, were reduced more visibly (Fig. 4). Therefore, these results demonstrated that silencing of NRAGE enhanced the sensitivity of HepG2 cells exposed to radiation and evidently inhibited tumor growth in vivo.

Knockdown of NRAGE enhances HB cells' radiosensitivity and inhibits tumor growth in vivo.
BARD1 and RNF8 mediate NRAGE-dependent HR inhibition
As reported in previous work, NRAGE plays an important role in the repair of DSBs by HR. 18 Therefore, the authors detected the foci formation of γH2AX in Huh6 cells irradiated by 8 Gy IR. The authors found that knockdown of NRAGE resulted in an obvious increase of γH2AX foci, and the formation of γH2AX IR-induced foci (IRIF) significantly increased after NRAGE knockdown under 8 Gy IR (Fig. 3F). The authors also found that knockdown of NRAGE in esophageal cell EC9706 results in degradation of RNF8 and BARD1 proteins instead of other DDR proteins, such as BRCA1, 53BP1, and PARP1. 18 To test whether NRAGE regulates RNF8 and BARD1 in HB cells, the expression level of NRAGE in HuH6 cells was interfered. From the results the authors found that the protein level of RNF8 and BARD1 was significantly reduced in NRAGE-shRNA group compared with NC-shRNA group (Fig. 5A, B). These data demonstrated that NRAGE participation in HR may be through regulating the protein level and recruitment of RNF8 and BARD1 in HB. As known to all, DDR plays an essential role in the cancer cell survival. To determine whether the inhibition of cell proliferation induced by the downregulation of NRAGE can be reversed by the overexpression of RNF8 and BARD1, GFP-RNF8 or GFP-BARD1 was transfected into HuH6/NC-shRNA and HuH6/NRAGE-shRNA cells (Fig. 5C). Intriguingly, overexpression of RNF8 and BARD1 partly rescued the inhibition of HuH6 cell growth (Fig. 5D, E). The above data indicated that NRAGE may have an effect on HR through altering the level of RNF8 and BARD1, and this potential mechanism explains why deletion of NRAGE enhances HB cells' radiosensitivity.

BARD1 and RNF8 mediated NRAGE-dependent HR inhibition.
NRAGE is required for RAD51-mediated HR in HB cells
Upon formations of DSBs, RNF8 and RNF168 are recruited to DSBs and lead to the K63-linked polyubiquitin chains located into histones at DSBs. 17 This ubiquitination cascade is responsible for the localization of the downstream repair factors, including BARD118 and RAD51. 19 Given that NRAGE can regulate the expression of RNF8 and BARD1 in HB cells, the authors speculate that NRAGE is required for RAD51-mediated HR.
To confirm the relationship of NRAGE and RAD51, the formation of RAD51 IRIF was detected after 1 h exposure to 8 Gy IR in HuH6/NC-shRNA and HuH6/NRAGE-shRNA cells. IF assay displayed that NRAGE deficiency resulted in significantly reduced IFIR formation of RAD51 following treatment with 8 Gy IR (Fig. 6A). To further identify the relationship of NRAGE with RAD51, the authors detected the formation of RAD51 IRIF at different time points after exposure to 8 Gy of IR (Fig. 6B). The RAD51 IRIF formation in NRAGE-shRNA group was markedly inhibited compared with NC-shRNA group at 1 h time point. Furthermore, the RAD51 IRIF intensity in NC-shRNA group was brighter than NRAGE-shRNA group, especially at the point of 1 h. Thus, it can be seen that RAD51-mediated HR was impaired after NRAGE knockdown following treatment with IR.
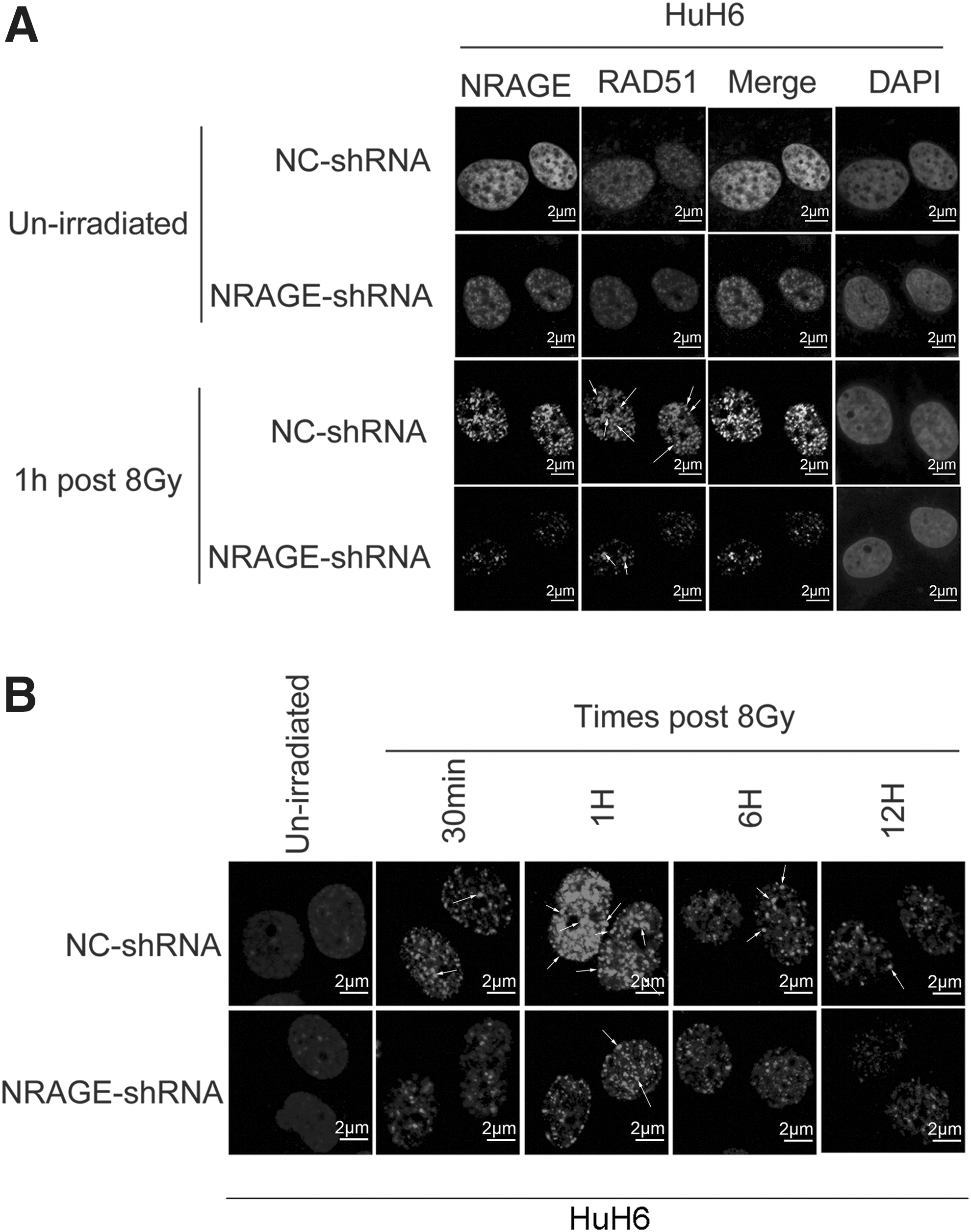
NRAGE is required for RAD51-mediated HR in HB cells.
Discussion
NRAGE is reported to participate in cellular homeostasis and oncogenesis, such as tumor occurrence, invasion, and metastasis. 28 –30 It was reported that NRAGE positively regulates esophageal cell proliferation through binding with PCNA using its DNAPIII domain, 24 and previous studies also found that overexpression of NRAGE interacting with apoptosis-antagonizing transcription factor affects the prognosis and progression of hepatocellular carcinoma. 31 As known to all, the function of NRAGE in HB cells has never been reported. In present research, the authors revealed that NRAGE takes an important role in HR in HB cells through regulating the expression and recruitment of RNF8, BARD1, and RAD51, and deletion of NRAGE significantly enhances the radiotherapy sensitivity of HB cells in vivo and in vitro.
Results from western blotting assay showed that NRAGE deficiency highly downregulated the expression of RNF8 and BARD1 in HB cells, which indicated that NRAGE participates in HR in HB cells. This result is consistent with a previous work that NRAGE regulates the RNF8 and BARD1 expression in esophageal cancer (EC) cells. 18 Then, the IRIF formation of RAD51 in HuH6 cells treated with IR showed that knockdown of NRAGE significantly impairs the recruitment of RAD51 in HB cells. All the above results indicated that NRAGE plays an important role in HB through regulating the expression and recruitment of RNF8, BARD1, and RAD51.
A previous work also revealed that NRAGE positively facilitates esophageal cell proliferation through binding with PCNA using its DNAPIII domain in the nucleus of the esophageal cells. 24 Consistently, in this article, the authors found that NRAGE is localized to the nucleus and served as an oncogene in HB cells evidenced by the results from colony formation assay and apoptosis assay, which showed that knockdown of NRAGE apparently inhibited HB cell proliferation and induced cell apoptosis. Shimizu et al. also demonstrated that NRAGE serves as a tumor promoting factor in hepatocellular carcinoma. 31 Moreover, results from Salehi et al. demonstrated that NRAGE interacts with p75 neurotrophin receptor (p75NTR) to promote the cell apoptosis. 32 However, Du et al. revealed that NRAGE inhibits proliferation, migration, and invasion of human breast cancer cells through upregulating the expression of p53, p21, and E-cadherin and downregulating the expression of β-catenin. 29 Three reasons can account for the different roles of NRAGE. First, NRAGE protein contains different functional domains (such as DNAPIII, IRD, and MHD), and the different domains act differently in the process of tumor development. 23,24 For example, Bragason and Palsdottir demonstrated that NRAGE can interact with CytPrP leading to Neuronal cell apoptosis through its repeat region (IRD domain). 33 Second, NRAGE has different subcellular localizations in different tumor cell lines. 34 Third, NRAGE regulates the expression of different proteins or comprises different complexes through binding with different proteins depending on cell type. For example, NRAGE is reported to interact with Dlx/Msx, TrkA to regulate cell differentiation and interact with XIAP-TAKITAB1 complex, p75NTR, Che-1, UNC5H1, CytPrP to regulate cell apoptosis. 23 Taken together, the authors can draw a conclusion that NRAGE has very complicated or contrary roles in tumorigenesis for its interaction with different proteins and different subcellular localizations in specific tumors.
Radiation therapy can play a role in the management of patients with HB, 35 but increased DNA repair efficiency contributes to chemo-resistance or radio-resistance results in poor outcomes in tumor patients. 6,7 Thus, targeting DDR pathways may be a promising way to enhance chemosensitivity or radiosensitivity. In the present study, knockdown of NRAGE enhanced the sensitivity of HB cells to IR by downregulating RNF8 and BARD1 expression and disturbed the recruitment of RNF8, BARD1, and RAD51. This corroborates a previous article which found that NRAGE is highly expressed in IR resistant cells and esophageal cancer tissues. 36
In conclusion, the authors preliminarily revealed that downregulation of NRAGE sensitizes HuH6 cells to IR in vitro and in vivo experiments through regulating the expression of RNF8 and BARD1, and NRAGE is required for RAD51-mediated HR in HB cells. Their results provide a promising molecular targeting therapeutic strategy for HB patients.
Footnotes
Acknowledgment
The authors are thankful for Shanghai Tenth People's Hospital for providing laboratory and experiment apparatuses.
Authors Contributions'
L.L. and Z.C. performed most of experiments, analyzed the data, and wrote the first draft of the article. J.Z., J.W., S.G., J.M., H.C., and L.H. performed a part of experiments. Y.S. and J.Y. designed the experiments and provided the funding. All the authors have reviewed and approved of the article prior to submission.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
The funding support was received from Municipal Human Resources Development Program for Outstanding Leaders in Medical Disciplines in Shanghai (2017BR036); Shanghai Municipal Education Commission-Gaofeng Clinical Medical Ground Support (20171926); Shanghai Pudong Minsheng Funding (PKJ2017-Y03); Tibet Autonomous Region Natural Science Foundation (2016ZR-QY-09); The Fundamental Research Funds for the Central Universities; Hongkou District Health Planning Commission (Hongwei 1703-14); and Shanghai Municipal Health and Family Planning Research Fund (No. 201840261).