
Abstract
Background:
Hepatocellular carcinoma (HCC), the fourth leading cause of cancer-related deaths worldwide, has increased public concern. Data from previous work have validated that long noncoding RNAs are active participators in the malignant processes of a host of cancers. Small nucleolar RNA host gene 7 (SNHG7) has been revealed to act as a tumor promoter in several cancers and SNHG7 inhibition was revealed to suppress cell invasion in HCC. Nevertheless, the specific role of SNHG7 in HCC deserves deeper exploration.
Aim of the Study:
This work aimed to uncover the role and the regulatory mechanisms of SNHG7 in HCC.
Materials and Methods:
The expression of SNHG7 and cyclin mediator 1 (CNNM1) in HCC cells were analyzed by quantitative real-time polymerase chain reaction. The influences of SNHG7 on HCC occurrence were studied by cell counting kit-8 (CCK-8), colony formation, flow cytometry analysis, and Western blot assays. Luciferase reporter assay or RNA immunoprecipitation assay was conducted to confirm the relationship between miR-9-5p and SNHG7 (or CNNM1).
Results:
SNHG7 was overexpressed in HCC tissues and cell lines. SNHG7 facilitated cell proliferation, while suppressed cell apoptosis in HCC. Moreover, miR-9-5p expression was negatively modulated by SNHG7 and therefore was downregulated in HCC cells. We also found that CNNM1 existed in miR-9-5p induced RNA-induced silencing complex and a series of assays verified that CNNM1 acted as the target gene of miR-9-5p. Consequently, the messenger RNA and protein level of CNNM1 were detected to be inversely regulated by miR-9-5p. Moreover, rescue assays demonstrated that CNNM1 overexpression could countervail the SNHG7 depletion-mediated cellular functions of HCC cells.
Conclusions:
SNHG7 sponges miR-9-5p to upregulate CNNM1 in promoting HCC progression.
Introduction
As the fourth cause of cancer-related deaths worldwide, hepatocellular carcinoma (HCC) has attracted the attention of researchers. 1 HBV, HCV infections, and alcohol intake may lead to the occurrence of HCC. 2 The enhanced cell growth and metastasis are distinct features of HCC. 3
Despite radiotherapy, chemotherapy and surgical resection (including liver resection and liver transplant) have been applied into the treatment of HCC sufferers, 4 the 5-year survival rate is still unsatisfactory, 1 which is largely attributed to the frequent metastatic behaviors and common recurrence. 5 Although some HCC-associated genes have been uncovered in regulating HCC progression at molecular level, 6 the functions and potential mechanisms of majority molecular in HCC still remain elusive. Deep investigation into the molecular mechanisms underlying the malignant behaviors of HCC will shed new light into the HCC pathology, therefore may offer novel target for the treatment of HCC patients.
It has been well established that noncoding RNAs (ncRNAs) play a crucial part in the processes of a variety of diseases and tumors. 7,8 ncRNAs have been confirmed to lack of protein-coding capability, which could be divided into long noncoding RNAs (lncRNAs) and microRNAs (miRNAs) according to different nucleotide lengths, lncRNAs are depicted as highly conserved ncRNAs with a length of over 200 nucleotides. 9 Accumulating studies uncovered that lncRNAs exert profound effects in the regulation of a wide array of malignant tumors, 10,11 including HCC. 12,13 A large body of studies manifested that there exists an appealing reciprocal modulation between lncRNAs and miRNAs. 14,15 More importantly, lncRNAs are able to serve as competing endogenous RNAs (ceRNAs) to sponge specific miRNAs, thus abolishing the repression of miRNA on messenger RNAs (mRNAs) expression and cellular functions in the development of cancers. 16 17 LncRNA small nucleolar RNA host gene 7 (SNHG7) has been revealed to function as an oncogenic gene in several cancers. 18,19 SNHG7 inhibition was revealed to suppress cell invasion in HCC. 20 Nevertheless, the specific role and mechanism of SNHG7 in HCC deserve further investigation.
This work was designed to uncover the role and the regulatory mechanisms of SNHG7 in HCC. The authors' research data supported that SNHG7 antagonizes the interaction of miR-9-5p with cyclin mediator 1 (CNNM1) and therefore facilitates CNNM1-induced cell proliferation in HCC.
Materials and Methods
Tissue samples
Total HCC tissue samples and adjacent normal tissues were acquired from recruited patients who did not undergo any preoperative chemotherapy or radiation therapy. HCC tissues and adjacent normal tissues were preserved at −80°C in liquid nitrogen till RNA extraction. All patients were informed and signed the written informed consent before the study. This study was approved and supervised by the Ethics Committee of Eastern Hepatobiliary Surgery Hospital, Second Military Medical University.
Cell culture
Normal human liver cell (THLE-3) and human HCC cells (BEL-7404, HCCLM3, Hep3B and HepG2) were all acquired from the Shanghai Cell Bank of the Chinese Academy of Sciences (Shanghai, China). Cells were incubated at 37°C under humidified condition with a constant 5% CO2 air flow. Cells were propagated with Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Waltham, MA) with additional 1% penicillin/streptomycin (Thermo Fisher Scientific) plus 10% fetal bovine serum (Thermo Fisher Scientific). Medium was refreshed once every 3 d.
Cell transfection
Specific shRNAs against SNHG7 (sh-SNHG7#1 and sh-SNHG7#2) and their corresponding negative control (NC) were constructed by Genechem (Shanghai, China). The sequences of SNHG7 sh-NC, together with sh-RNAs, are displayed below: sh-NC: CCGGATCTTGTACTGTATACTAGATCTCGAGATCTAGTATACAGTACAAGATTTTTTG; sh-SNHG7#1: CCGGCTCAAATTTTGCCAACACAGACTCGAGTCTGTGTTGGCAAAATTTGAGTTTTTG; sh-SNHG7#2: CCGGAACACAGAAATATTATCCTTACTCGAGTAAGGATAATATTTCTGTGTTTTTTTG.
Design and synthesis of miR-9-5p mimics and miR-9-5p inhibitor, as well as NC mimics and NC inhibitor, were accomplished by GenePharma (Shanghai, China). The sequences of NC mimics, miR-9-5p mimics, NC inhibitor, and miR-9-5p inhibitor were shown below: NC mimics: UCAAACCUUAUCUAGCUGUAUGA; miR-9-5p mimics: UCUUUGGUUAUCUAGCUGUAUGA; NC inhibitor: TCATACAGCTAGATAAGGUUUGA; and miR-9-5p inhibitor: TCATACAGCTAGATAACCAAAGA.
The pcDNA3.1 vector targeting CNNM1 and the empty vector were simultaneously acquired from Genechem. Plasmids mentioned above were individually transfected into Hep3B or HCCLM3 cells by applying Lipofectamine 3000 (Invitrogen, Carlsbad, CA). Twenty nanomolar of shRNA, 50 nM of miR-9-5p mimics, or 100 nM of miR-9-5p inhibitor were transfected into cells using Lipofectamine 3000 (Invitrogen). Forty-eight hours after transfection, cells were collected and the transfection efficiency was analyzed by quantitative real-time polymerase chain reaction (RT-qPCR) analysis.
Quantitative real-time polymerase chain reaction
TRIzol® reagent purchased from Thermo Fisher Scientific was applied for isolating total RNA, which was subsequently reverse transcribed into cDNA utilizing a High Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific). After that, qPCR was carried out by using SYBR Green Reverse Transcription PCR Master mix (Takara Biotechnology, Tokyo, Japan) on an Applied Biosystems 7300 (Applied Biosystems, Foster City, CA) plus reverse transcription PCR system (Thermo Fisher Scientific). The 2−ΔΔCt cycle threshold method was used for determining expression levels. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6 were acted as internal reference genes.
Cell counting kit-8 assay
Cell counting kit-8 (CCK-8) assay was carried out to study cell proliferation. After transfection, Hep3B or HCCLM3 cells were inoculated in 96-well plates. CCK-8 solution (Dojindo Laboratories, Kumamoto, Japan) was added at indicated time points. Subsequently, each well was examined spectrophotometrically at 450 nm.
Colony formation assay
Transfected Hep3B or HCCLM3 cells were placed to six-well culture plates. Fourteen days later, cells were stained in 0.1% crystal violet solution (Sigma-Aldrich, St. Louis, MO), followed by assessing the number of colonies via a microscope (Olympus, Tokyo, Japan). Colonies with at least 50 cells were counted.
Flow cytometry analysis
Transfected Hep3B or HCCLM3 cells were planted in six-well plates and washed twice with cold phosphate-buffered saline (Sigma-Aldrich), after which were resuspended in 1 × binding buffer (Thermo Fisher Scientific). After that, cells were incubated without light for 15 min by Annexin V-FITC/PI (Invitrogen) and analyzed with the Accuri C6 flow cytometer (BD Biosciences, Mountain View, CA).
Western blot
Radioimmunoprecipitation assay lysis buffer obtained from Sigma-Aldrich was used for isolating total protein from transfected Hep3B or HCCLM3 cells. Then, qualification of total protein was conducted with the BCA Detecting Kit (Keygen, Nanjing, China).
Next, proteins samples were separated utilizing 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (Millipore, Bedford, MA) and subsequently transferred onto polyvinylidene fluoride membranes (Millipore), followed by incubation overnight at 4°C with primary antibodies, including anti-Bcl-2 antibody (ab692; Abcam), anti-Bax antibody (ab182733; Abcam), anti-cleaved-caspase-3 antibody (ab32042; Abcam), anti-total-caspase-3 antibody (ab32351; Abcam), anti-CNNM1 antibody (ab122648; Abcam), and anti-GAPDH antibody (ab8245; Abcam), respectively.
Membranes were incubated with secondary antibodies (Abcam) the next day for 1 h. Protein bands were finally analyzed on X-ray film with an enhanced chemiluminescence detection system (Santa Cruz Biotechnology, Santa Cruz, CA). GAPDH was adopted as a loading control.
Subcellular fractionation assay
For the purpose of determining the cellular distribution of SNHG7 in HCC cells, separation of the nuclear fraction of Hep3B or HCCLM3 cells from cytoplasm was implemented via a PARIS Kit (Thermo Fisher Scientific) as per the supplier's guide. RNA was separated from the nuclei and cytoplasm of Hep3B or HCCLM3 cells, and subsequently, the SNHG7 expression in the nuclear and cytoplasm was assayed via RT-qPCR. GAPDH and U6 were, respectively, utilized as the cytoplasmic and nuclear controls.
RNA pull-down assay
Hep3B or HCCLM3 cells were transfected with SNHG7 biotin probe or SNHG7 no-biotin probe. Forty-eight hours later, cells were reaped and biotin-labeled RNA was pulled down through incubating cell lysates together with Dynabeads™ M-270 Streptavidin Beads (Invitrogen). After that, purification of the bound RNA was carried out with the PureLink™ RNA Mini Kit (Invitrogen). Results were analyzed by RT-qPCR.
Luciferase reporter assay
SNHG7-WT/Mut or CNNM1-WT/Mut was inserted into the pmirGLO dual-luciferase plasmid (Promega, Madison, WI) for constructing pmirGLO-SNHG7-WT/Mut or pmirGLO-CNNM1-WT/Mut. HEK-293Tcells were subsequently cotransfected with pmirGLO-SNHG7-WT/Mut or pmirGLO-CNNM1-WT/Mut and miR-9-5p mimics or NC mimics. Relative activity was eventually evaluated via dual-luciferase reporter assay system (Promega).
RNA immunoprecipitation assay
As per the specification of the RNA-Binding Protein Immunoprecipitation Kit (Millipore), RNA immunoprecipitation assay was implemented. Anti-Ago2 antibody (Millipore) or anti-IgG antibody ((Millipore) was applied for carrying out RNA immunoprecipitation. Enrichment was determined with RT-qPCR.
In vivo experiments
BALB/c nude mice (6 weeks old) were bought from the Experiments Animal Center of Nanjing Biological Institute (Nanjing, China) and were housed under standard SPF-condition. Hep3B cells transfected with sh-NC, sh-SNHG7#1, or sh-SNHG7#1+pcDNA3.1/CNNM1 were subcutaneously injected to mice. Mice were randomly divided into three groups accordingly to different transfection conditions. Tumors volume was measured every 4 d. After 4 weeks of injection, mice were euthanized and the tumors were excised. The tumor was photographed and the weight of the collected tumors was measured in collected tumors. Animal experiments obtained the approval of the Animal Research Ethics Committee of Eastern Hepatobiliary Surgery Hospital, Second Military Medical University.
Statistical analysis
In this study, statistical analysis was carried out with GraphPad Prism 5 software (GraphPad Software, San Diego, CA). Student's t-test or one-way analysis of variance (ANOVA) was applied for estimating significance of differences between or among groups. p < 0.05 indicated statistical significances. Data were listed as mean ± standard deviation from experiments which were conducted in triplicate.
Results
SNHG7 facilitates the progression of HCC
The cancer RNA-Seq Nexus database revealed 20 lncRNAs that are upregulated in HCC (Fig. 1A). It has been reported that SNHG1/12/20 accelerates the tumorigenesis of HCC. 21 Therefore, SNHG7 was selected for further investigation. Then, RT-qPCR results manifested that SNHG7 exhibited notable higher expression in HCC cell lines (BEL-7404, HCCLM3, Hep3B, and HepG2) than normal human liver cell line THLE-3 (Fig. 1B). Later, SNHG7 was silenced in HCCLM3 and Hep3B cells for the following loss-of-function assays. The results suggested that the transfection of sh-SNHG7#1/2 indeed remarkably silenced SNHG7 expression, and sh-SNHG7#1 possessed better inhibitory efficiency of knockdown (Fig. 1C).
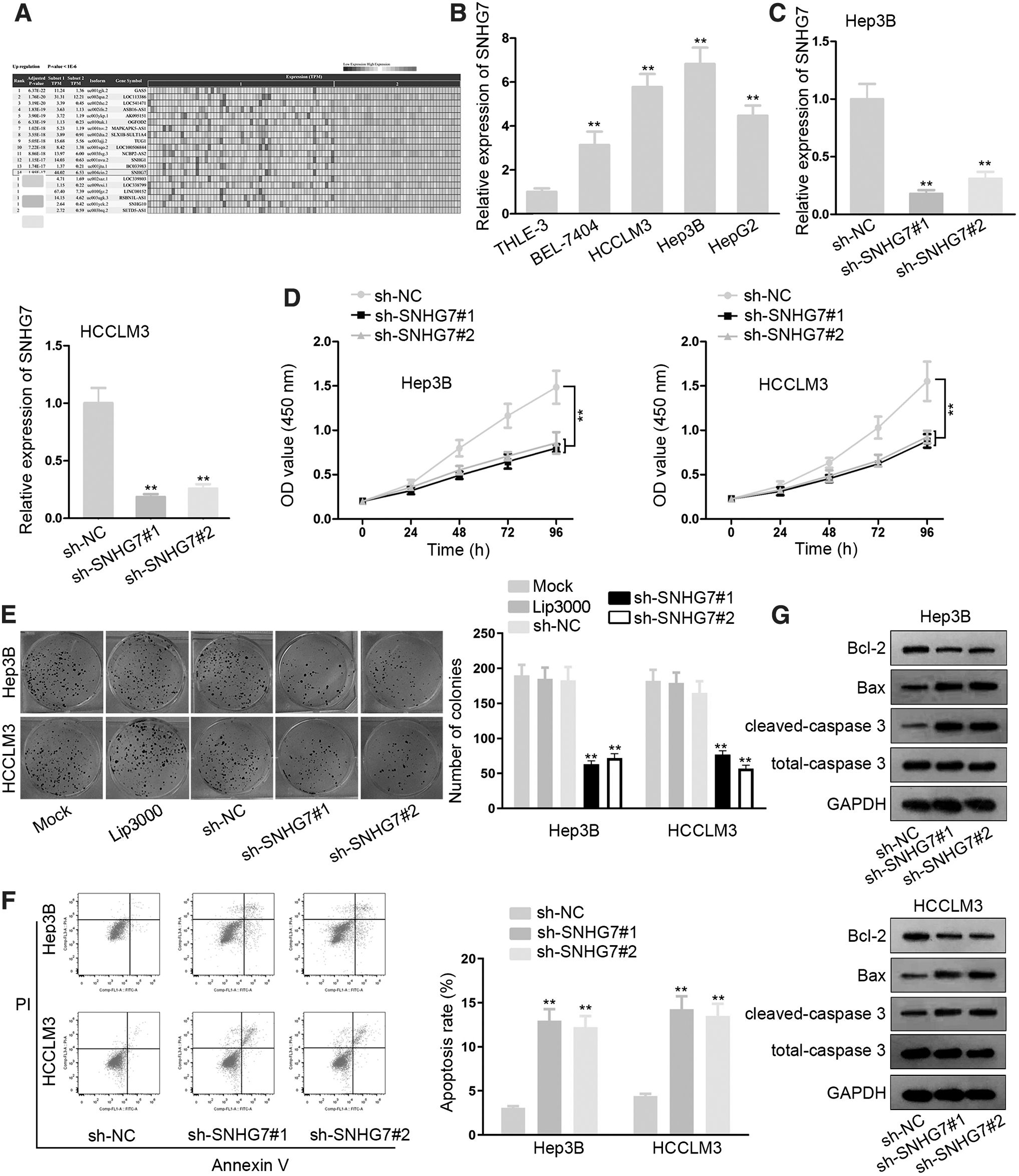
SNHG7 promotes the progression of HCC.
Subsequently, CCK-8 assays demonstrated that cell growth was suppressed in sh-SNHG7#1/2-transfected cells (Fig. 1D). Moreover, colony formation assay indicated that SNHG7 silence weakened cell proliferation ability of HCCLM3 and Hep3B cells compared with NC group, whereas results in NC group were almost the same as those in mock group (Fig. 1E). Furthermore, flow cytometry analysis showed that SNHG7 silence increased the apoptotic fraction of HCC cells, indicating the promoting effect on apoptosis of SNHG7 silence (Fig. 1F). Last but not least, Western blot assay reflected that the level of Bcl-2 was decreased while the levels of Bax and cleaved-caspase 3 were increased in response to SNHG7 silence (Fig. 1G).
All in all, SNHG7 is overexpressed in HCC. SNHG7 knockdown represses cell proliferative ability as well as facilitates cell apoptosis in HCC.
SNHG7 directly interacts with miR-9-5p
To explore the mechanism of SNHG7 in HCC, the localization of SNHG7 in HCCLM3 and Hep3B cells was first determined. Subcellular fractionation assay displayed that SNHG7 mainly localized in the cytoplasm of HCCLM3 and Hep3B cells (Fig. 2A). The starBase website was used to search potential combinable miRNAs for SNHG7. It screened out five miRNAs that may bind with SNHG7 (Fig. 2B). Subsequently, in RNA pull down assay detected that miR-9-5p was the only miRNA that significantly pulled down by SNHG7 biotin probe compared with other candidate miRNAs (Fig. 2C). Thus, miR-9-5p was chosen to be investigated.
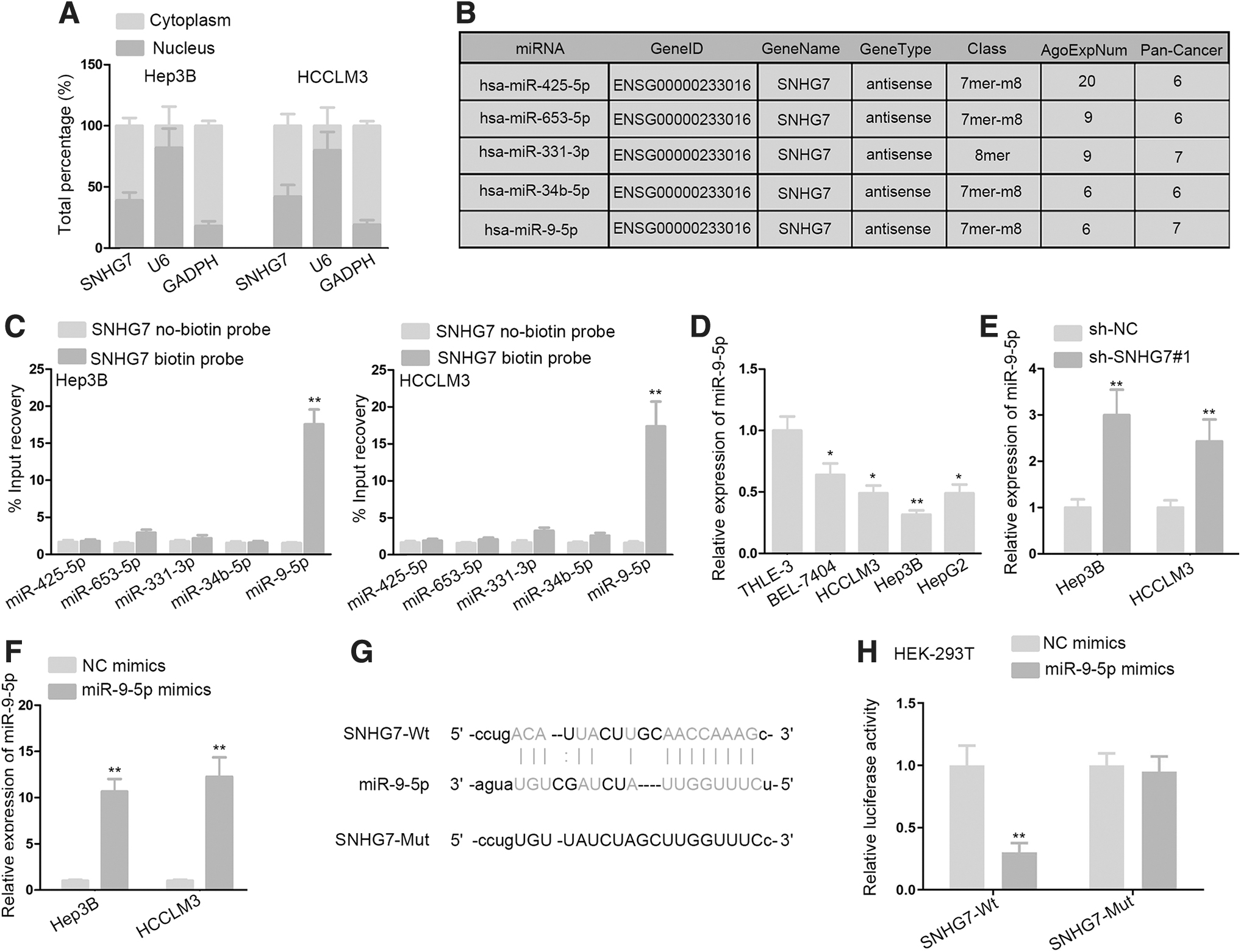
SNHG7 acts as a sponge of miR-9-5p.
RT-qPCR validated that miR-9-5p expression was considerably downregulated in HCC cells in contrast with normal THLE-3 cells (Fig. 2D). Besides, miR-9-5p expression was measured and found to be upregulated in SNHG7-depleted HCCLM3 and Hep3B cells (Fig. 2E). RT-qPCR analysis was utilized to examine the overexpression efficiency of miR-9-5p. Its results manifested that miR-9-5p was upregulated in reaction with the transfection of miR-9-5p mimics into HCCLM3 and Hep3B cells (Fig. 2F). Next, bioinformatics analysis displayed that SNHG7 had highly conserved target sites of miR-9-5p (Fig. 2G). Furthermore, luciferase activity assay data manifested that miR-9-5p upregulation markedly weakened the luciferase activity of SNHG7-Wt reporter, while that of SNHG7-Mut reporter showed no change in HEK293T cells (Fig. 2H).
To further study whether SNHG7 enhances HCC development via sponging miR-625-5p, the authors carried out an array of rescue assays. They depleted miR-625-5p expression in Hep3B and HCCLM3 cells by transfecting miR-625-5p inhibitor (Supplementary Fig. S1B). Rescue assays results revealed that miR-625-5p inhibitor could dramatically offset the effects of SNHG7 knockdown-induced proliferation (Supplementary Fig. S1C, D) and apoptosis (Supplementary Fig. S1E, F) in Hep3B and HCCLM3 cells. In conclusion, miR-9-5p is downregulated in HCC cells and sponged by SNHG7 in HCC.
CNNM1 is a downstream target of miR-9-5p
It has been proven by extensive research that mRNAs are regulated by miRNAs to exert their functions in cancer development. 22 Thus, the authors applied the starBase website and screened out three possible downstream targets of miR-9-5p (Fig. 3A). Next, RT-qPCR analyses were used to probe into the influence of miR-9-5p on these predicted targets. The results demonstrated that CNNM1 mRNA level was conspicuously downregulated by miR-9-5p overexpression, while the mRNA level of the other 2 candidates displayed no significant change (Fig. 3B).
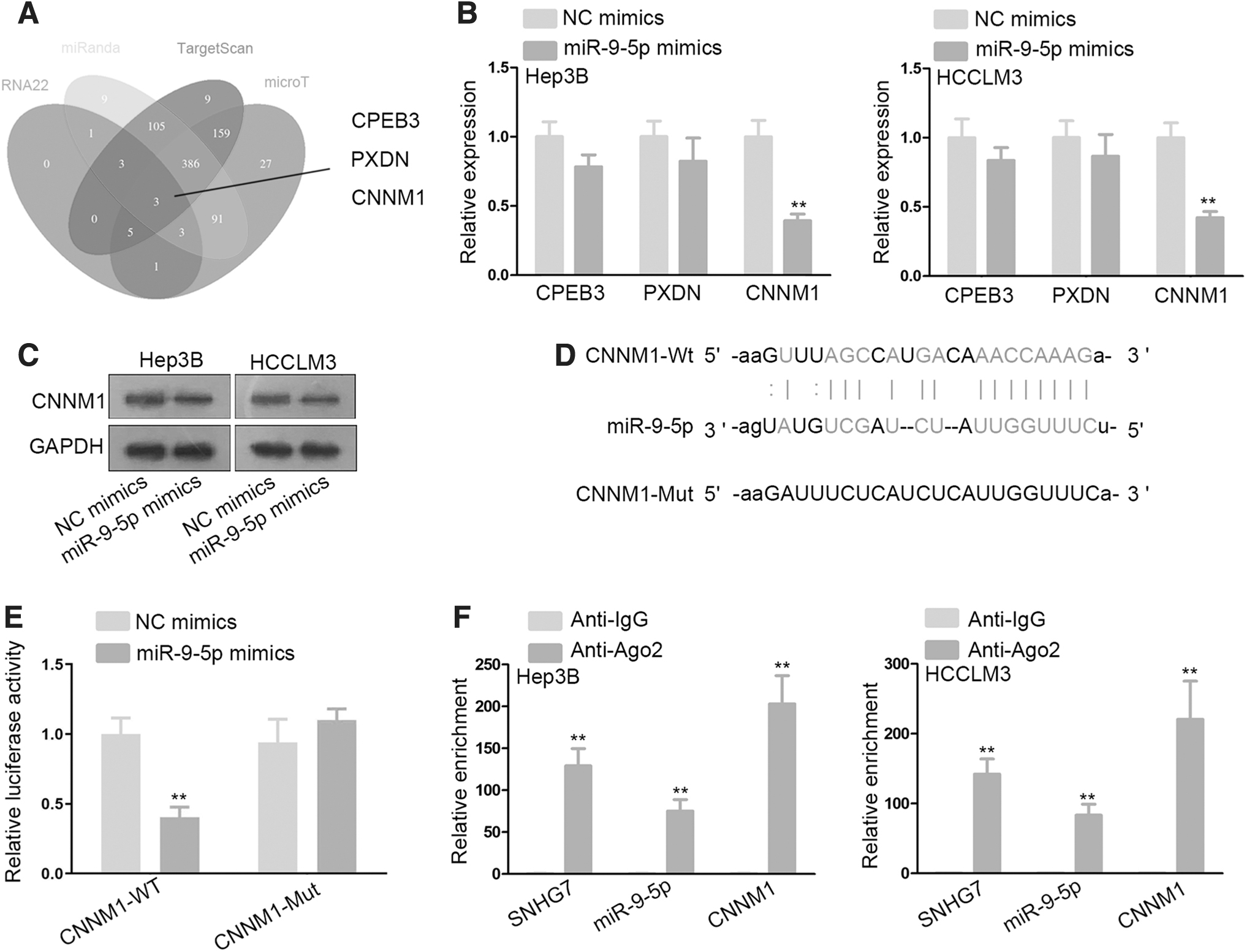
CNNM1 is a downstream target of miR-9-5p.
Then, Western blot assay showed that the protein level of CNNM1 was also depleted in reaction with miR-9-5p overexpression in selected HCC cell lines (Fig. 3C). Furthermore, they obtained the binding sites between miR-9-5p and CNNM1 by bioinformatics analysis (Fig. 3D). Subsequently, luciferase reporter assay manifested that the luciferase activity of CNNM1-WT was dramatically attenuated in HEK-293T as a result of miR-9-5p mimics cotransfection, while no luciferase activity variation of CNNM1-Mut in response to miR-9-5p overexpression, indicating that CNNM1 was negatively modulated by miR-9-5p (Fig. 3E).
Finally, an RNA immunoprecipitation assay was conducted. RNA was extracted from the protein precipitate. The data revealed evident enrichment of SNHG7, miR-9-5p, and CNNM1 in antibody targeting Ago2 group, indicating the coexistence of them in RNA-induced silencing complex (RISC) (Fig. 3F). This finding provided potent evidence for the ceRNA regulatory network involving SNHG7, miR-9-5p, and CNNM1 in HCC. In other words, CNNM1 is a downstream target of miR-9-5p and might be regulated by SNHG7 through miR-9-5p.
SNHG7 enhances HCC development via targeting CNNM1
To elucidate whether SNHG7 regulates HCC progression via targeting CNNM1, rescue assays were conducted. First of all, CNNM1 mRNA and protein level was increased after the transfection of pcDNA3.1/CNNM1 into HCCLM3 and Hep3B cells, as reflected by RT-qPCR and Western blot assay, respectively (Supplementary Fig. S1A).
Subsequently, CCK-8 and colony formation assays revealed that the cell proliferation was inhibited by SNHG7 downregulation, but was overtly reversed by CNNM1 overexpression (Fig. 4A, B). More importantly, flow cytometry analysis elucidated that cell apoptosis was stimulated by SNHG7 knockdown, yet, hampered by CNNM1 overexpression (Fig. 4C). Identical with the findings of flow cytometry results, the expression alternation of apoptosis-associated proteins further validated that the counteracting impacts of CNNM1 overexpression on sh-SNHG7 promoted apoptotic ability (Fig. 4D).

SNHG7/miR-9-5p/CNNM1 axis enhances HCC development.
To further explore the tumor suppressing role of SNHG7 silence and the rescuing effect of CNNM1, the authors established xenograft mice models by injecting Hep3B cells. They found that the tumor growth speed was slowed down in sh-SNHG7 mice group, yet, fastened by overexpressing CNNM1 (Fig. 4E). Moreover, the tumor volume and weight were measured and found diminished in sh-SNHG7 mice group, yet, these inhibitory effects of sh-SNHG7 were abolished by overexpressing CNNM1 (Fig. 4F, G). Collectively, these in vivo data further elucidate that SNHG7 promotes HCC tumor growth via modulating CNNM1.
The expression patterns and survival analyses of SNHG7, miR-9-5p, and CNNM1 in tissue samples
To validate above data, the authors further explored the expression patterns of SNHG7 from bioinformatics channel and human tissue samples. They found a notable upregulation of SNHG7 in HCC tissues by browsing bioinformatics tool gene expression profiling interactive analysis, as data displayed in Figure 5A. Consistently, SNHG7 was detected highly expressed in HCC tissues in contrast to adjacent normal tissues (Fig. 5B). Kaplan–Meier survival analysis finding revealed that the high expression of SNHG7 was related to overall shorter survival among HCC patients (Fig. 5C).
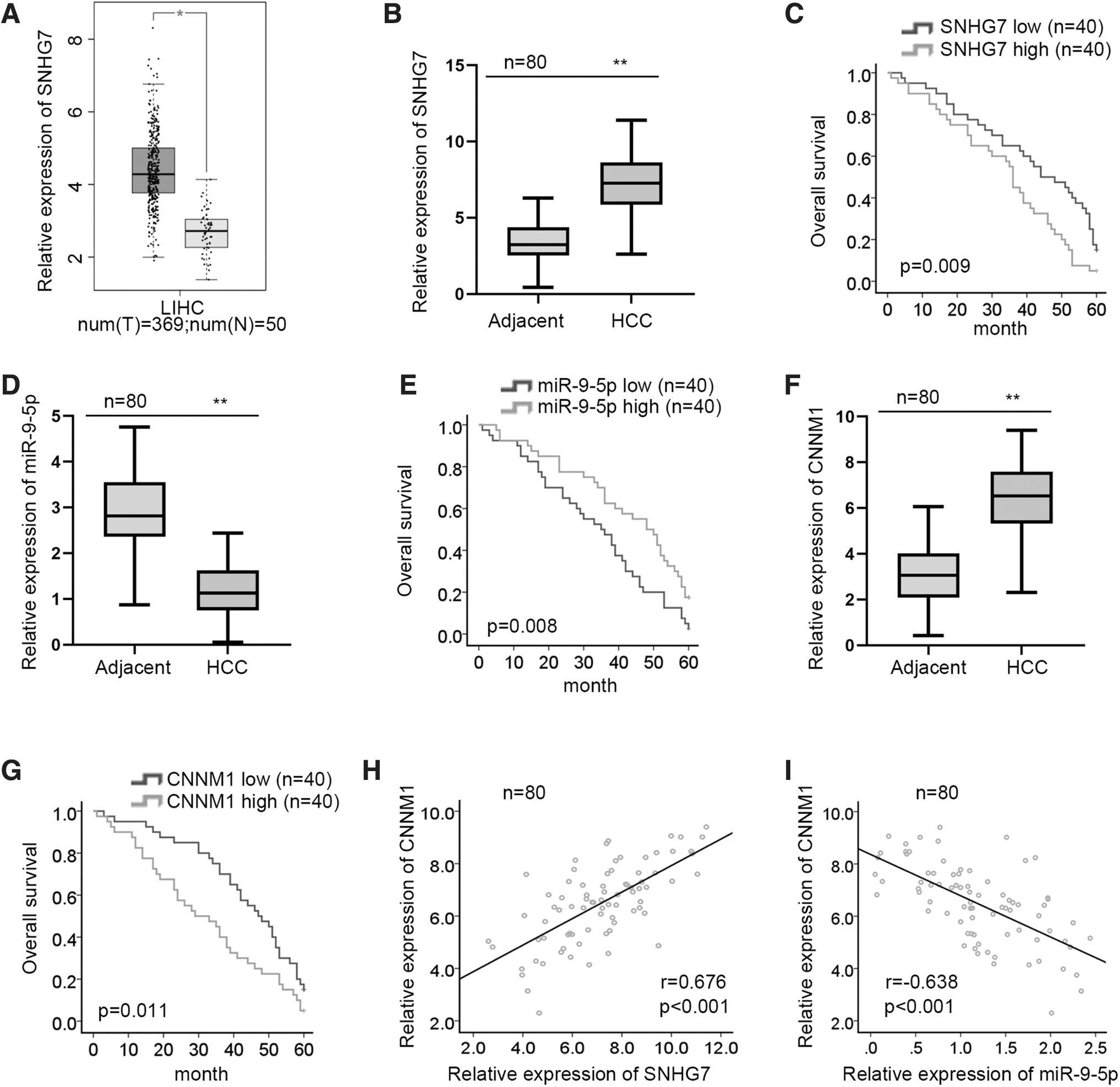
The expression patterns and survival analyzes of SNHG7, miR-9-5p, and CNNM1 in tissue samples.
Moreover, the authors observed that miR-9-5p was aberrantly depleted in HCC tissues compared with adjacent normal ones (Fig. 5D). They then studied the influence of miR-9-5p on HCC patients' overall survival by Kaplan–Meier survival analysis, which indicated that high expression of miR-9-5p had a relatively longer overall survival time (Fig. 5E). They then detected an abnormal upregulation of CNNM1 in HCC tissue specimens and an unfavorable effect of CNNM1 on overall survival rate (Fig. 5F, G). Thereafter, they examined the expression correlation between SNHG7/miR-9-5p and CNNM1 via Pearson correlation analysis. It was revealed that CNNM1 expression was positively correlated with SNHG7 expression, while negatively related to miR-9-5p expression in HCC samples, adding evidence for the SNHG7/miR-9-5p/CNNM1 ceRNA network (Fig. 5H, I).
Taken together, SNHG7 promotes HCC development via miR-9-5p/CNNM1 axis.
Discussion
Increasing novel biomarkers have been reported in combating HCC progression from molecular level. In the current work, the authors uncovered the critical role of SNHG7 in regulating HCC progression. Serving as a ceRNA, SNHG7 upregulates the mRNA level of CNNM1 via sequestering miR-9-5p in HCC cells.
lncRNAs play crucial roles in cell growth, differentiation, and epigenetic processes. 23,24 More importantly, lncRNAs are critical players in the malignant processes of HCC. 12,13 Previously, SNHG7 has been verified to enhance the development of colorectal cancer and nonsmall cell lung cancer. 18,19 SNHG7 inhibition could induce cell invasion inhibition in HCC. 20 In their study, it was validated that SNHG7 was overexpressed in HCC tissues and cell lines. SNHG7 silence weakened cell proliferation, while enhanced cell apoptosis in HCC. Judging by these data, SNHG7 is a carcinogenic gene in HCC.
ceRNA hypothesis is a classical posttranscriptional molecular regulatory mechanism of lncRNAs. In this novel regulatory mechanism, lncRNA could sponge miRNA to block miRNAs' function in repressing downstream target, thus affects cancer development. 25
Their study discovered that SNHG7 was mainly located in the cytoplasmic area of HCC cells, suggesting that SNHG7 may take part in the posttranscriptional regulation in HCC cells. 26 miR-9-5p has been elucidated as an anticancer gene in some cancers, such as papillary thyroid cancer. 27 In the research, the authors found a conspicuous downregulation of miR-9-5p in HCC cell lines. Furthermore, mechanical exploration validated that SNHG7 could bind with miR-9-5p. miRNAs mainly induce function via targeting and negatively modulating their downstream targets. The data suggested that SNHG7 may exert oncogenic functions through modulating miR-9-5p to affect its downstream target in HCC cells.
CNNM1 has been reported to be suppressed by ginsenoside Rh2 in restraining the malignant cell phenotype of prostate cancer. 28 Their research revealed that both mRNA and protein level of CNNM1 were upregulated by miR-9-5p overexpression in HCC cells. Moreover, CNNM1 was found to bind with miR-9-5p and exist in RISC. 29 A positive correlation between SNHG7 and CNNM1 was found in HCC tissue sample. Interestingly, rescue assays elucidated that the restrained HCC progression elicited by SNHG7 knockdown was largely offset by CNNM1 overexpression or miR-9-5p inhibition, implying that SNHG7 regulates HCC occurrence via targeting miR-9-5p/CNNM1 axis.
The authors' research experimentally confirmed the ceRNA regulatory mechanism of SNHG7 in HCC. Mechanically speaking, SNHG7 elevates CNNM1 expression via competitively interacting with shared miR-9-5p, thus facilitate CNNM1-induced cell growth in HCC. Their discovery may provide potent evidence for targeting SNHG7/miR-9-5p/CNNM1 axis in clinical treatment strategies of HCC, so as to improve the current unsatisfactory prognosis of HCC.
Footnotes
Acknowledgment
The authors appreciate the support of Eastern Hepatobiliary Surgery Hospital, Second Military Medical University.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.