
Abstract
Background:
Recently, signal peptide-CUB-EGF-like domain containing protein 3 (SCUBE3) has been found to be associated with the development of several cancers. However, the biological role of SCUBE3 in breast cancer progression has not been reported.
Materials and Methods:
Western blotting and quantitative real-time polymerase chain reaction were used to measure the expressions of SCUBE3, TGF-β (transforming growth factor-β) signaling pathway markers, and epithelial–mesenchymal transition markers. The influence of SCUBE3 on the breast cancer cell proliferation, invasion, and migration was detected using methyl thiazolyl tetrazolium, wound healing, colony formation, and transwell assay. The role of SCUBE3 in vivo was confirmed using tumor xenograft experiment.
Results:
SCUBE3 expression was markedly increased in breast cancer cells and tissues. Knockdown of SCUBE3 suppressed cell growth, invasion, and migration, while SCUBE3 overexpression promoted cell growth, invasion, and migration in breast cancer cells. In addition, TGF-β1 and its downstream proteins were positively regulated by SCUBE3, and the promotion effect on TWIST1 expression induced by pcDNA3.1-SCUBE3 can be reversed by TGF-β1 inhibitor in breast cancer cell lines. Moreover, silencing of SCUBE3 suppressed breast cancer cell growth and tumorigenesis through reducing TGF-β1 in vivo.
Conclusion:
Knockdown of SCUBE3 downregulated TGF-β1 and TWIST1 expression, thereby inhibiting breast cancer cell growth and tumorigenesis.
Introduction
Breast cancer is the second most common cancer in the world, with a global incidence of 16.7 million new breast cancer cases in 2012. 1 According to the American Cancer Society, there were more than 231,000 new cases of breast cancer in women in the United States in 2015 and about 2350 new cases of breast cancer in men. 2 In recent years, with the continuous research on the biological functions of some important oncogenes and tumor suppressor genes, the molecular mechanism of breast cancer development has been gradually explained. 3 Some progress has been made in the diagnosis and treatment of breast cancer, but the prognosis is still poor due to the characteristics of distal recurrence and metastasis after operation. Thus, a better understanding of the molecular mechanism in breast cancer development and the discovery of the marker oncogenes in breast cancer growth and metastasis are particularly important for finding new treatments for metastatic and recurrent breast cancer.
Signal peptide-CUB-EGF-like domain containing protein 3 (SCUBE3), a member of the SCUBE gene family, is located on chromosome 6p21.3. 4 SCUBE3 is a secretory cell surface glycoprotein, which was first discovered in human umbilical vein endothelial cells. 5 SCUBE3 is mainly expressed in the normal tissues in central nervous system, gonads, vascular endothelium, heart, bone, and so on. 6 It was reported that SCUBE3 was highly expressed in lung cancer, and the tumor size, tumor metastasis, and poor prognosis were associated with its increased expression. 7 In addition, SCUBE3 high expression is closely associated with osteosarcoma prognosis. 8 However, whether SCUBE3 participates in the occurrence and development of breast cancer is still unclear.
Previous research has found that the role of SCUBE3 in lung cancer may be relevant to transforming growth factor-β (TGF-β) signaling pathway. 9 The present study found that SCUBE3 could regulate TGF-β1 expression in breast cancer. As a member of TGF-β signaling, TGF-β1 is a multifunctional protein, which can regulate cell growth, differentiation, immune regulation, and inflammation. 10,11 TGF-β1 has been proved to play a vital role in breast cancer progress. 12 Inhibition of TGF-β1 expression can repress breast cancer cell growth and migration. 13 Based on the above findings, the authors speculate that the relationship between SCUBE3 and TGF-β1 is related to the progression of breast cancer.
Hence, this study investigated the expression and the underlying mechanisms of SCUBE3 in breast cancer, aiming to find new targets for breast cancer treatment.
Materials and Methods
Patients' samples
Thirty tissue samples of breast cancer (tumor group) and 30 adjacent tissue samples (normal group) were obtained from Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology. No participants received preoperative chemotherapy or radiotherapy. The tissues were stored at −80°C for further analysis. Written informed consents of all participants had been obtained. The study was consistent with World Medical Association Declaration of Helsinki and approved by the Ethics Committee of Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology.
Cell lines and culture
Breast cancer cell lines (MDA-MB-468, HTB-132™; MDA-MB-231, HTB-26™; and MCF7, HTB-22™) and human breast fibroblast cell line (CCD-1095Sk, CRL-2122™) were purchased from ATCC (Manassas, VA). MCF-7 cell lines were maintained in RPMI-1640 medium. MDA-MB-468, MDA-MB-231, and CCD-1095Sk cell lines were cultured in DMEM. All the mediums contained 100 U/mL penicillin, 10% fetal bovine serum (FBS), and 100 μg/mL streptomycin (all from Gibco, Waltham, MA). Cells were grown in 5% CO2 incubator under 37°C.
Cell transfection
SCUBE3 knockdown plasmid (shSCUBE3#1, shSCUBE3#2) and its negative control (shNC), as well as pcDNA3.1-SCUBE3 (SCUBE3 overexpression plasmid) and pcDNA3.1-NC (the negative control plasmid), were purchased from GenePharma (Shanghai, China). SB525334 (an inhibitor of TGF-β1 14 ) was purchased from Abmole Bioscience, Inc. (Houston, TX). Cell transfections were performed using Lipofectamine 2000 (Invitrogen, Carlsbad) following the manufacturer's protocol. The cells were harvested 24 h after transfection.
Quantitative real-time polymerase chain reaction
TRIzol reagent (Invitrogen) was used to extract total RNAs from cells or tissues following the manufacturer's instructions. Reverse Transcriptase Kit (Takara, Otsu, Japan) was used to reverse transcribe cDNA. SYBR Green PCR Master Mix (Takara) with the ABI StepOnePlus Real-Time PCR system (Applied Biosystems, Foster City, CA) was used for quantitative real-time polymerase chain reaction (qRT-PCR). SCUBE3 mRNA expression was calculated relative to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) using the 2−ΔΔCt method. Each sample was performed thrice. The sequence of primers for SCUBE3 and GAPDH is listed in Table 1.
List of Primer Sequences Used in the Study
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; SCUBE3, signal peptide-CUB-EGF-like domain containing protein 3.
Immunohistochemistry assay
The clinical tissues or the xenograft tumor tissues were collected for immunohistochemistry (IHC) assay. After fixing in 10% formaldehyde and embedding in paraffin, the tissues were cut into thin slices of 4 μm thickness. The sections were then cultured with primary antibodies, including anti-SCUBE3 antibody (1:200; Abcam, Cambridge, MA, UK), anti-Ki-67 antibody (1:1000; Abcam), and anti-TWIST1 antibody (1:200; Abcam) at 4°C overnight. Next day, the sections were developed using DAB Plus Kit after incubating with the secondary HRP-conjugated anti-Rabbit antibody at room temperature for 1 h. An Olympus microscopy was used to capture the stained images.
Western blotting
Total protein was extracted from cells or tissues using RIPA lysis buffer (Beyotime, Beijing, China), and protein concentrations were measured using a BCA Protein Assay Kit (Beyotime) following the manufacturer's protocols. The protein samples were then separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) (12%) and transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore). Next, 5% nonfat milk was used to block the membranes. After being cultured for 2 h at room temperature, the membranes were incubated with primary antibodies, including anti-SCUBE3 antibody (1:1000; Abcam), anti-γ-catenin antibody (1:1000; Abcam), anti-E-cadherin antibody (1:2000; Abcam), anti-N-Fibronectin antibody (1:1000; Abcam), anti-N-cadherin antibody (1:1000; Abcam), anti-p-Smad2 antibody (1:1000; Abcam), anti-Smad2 antibody (1:2000; Abcam), anti-MMP-2 antibody (1:1000; Abcam), anti-MMP-9 antibody (1:1000; Abcam), anti-TWIST1 antibody (1:1000; Abcam), and anti-GAPDH antibodies (1:2000; Abcam), at 4°C overnight. Next day, the membranes were cultured with anti-human or anti-rabbit horseradish peroxidase-conjugated secondary antibody (1:2000; Santa Cruz) at room temperature for 1 h. The enhanced chemiluminescence detection system (Roche Molecular Biochemicals) was used to measure the blots.
Methyl thiazolyl tetrazolium assay
Methyl thiazolyl tetrazolium (MTT) assay was used to measure the cell viability. First, cells were harvested and seeded in 96-well plates (1 × 103 cells/mL in each well). When confluence reached 80%, MTT solution (15 μL, 5 mg/mL, Sigma, St. Louis) was added into each well. After incubating at 37°C for 4 h, 150 μL DMSO was added into each well to dissolve the formazan crystal. A micro-plate reader (BioTek, Winooski) was used to determine the optical density at 490 nm.
Colony formation assay
Cell proliferation was detected by colony formation assay. First, cells were seeded into six-well plates and cultured for 14 d. After being fixed with formaldehyde (10%) for 30 min, the colonies were stained with crystal violet (0.5%) for 5 min. A camera (Olympus, Tokyo, Japan) was used to photograph the colonies.
Wound-healing assay
First, cells were harvested and plated into six-well culture plates. After growing to 80% confluence, a 10 μL Eppendorf tip was used to scratch cell monolayer. Olympus microscope (10 × 10) was used to record the scratch wounds in the same position at 0 and 24 h. ImageJ software was used to quantify the denudation area. The experiment was repeated in triplicate. The formula for wound closure is as follows: (Original width—width of actual wound at 24 h)/Original width × 100%.
Transwell assay
The ability of cell invasion was determined by transwell assay. Cells seeded into the Matrigel-coated upper chamber were maintained in FBS free medium. The medium containing FBS (10%, 600 μL) was added in the lower chamber. Twenty-four hours later, the invaded cells were fixed with 4% paraformaldehyde and stained with 0.5% crystal violet for 10 min. Next, cells were counted in five random visual fields under an Olympus microscope (40 × 10) to evaluate the invasive capacity of breast cancer cells.
Enzyme-linked immunosorbent assay
TGF-β1 levels in cells or tissues were detected using a TGF-β1 ELISA Kit (Invitrogen) following the manufacturer's instructions. An enzyme-labeled colorimeter was used to determine the absorbance at 450 nm.
Tumor xenograft experiments
Animal experiments were approved by the Ethics Committee of Liyuan Hospital of Tongji Medical College of Huazhong University of Science and Technology and performed following the Guide for the Care and Use of Laboratory Animals. BALB/c nude mice (n = 30, 5–6 weeks old; Vital River Laboratory, Beijing, China) were kept under sterile specific pathogen-free facility. Xenograft mouse model was used to evaluate the tumorigenic effects in vivo. MDA-MB-231 cells (1 × 105) stably transfected with Ad-shNC or Ad-shSCUBE3 were subcutaneously injected into the buttocks of the mice. The formula for tumor volume is as follows: V = 0.5 × length × width2. Tumor size was detected every 7 d. Mice were sacrificed for the further analysis after 5 weeks.
Statistical analysis
The measurement data were presented as mean ± standard deviation. GraphPad Prism 5.0 (GraphPad Software, La Jolla, CA) and SPSS 19.0 (SPSS, Chicago, IL) were used for graphical analysis and statistics. The difference between two groups was compared using Student's t-test, and one-way analysis of variance (ANOVA) followed by Bonferroni post hoc test was performed to evaluate the statistical significance in multiple group comparisons. All assays were performed at least in triplicate. p < 0.05 was considered to have statistical significance.
Results
SCUBE3 expression in breast cancer tissues and cell lines was increased
The authors first examined the expressions of SCUBE3 in breast cancer tissues by qRT-PCR to verify the relationship between SCUBE3 and breast cancer. Compared with the normal tissues, SCUBE3 expressions in breast cancer tissues were increased (Fig. 1A). Then, SCUBE3 expression was determined using IHC assay. The results showed that the yellowish-brown area is larger in Tumor group than Normal group (Fig. 1B). In addition, the correlation between SCUBE3 expression and clinicopathological parameters of breast cancer patients was analyzed. The details are shown in Table 2. The strengthened expression of SCUBE3 was positively correlated with tumor size (T3-4), metastasis, and advanced stage (III–IV) in patients (Table 2). Moreover, SCUBE3 expression was also upregulated in breast cancer cell lines (MDA-MB-231, MDA-MB-468, and MCF7) compared to human breast fibroblast cell line (CCD-1095Sk, Fig. 1C). Because the expression of SCUBE3 was slightly lower in MDA-MB-468 cells than MDA-MB-231 cells, the MDA-MB-231 and MCF7 cell lines were selected for the further experiments (Fig. 1C). These data suggested that SCUBE3 expression was increased in breast cancer cell lines and tissues, indicating high carcinogenicity in patients with breast cancer.
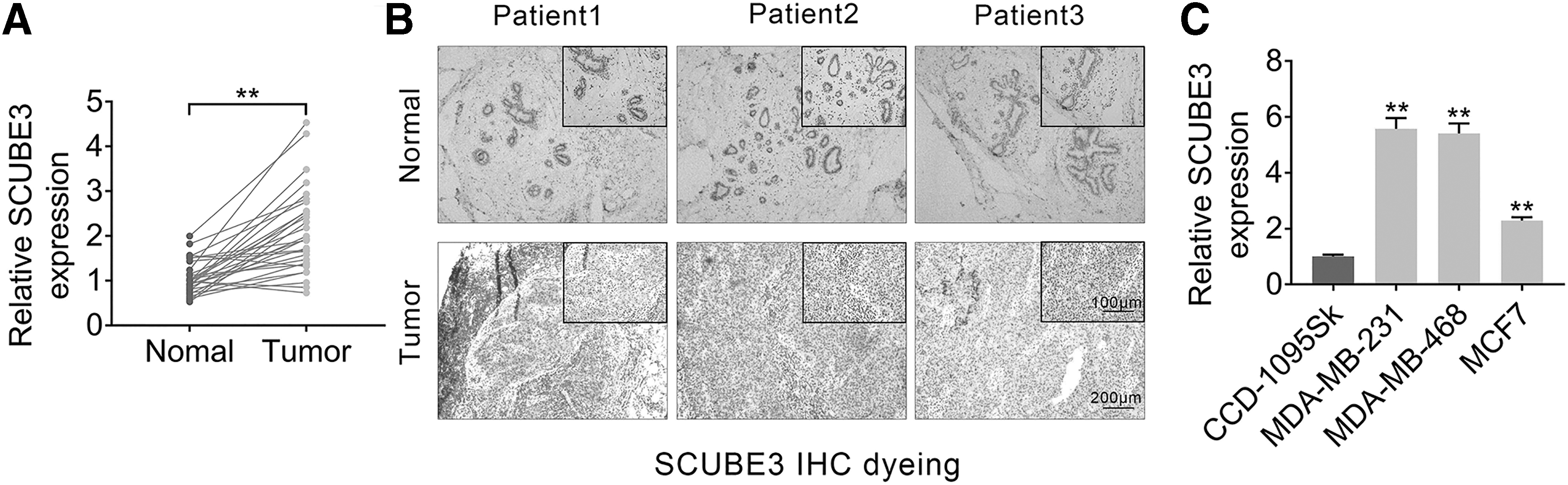
SCUBE3 expression was upregulated in breast cancer tissues and cell lines. To verify the relationship between SCUBE3 and breast cancer, we measured SCUBE3 expression in breast cancer tissues and cell lines.
Correlation Between SCUBE3 Expression and Clinical Features (n = 30)
Low/high by the sample median. Pearson χ 2 test. * p < 0.05 was considered statistically significant.
The effect of SCUBE3 expression changes on breast cancer cell proliferation
Next, function loss and enhancement experiments were conducted to test the role of SCUBE3 in MDA-MB-231 and MCF7 cell lines. To check the knockdown efficiency, two shRNAs of SCUBE3 were transfected into MDA-MB-231 cells. The results showed that shSCUBE3#1 and shSCUBE3#2 significantly downregulated the mRNA expression of SCUBE3 in MDA-MB-231 cells (Fig. 2A). The knockdown efficiency between shSCUBE3#1 and shSCUBE3#2 was similar (Fig. 2A). In addition, pcDNA3.1-SCUBE3 was transfected into MCF7 cells to increase SCUBE3 expression. As shown in Figure 2A, the mRNA expression of SCUBE3 was remarkably upregulated in pcDNA3.1-SCUBE3 group compared to pcDNA3.1-NC group. As expected, the protein expression of SCUBE3 showed the same change in trend as its mRNA expression in same groups (Fig. 2B). MTT assay suggested that silencing of SCUBE3 decreased cell viability in MDA-MB-231 cells, while overexpression of SCUBE3 enhanced cell viability in MCF7 cells (Fig. 2C). Colony formation assay demonstrated that knockdown SCUBE3 inhibited the colonies number of MDA-MB-231 cells, but strengthened SCUBE3 expression elevated the colonies number of MCF7 cells (Fig. 2D). These data revealed that SCUBE3 positively regulated breast cancer cell growth.

The effect of SCUBE3 expression changes on breast cancer cell proliferation. MDA-MB-231 cells were transfected with shRNAs of SCUBE3 (shSCUBE3#1 and shSCUBE3#2). MCF7 cells were transfected with pcDNA3.1-SCUBE3.
The effect of SCUBE3 expression changes on breast cancer cell migration and invasion
Furthermore, the ability of cell migration and invasion influenced by SCUBE3 expression was detected by wound-healing assay and transwell assay. As shown in Figure 3A and B, MDA-MB-231 cells transfected with shSCUBE3 suppressed the ability of cell migration and invasion, whereas MCF7 cells transfected with pcDNA3.1-SCUBE3 enhanced the abilities. In addition, the expressions of epithelial–mesenchymal transition (EMT)-related proteins were measured by Western blotting. Interference of SCUBE3 upregulated the expressions of epithelial marker γ-catenin and E-cadherin, but downregulated mesenchymal marker Fibronectin and N-cadherin in MDA-MB-231 cells compared to shNC group (Fig. 3C). Meanwhile, the expressions of γ-catenin and E-cadherin were decreased, and the expressions of Fibronectin and N-cadherin were increased in MCF7 cells transfected with pcDNA3.1-SCUBE3 compared to pcDNA3.1-NC (Fig. 3C). Altogether, these data indicated that breast cancer cell migration and invasion were positively regulated by SCUBE3.
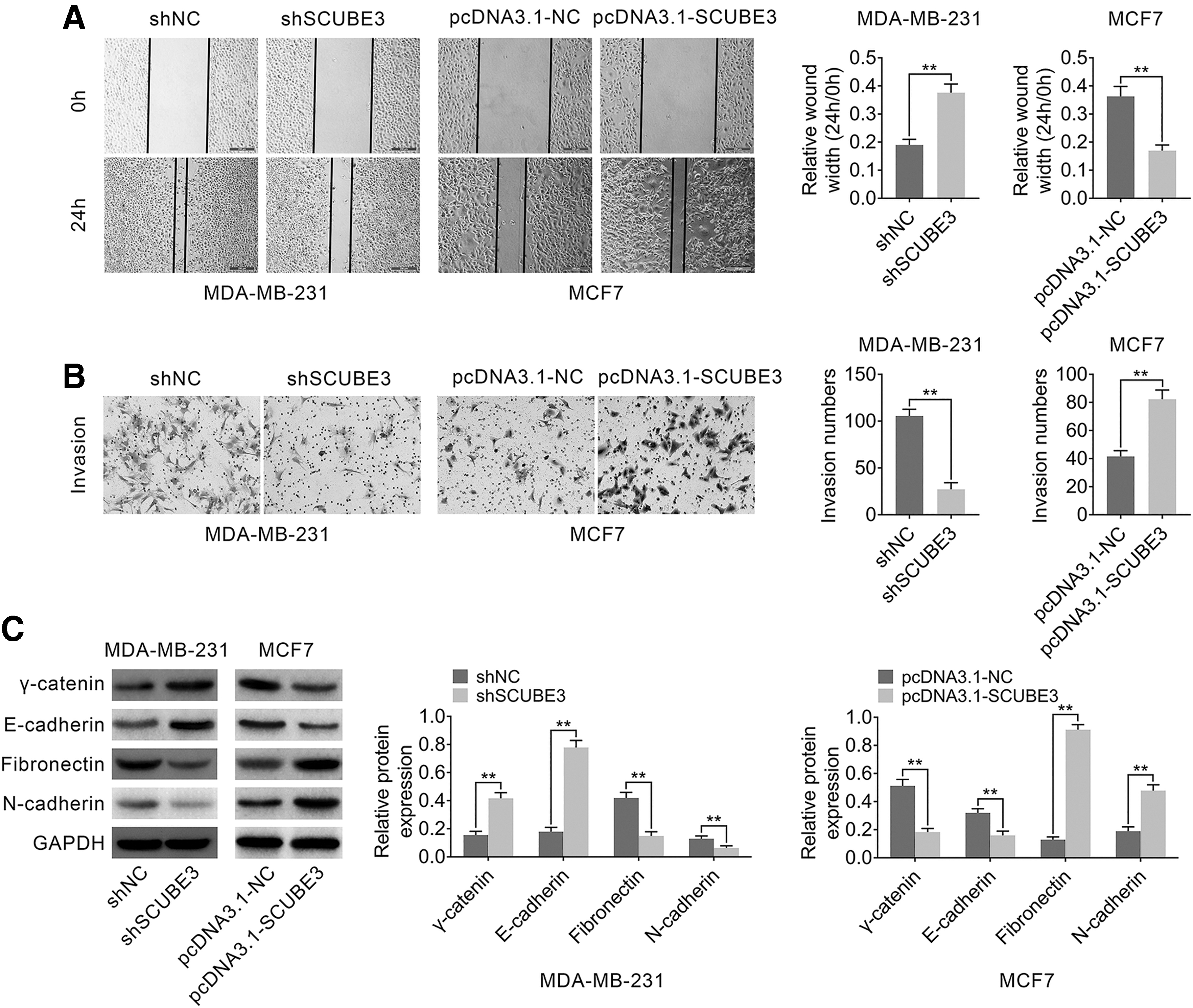
The effect of SCUBE3 expression changes on cell migration and invasion in breast cancer cell lines. MDA-MB-231 cells were transfected with shSCUBE3. MCF7 cells were transfected with pcDNA3.1-SCUBE3.
SCUBE3 positively regulated TGF-β1 expression in breast cancer cell lines
EMT is the key process of tumor metastasis, 15 and the activation of TGF-β signaling pathway is closely related to EMT. 16 In these experiments, the authors examined the effects of SCUBE3 on TGF-β signaling pathway in breast cancer cell lines. First, TGF-β1 expression was determined by enzyme-linked immunosorbent assay (ELISA), and the results suggested a low expression of TGF-β1 in MDA-MB-231 cells transfected with shSCUBE3 compared to shNC (Fig. 4A) and a high expression of TGF-β1 in MCF7 cells after overexpressing SCUBE3 (Fig. 4B). Then, Western blotting results showed that interference of SCUBE3 inhibited the expression of phosphorylation of Smad2 (p-Smad2), a recognized marker of the activated TGF-β pathway, 17 and decreased the expressions of EMT markers (including matrix metalloproteinase-2 [MMP2] and MMP9) and TWIST1 in MDA-MB-231 cells (Fig. 4A). In addition, the expression trends of these proteins were opposite in MCF7 cells treated with pcDNA3.1-SCUBE3 (Fig. 4B). Taken together, these results suggested that TGF-β1 and its downstream proteins were positively regulated by SCUBE3.
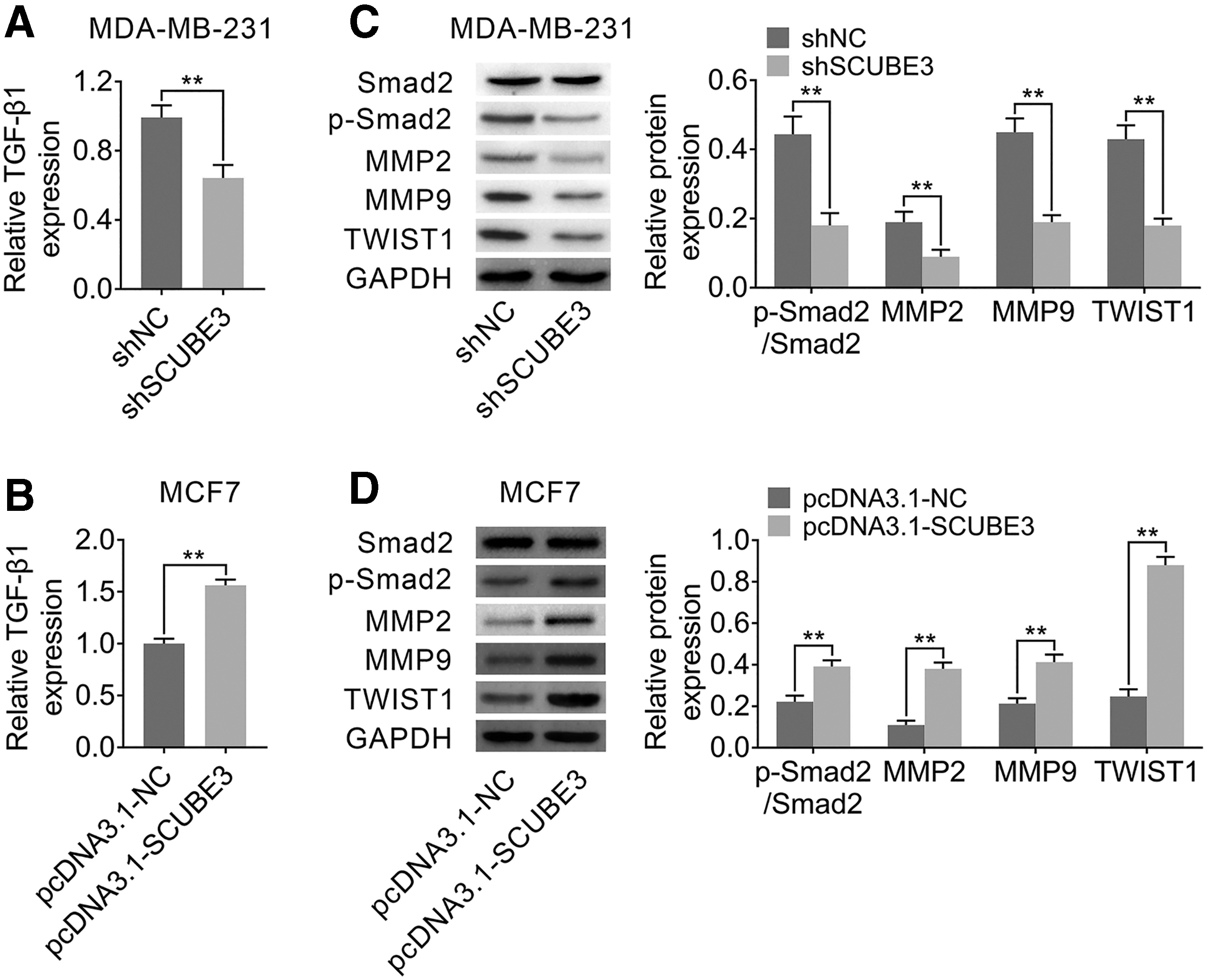
SCUBE3 regulated TGF-β1 expression in breast cancer cell lines.
SCUBE3 positively regulated TWIST1 through TGF-β1
To detect the mechanism of SCUBE3, pcDNA3.1-SCUBE3 and SB525334 (an inhibitor of TGF-β1) were used to treat MCF7 cells. Gene expressions of Smad2, p-Smad2, MMP2, MMP9, and TWIST1 were evaluated using Western blotting. As shown in Figure 5, overexpression of SCUBE3 upregulated the expressions of p-Smad2, MMP2, MMP9, and TWIST1, while the promotional effects were reversed by SB525334. These data demonstrated that inhibition of TGF-β1 abrogated the regulation of SCUBE3 overexpression on the downstream genes of TGF-β signaling pathway.

SCUBE3 regulated TWIST1 using TGF-β1. MCF7 cells were treated with pcDNA3.1-SCUBE3 and SB525334 (an inhibitor of TGF-β1). Gene expressions of Smad2, p-Smad2, MMP2, MMP9, and TWIST1 were evaluated using Western blotting. **p < 0.01 versus pcDNA3.1-NC or pcDNA3.1-SCUBE3.
Knockdown of SCUBE3 inhibited breast cancer cell tumorigenesis by reducing TGF-β1 in vivo
Finally, the effect of SCUBE3 on breast cancer cell tumorigenesis in vivo was evaluated using xenograft mouse models. MDA-MB-231 cells stably transfected with Ad-shSCUBE3 or Ad-shNC were subcutaneously injected into the buttocks of mice. The tumor size in Ad-shSCUBE3 group was smaller than Ad-shNC group (Fig. 6A). As expected, TGF-β1 expression was downregulated in shSCUBE3 group compared with Ad-shNC group, confirmed by ELISA (Fig. 6B). Western blotting results showed that the protein levels of SCUBE3, p-Smad2, MMP2, MMP9, and TWIST1 in vivo were decreased in Ad-shSCUBE3 group compared to Ad-shNC group (Fig. 6C). Moreover, IHC assay was performed to check the expression of Ki67 (a proliferative indicator), SCUBE3, and TWIST1. As shown in Figure 6D, the levels of Ki67 SCUBE3 and TWIST1 in Ad-shSCUBE3 group were markedly lower than that in Ad-shNC group. The above data revealed that silencing of SCUBE3 suppressed breast cancer cell growth and tumorigenesis through TGF-β1 in vivo.
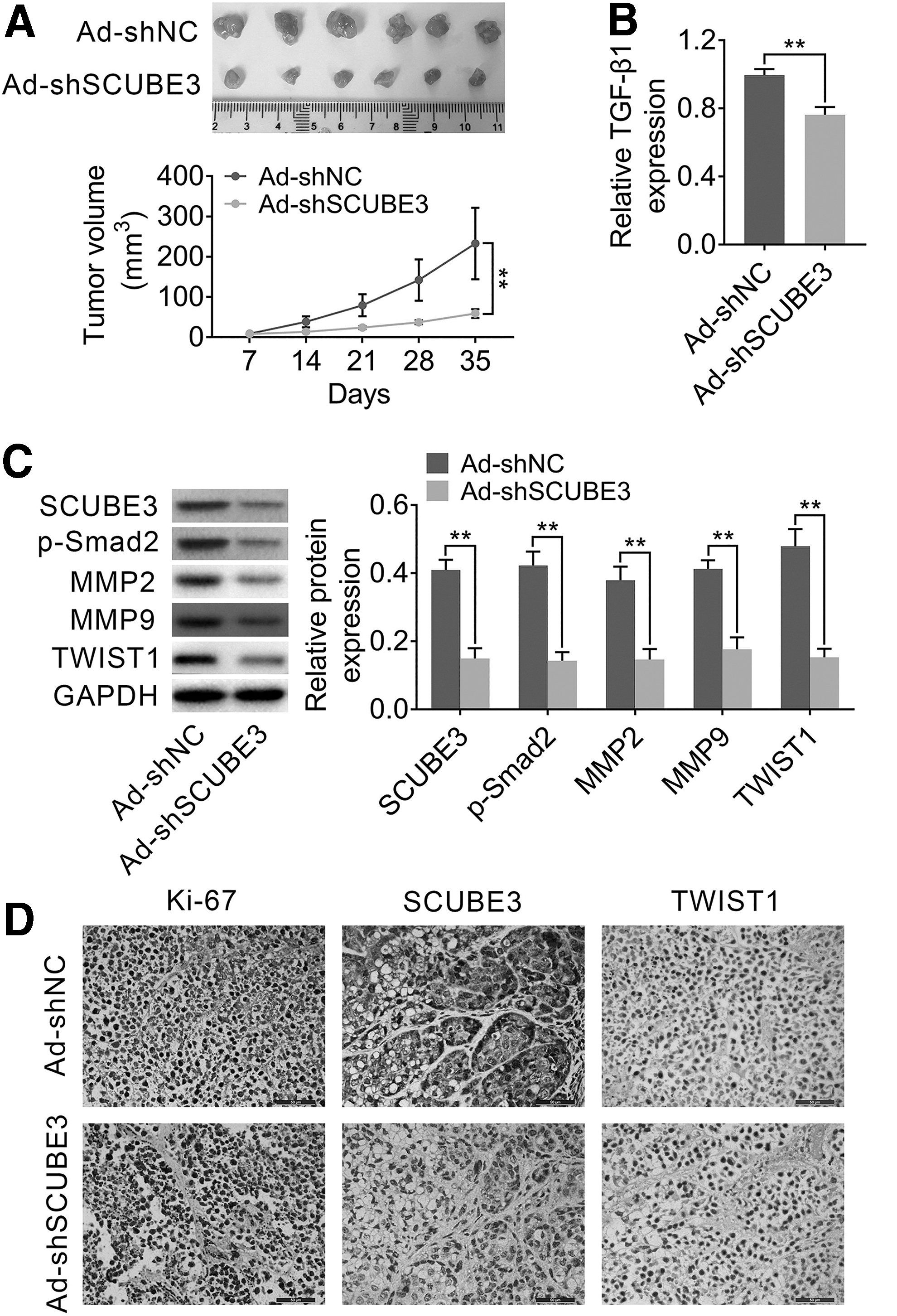
Knockdown of SCUBE3 inhibited breast cancer cell tumorigenesis by reducing TGF-β1 in vivo. MDA-MB-231 cells stably transfected with Ad-shSCUBE3 or Ad-shNC were subcutaneously injected into the buttocks of BALB/c nude mice.
Discussion
Breast cancer is one of the most common malignant tumors in women, and its incidence and mortality show ascending tendency in recent years. 18 In addition, the study has found that the incidence of breast cancer appears to be a younger trend. 19 Hence, there is an urgent need to discover effective biomarkers and treatments for breast cancer. In their study, the authors focused on SCUBE3. The authors first found that SCUBE3 was strongly expressed in human breast cancer cell lines and tissues, and knockdown of SCUBE3 could suppress breast cancer cell growth, invasion, and migration. The authors also revealed that the role of SCUBE3 in the occurrence and development of breast cancer was related to TGF-β1 and TWIST1.
SCUBE3 was originally identified by the transcription analysis of vascular endothelial cells and showed obvious enrichment in primitive osteoblasts and long bones. 20,21 SCUBE3 can be expressed in multiple tissues during development. 22 Recent studies have found that SCUBE3 is associated with the development of several cancers. For instance, SCUBE3 was highly expressed in lung cancer tissues, and SCUBE3 knockdown inhibited the early lung cancer angiogenesis and metastatic progression 23 ; SCUBE3 was strongly expressed in osteosarcoma tissues and cells, and silencing of SCUBE3 reduced the proliferation of osteosarcoma cells. 8 These previous studies reveal that SCUBE3 is an oncogene. However, whether SCUBE3 is involved in breast cancer pathological process has not been reported. In the current study, the authors find that SCUBE3 has high carcinogenicity in patients with breast cancer. Furthermore, SCUBE3 can induce breast cancer cell growth, invasion, and migration. This is the first time that SCUBE3 has been found to participate in breast cancer development. Hence, it is of great significance to research the role of SCUBE3 in breast cancer.
Previous research has found that SCUBE3 can activate TGF-β signaling, thus triggering EMT and promoting the lung cancer progression. 9 In the current study, the authors also found that SCUBE3 could positively regulate TGF-β signaling pathway in breast cancer cells. TGF-β signaling pathway is an important intracellular signal transduction pathway, and mounting evidence has demonstrated that TGF-β signaling pathway is closely related to tumor development, 24,25 including breast cancer. Tang and his colleagues suggested that SIRT7 deficiency promoted breast cancer metastasis through activating TGF-β signaling. 26 Wang et al. demonstrated that blocking TGF-β suppressed the invasiveness, migration, and angiogenesis of breast cancer cells through ERK/S100A4 signal. 27 In addition, studies have showed that the role of TGF-β signaling pathway in breast cancer is related to EMT. 28 It was reported that TGF-β pathway played a vital role in triple-negative breast cancer epirubicin resistance through regulating EMT. 29 In this study, the authors found that SCUBE3 could positively regulate TGF-β signaling pathway and EMT in breast cancer cells, while the effect was reversed by TGF-β1 inhibitor, which indicated that the role of SCUBE3 in breast cancer was associated with TGF-β1. Moreover, the authors also found that the TWIST1 expression was affected by the changes of SCUBE3 and TGF-β1 expression. As a member of the basic helix-loop-helix transcription factor family, TWIST1 is one of the major transcription factors that induce EMT, cell migration, and invasion in cancer cells, including breast cancer cells. 30 In vivo experiments confirmed that interfering of SCUBE3 downregulated the expression of TGF-β1 and TWIST1 and suppressed breast cancer tumorigenesis. However, due to time and funding problems, the authors did not verify whether there is a targeting relationship among SCUBE3, TGF-β1, and TWIST1, which will be validated in subsequent experiments.
In summary, the current study demonstrates that activation of TGF-β1 by SCUBE3 regulates TWIST1 expression and promotes breast cancer progression. This is the first study to report that SCUBE3 was increased in breast cancer, and SCUBE3 played a vital role in accelerating breast cancer tumorigenesis. These findings proposed that SCUBE3 might serve as a key regulatory hub and a novel target for breast cancer treatment.
Footnotes
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Authors' Contributions
X.H.Y. and J.D. conceived and designed the experiments, J.Q.H. analyzed and interpreted the results of the experiments, and C.C.S. performed the experiments.
Ethics Approval and Consent to Participate
The animal use protocol listed below has been reviewed and approved by the Animal Ethics and Welfare Committee.
Informed Consent
Written informed consent was obtained from a legally authorized representative(s) for anonymized patient information to be published in this article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.