
Abstract
Background:
Accumulating reports have demonstrated that long-noncoding RNAs (lncRNAs) play critical roles in the pathological progression of colorectal cancer (CRC). However, the role of lncRNA zinc finger E-box binding homeobox 2 antisense RNA 1 (ZEB2-AS1) in CRC remains largely unknown.
Methods:
The authors detected the ZEB2-AS1 expression in CRC tissue sample and CRC cell lines. The effects of ZEB2-AS1 on CRC were identified through in vitro assays (i.e., transwell assay, wound-healing assay, immunofluorescence assay, and Western blot) in a ZEB2-AS1 knockdown system. The molecular mechanism of ZEB2-AS1 was explored via bioinformatic tools, quantitative real-time polymerase chain reaction (qRT-PCR), dual-luciferase reporter assay, RNA immunoprecipitation assay, and so on. Moreover, a series of gain-of-function experiments were performed to identify the effect of ZEB2-AS1 and miR-1205 on epithelial-to-mesenchymal transition (EMT) in CRC cells.
Results:
This analysis clarified that ZEB2-AS1 was upregulated in both CRC tissue sample and cells lines; meanwhile, the high expression of ZEB2-AS1 was correlated with poor overall survival rate. ZEB2-AS1 knockdown significantly suppresses the EMT in CRC cells. Furthermore, the authors identified that the expression of ZEB2-AS1 was negatively correlated with expression of miR-1205, and CRKL could be a direct target of miR-1205. Through the gain-of-function experiments, they found that ZEB2-AS1 accelerates EMT in CRC cells via modulating the expression of miR-1205 and CRKL.
Conclusion:
Taken together, this study revealed that ZEB2-AS1 accelerates EMT in CRC through the miR-1205/CRKL pathway, suggesting that ZEB2-AS1 may potentially serve as a target of CRC.
Introduction
Colorectal cancer (CRC) is now ranked the third-most wide-ranging cancer and is increasingly becoming the leading cause of morbidity and mortality globally. 1 The most widely used treatment strategies are surgical resection followed by adjuvant chemotherapy and radiation therapy. 2 –4 During the last few decades, despite great advances have been achieved regarding the early diagnosis and treatment of CRC, the long-term survival rate of patients still remains at a low level. 5,6 The most common reason of low survival rate is the frequency of recurrence and distant metastasis in advanced stage of CRC. 7 Unfortunately, until now, the underlying mechanisms of malignant growth and metastatic behaviors of CRC cells are not fully understood.
Cancer metastasis consists of many steps, including sequential, interlinked, and selective processes. 8 Among those steps, the cross talk between two cellular states (epithelial and mesenchymal phenotypes), the so-called epithelial-to-mesenchymal transition (EMT), has been confirmed to functionally promote CRC metastasis. 9,10 The process of EMT is quite complex and includes multiple biochemical changes that could disrupt cell–cell adherence, remodel the cytoskeleton, as well as enhance migratory capacity and cell invasiveness. 11 In CRC, the main transcriptional regulators involved in the EMT process include Snail zinc-finger family, basic helix–loop–helix family, and zinc-finger E-box-binding homeobox family. 12 –15
Recently, numerous studies have highlighted that noncoding RNAs (ncRNAs) such as microRNAs (miRNAs) and long-noncoding RNAs (lncRNA) play regulatory roles in the pathogenesis of CRC. LncRNAs are usually defined as transcripts longer than 200 nucleotides, which involved in transcriptional and translational regulation in a wide range of cellular processes, such as gene transcription regulation and X chromosome inactivation. 16,17 Different from lncRNAs, the length of miRNAs is much shorter (22–25 nucleotides). In addition, miRNAs basically negatively regulate target genes by binding to 3′-untranslated regions (3′-UTR) of their target mRNAs. 18 So far, numerous ncRNAs have been proven to play essential roles in multiple cellular and molecular processes, including migration, invasion, apoptosis, and angiogenesis in the progression of many cancers. 19,20
In CRC, EMT progression can be regulated by many lncRNAs, such as CPS1 intronic transcript 1 (CPS1-IT1), 21 cardiac hypertrophy-related factor, 22 and SLC25A25-AS1. 23 As a newly found tumor-associated lncRNA, zinc finger E-box binding homeobox 2 antisense RNA 1 (ZEB2-AS1) was demonstrated to be highly expressed in many cancer cells. 24 –26 However, the functional role of ZEB2-AS1 in CRC is yet to be fully understood.
In the present study, the authors aimed to clarify the biologic function of lncRNA ZEB2-AS1 in CRC. Furthermore, they also would like to investigate the related miRNA and its pathway involved in CRC pathogenesis. These findings could set a novel insight into the pathologic mechanism of CRC and might provide new therapeutic strategies for this disease.
Materials and Methods
Tumor sample collection
Patients (n = 50, 45–70 years old) with CRC who underwent colorectal resection at the First People's Hospital of Wenling were enrolled in this study. Fifty CRC tissue sample and paired adjacent tissue samples were collected and preserved from April 2016 to September 2018. All samples were snap-frozen in liquid nitrogen immediately after surgery (−80°C) for further experiments. Before surgery, special therapies (chemotherapy or radiotherapy) were not applied to any one of the patients. In addition, the Ethics Committee of the First People's Hospital of Wenling reviewed and approved all protocols of this study. Meanwhile, all participants provided written informed consent.
Cell line selection and culture
The cell lines used in this study were as follows: normal human colon mucosal epithelial cell line (NCM460), human CRC cell lines (T84, SW620, SW480, HT-29), and human embryonic kidney 293 cells (HEK293). All cells were purchased from the Chinese Academy of Sciences (Shanghai, China) and cultured at a constant temperature in a humidified cell incubator with 95% air and 5% CO2 at 37°C in DMEM (Invitrogen, Carlsbad, CA) supplemented with 10% FBS (Gibco, Grand Island, NY) and 1% streptomycin/penicillin (100 μg/mL and 100 μg/mL) (Invitrogen).
Cell transfection
Short hairpin RNA (shRNA) targeting ZEB2-AS1 (sh1#-ZEB2-AS1, sh2#-ZEB2-AS) and negative control (scramble), plasmid pcDNA3.1-ZEB2-AS1 for overexpression of ZEB2-AS1 and control vector, miR-1205 mimics for overexpression and mimics control (NC mimics) were obtained from GeneChem (Shanghai, China). Cells were transfected using Lipofectamine 2000 (Invitrogen).
Quantitative real-time polymerase chain reaction
Total RNA from different groups was extracted by the TRIzol reagents (Invitrogen). Then, cDNA and reverse-transcribed RNA were obtained. cDNA's quality and quantity were redetermined via detecting the absorbance at 260 nm/280 nm (A260/A280). The primers used in these experiments were purchased from Sangon Biotech (Shanghai, China) and are listed in Table 1. Then, qRC-PCR was performed using the SYBR Green Real-Time PCR Master Mix (Toyobo, Japan) and performed on ABI 7500 Real-Time PCR Systems. The internal controls for miR-1205 and CRKL mRNA were U6 and GAPDH, respectively. The relative abundance of the target genes was calculated based on the formula: RQ = 2−ΔΔCt.
Primers Used in the Present Study
Wound-healing assay
Different groups of cells (5 × 105) were suspended in 0.6 mL cell growth medium and seeded into a six-well culture plate. The cell platform was scratched using a 10-μL pipette tip and then washed at least three times with PBS. Subsequently, cell movement was recorded under an Olympus IX70 inverted microscope.
Transwell assay
Transwell assay was conducted in this study to clarify the migration and invasion abilities of CRC cells. Briefly, a total number of 5 × 103 cells cultured in upper compartments were in an environment with DMEM (without FBS). Differently, the lower compartments were filled with DMEM (with 20% FBS). After 24 h, the invaded cells were fixed with 4% paraformaldehyde (PFA) and stained with 0.5% crystal violet. The number of migrated and invasive cells was counted under an Olympus microscope (40 × 10).
Immunofluorescence assay
Immunofluorescence staining was performed on cells plated on chamber slides (Nalge Nunc) based on a previous protocol. 27 Subsequently, cells were treated with 100 ng/mL CTGF for 96 h and then were fixed with 4% (PFA) for 5 min. After being incubated with primary antibodies, cells were probed with mouse antibodies against human E-cadherin and N-cadherin. Then, a secondary antibody, Texas Red-conjugated anti-mouse IgG (Vector Labs), was added. The cells were counterstained with 4′,6′-diamidino-2-phenylindole (DAPI; ab228549, Abcam, Cambridge, United Kingdom). All the above procedures were carried out in a dark room. The samples were observed and imaged under a fluorescence microscope (Leica DMI6000, Solms, Germany).
Western blot assay
Total proteins from different groups of SW620 and SW480 cells were extracted using the RIPA lysis buffer. Protein samples were loaded onto 10% or 12% sodium dodecyl sulfate/polyacrylamide gel electrophoresis gels depending on molecular weight and transferred onto 0.45 mm polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA). The PVDF membranes were blocked in 5% nonfat skim milk/TBST. The primary antibodies were as following: rabbit anti-ZEB1 (1:1000; ab203829; Abcam); rabbit anti-Snail (1:1200; ab53519; Abcam); anti-MMP2 (1:1000; ab37150; Abcam); anti-CRKL (1:1000; ab151791; Abcam); and rabbit anti-GAPDH (1:1200; ab8245; Abcam). Subsequently, goat anti-rabbit secondary detection antibodies (1:4000; ab205718; Abcam) were added. The protein of interest was detected using the ChemiDoc XRS detection system (Bio-Rad, Milan, Italy).
Dual-luciferase reporter assay
The 3′-UTR of ZEB2-AS1 was cloned into the pmirGLO vector (Promega, Madison, WI) and named wild-type (WT) ZEB2-AS1–3′-UTR. In addition, CRKL 3′UTR containing the predicted miR-1205 binding site or the corresponding mutant binding sites were also cloned into the pmirGLO vector (Promega). Subsequently, ZEB2-AS1 WT or ZEB2-AS1 MUT, as well as CRKL 3′UTR WT or CRKL 3′UTR MUT, was cotransfected with miR-1205 mimics or NC mimics in HEK 293 cells using Lipofectamine 3000 (Invitrogen, Waltham, MA). The dual-luciferase reporter assay was conducted using a Dual-Luciferase Reporter Assay System (Promega).
RNA immunoprecipitation assay
To identify the relationship between endogenous ZEB2-AS1 and miR-1205, the RNA immunoprecipitation (RIP) assay was conducted. Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA) was used in this study. Based on the manufacturer's instructions, different groups of cell lysates were with RIP buffer containing magnetic beads conjugated with the human anti-Ago2 antibody or negative control IgG. Then, the precipitation of RNA was isolated and analyzed by qRT-PCR.
Statistical analysis
All cell experiments were repeated independently at least three times. All data were exhibited as mean ± SD. The SPSS (17.0, Inc., Chicago, IL) was used for statistical analysis. Statistical analysis was performed using two-tailed Student's t-test or one-way ANOVA. In addition, survival curves and correlation analysis were evaluated by Kaplan–Meier analysis and Pearson correlation coefficient, respectively. *p < 0.05, **p < 0.01, or ***p < 0.001 was considered to indicate statistical significance.
Results
The expression of ZEB2-AS1 was upregulated in CRC tissues and cells
In the very beginning, the authors detected the ZEB2-AS1 expression in 50 human CRC tissues and adjacent normal tissues with qRT-PCR. The expression of ZEB2-AS1 was markedly upregulated compared with normal tissues (p < 0.01, Fig. 1A). Furthermore, a Kaplan–Meier survival analysis showed that high expression of ZEB2-AS1 correlated with a lower overall survival rate (p < 0.01, Fig. 1B).

The expression of ZEB2-AS1 in colorectal cancer tissues and cells.
In addition, the expression of ZEB2-AS1 was detected in both normal human colon mucosal epithelial cell line (NCM460) and CRC cell lines (T84, SW620, SW480, HT-29) with qRT-PCR. The results demonstrated that ZEB2-AS1 was upregulated in all CRC cell lines compared with NCM460 cell line, especially in SW620 and SW480 cell lines (p < 0.01, Fig. 1C). Therefore, the authors used SW620 and SW480 cell lines for functional studies. The above results suggested that LncRNA ZEB2-AS1 was upregulated in both CRC tissues and cell lines and negatively correlated with the survival rate of CRC patients.
Knockdown of ZEB2-AS1 attenuated EMT process in CRC cells
To identify the role of ZEB2-AS1 in the pathogenesis of CRC, the authors transfected two shRNAs targeting ZEB2-AS1 (sh1#-ZEB2-AS1, sh2#-ZEB2-AS) to test the knockdown efficiency. As shown in Figure 2A, sh2#-ZEB2-AS had better knockdown efficiency and thus was chosen for further studies (p < 0.01). Subsequently, a series of loss-of-function experiments were performed. Wound-healing assay and transwell assay were used for determining the migration and invasion ability of CRC cells.
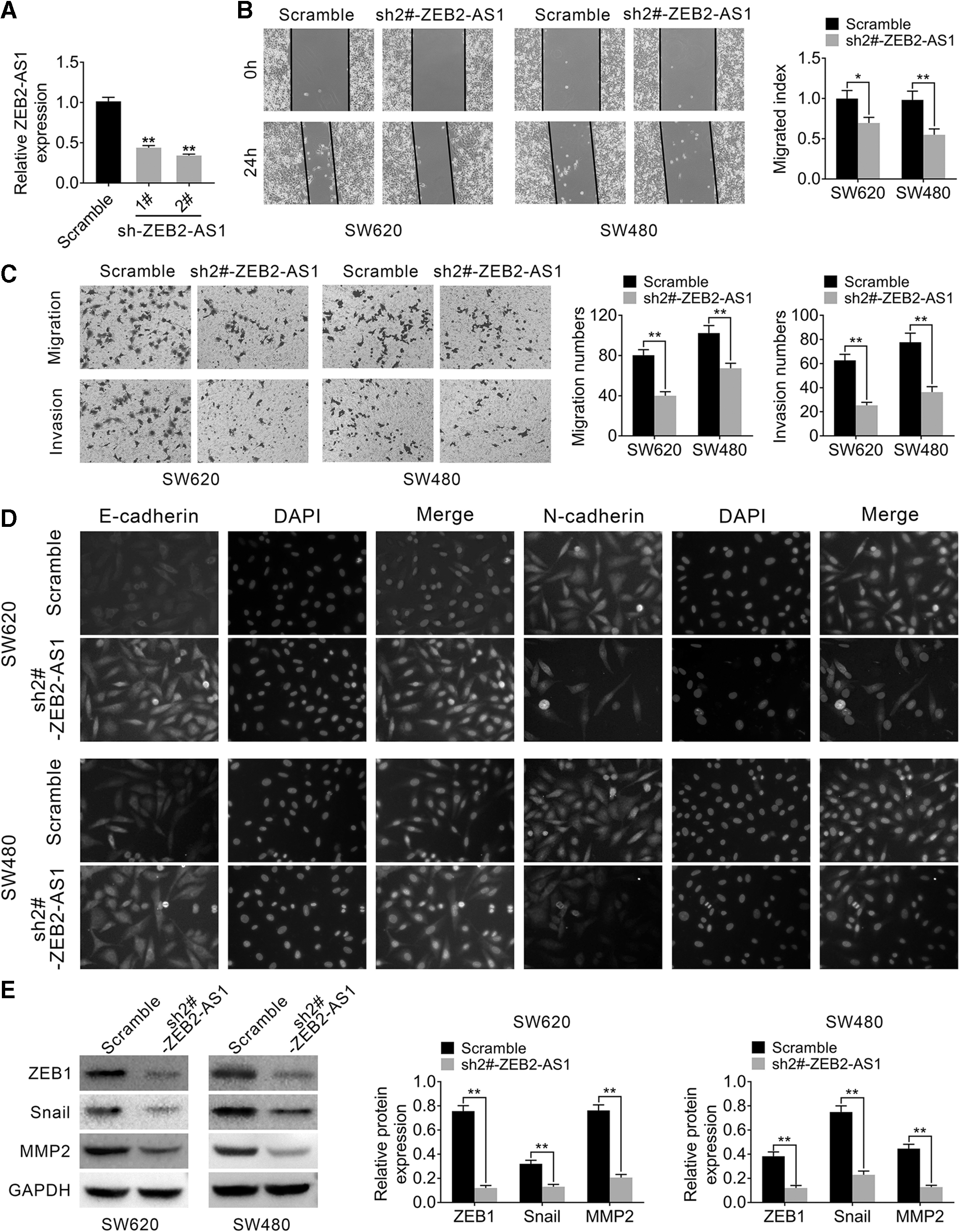
Knockdown of ZEB2-AS1 suppressed EMT in CRC cells.
As shown in Figure 2B and C, the migration and invasion abilities of CRC cells were significantly suppressed by knockdown of ZEB2-AS1 compared with the scramble group (p < 0.05, p < 0.01). Moreover, to understand the underlying mechanism, immunofluorescence assay and Western blot were performed. The authors found that epithelial markers of EMT (such as E-cadherin), which could attenuate the EMT process, were upregulated in the ZEB2-AS1-knockdown group. However, the EMT, which could promote the EMT processes such as N-cadherin, ZEB1, Snail, and MMP2, was downregulated in the ZEB2-AS1-knockdown group at the protein level, suggesting that knockdown of ZEB2-AS1 suppressed the EMT in CRC cells (p < 0.01, Fig. 2D, E).
The expression of ZEB2-AS1 was negatively correlated with expression of miR-1205
To further clarify the molecular mechanisms underlying the effects of ZEB2-AS1 on CRC cellular behavior, the bioinformatic analysis (miRDB and TargetScan) was used to explore the potential target of ZEB2-AS1. Interestingly, the authors found the putative miR-1205 response elements on ZEB2-AS1 (Fig. 3A). First, the overexpression efficiency of miR-1205 was detected by qRT-PCR. As shown in Figure 3B, in both SW620 and SW480 cell lines, the expression of miR-1205 was significantly upregulated in the miR-1205 mimic group compared with miR-NC control, indicating that the present condition was suitable for further experiments (p < 0.01). Furthermore, the dual-luciferase reporter assay was conducted to determine the relationship between ZEB2-AS1 and miR-1205. The results showed that the luciferase activity of cells, which transfected with the ZEB2-AS1 WT plasmid, was markedly suppressed by overexpression of miR-1205 (p < 0.01), however, the ZEB2-AS1 MUT plasmid group showed no difference (p > 0.05, Fig. 3C), suggesting that miR-1205 was able to bind the ZEB2-AS1 transcript. To validate the interaction between miR-1205 and ZEB2-AS1, RIP assay was conducted.
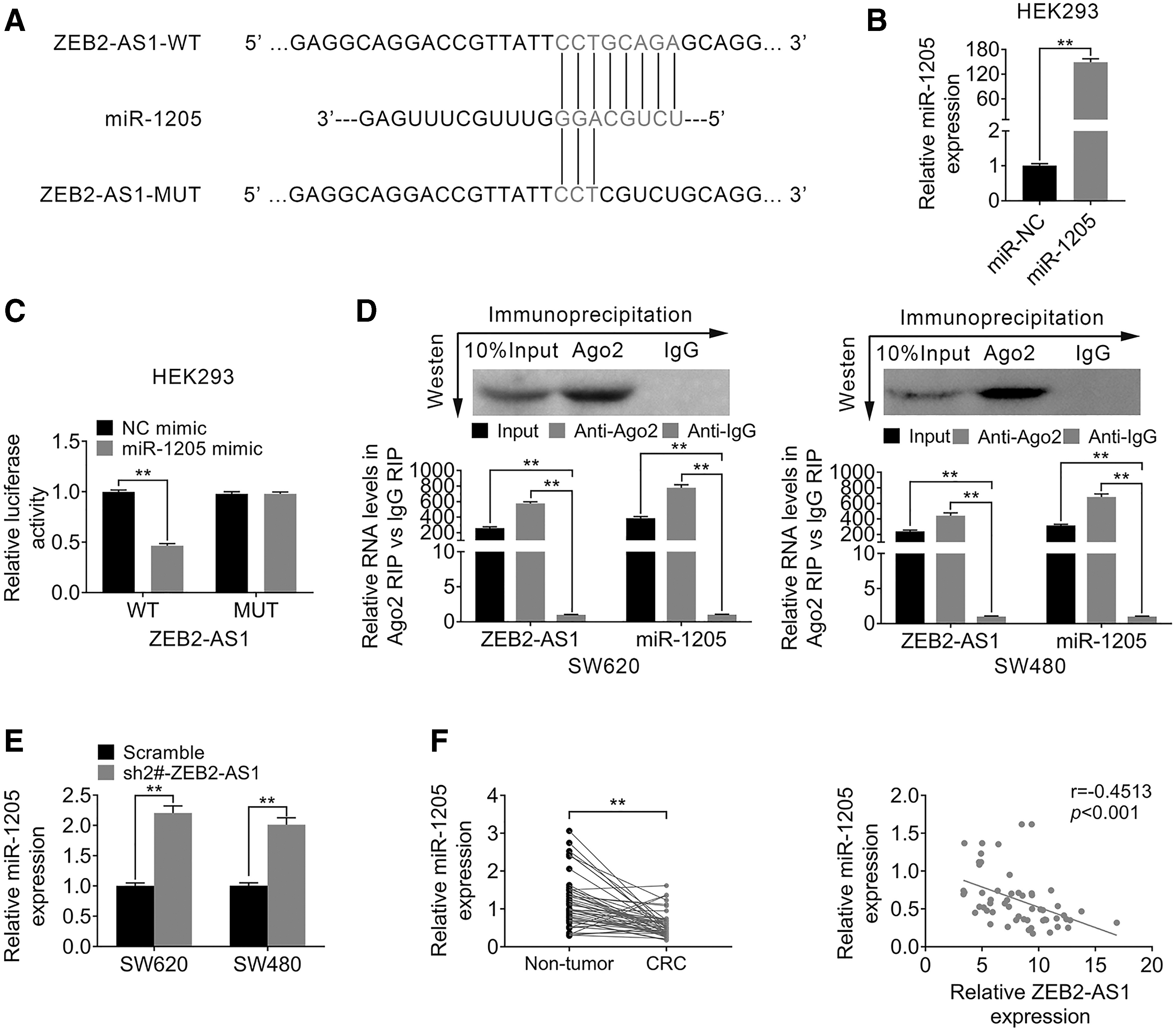
The expression of ZEB2-AS1 was negatively correlated with expression of miR-1205.
As shown in Figure 3D, in both SW620 and SW480 cell lines, the endogenous miR-1205 expression was specifically enriched in the cells treated with the ZEB2-AS1 overexpression plasmid (p < 0.01, Fig. 3D). Furthermore, knockdown of ZEB2-AS1 could markedly promote the expression of miR-1205 (p < 0.01, Fig. 3E). Moreover, the expression of miR-1205 was detected in human CRC and normal tissues. The data demonstrated that the expression level of miR-1205 was significantly less in CRC tissues, which is opposite with the expression of ZEB2-AS1; the correlation analysis further showed that the level of miR-1205 was negatively correlated with expression of ZEB2-AS1 (r = 0.4513, p < 0.001, Fig. 3F). Taken together, these data suggested that the expression of ZEB2-AS1 was negatively correlated with expression of miR-1205; moreover, ZEB2-AS1 could be a target of miR-1205.
CRKL was a target of miR-1205
To further understand the role of miR-1205 and its pathway in the pathogenesis of CRC, the authors used the bioinformatic tool (miRDB and TargetScan) and found that V-Crk avian sarcoma virus CT10 oncogene homolog-like (CRKL) was a potential target of miR-1205 (Fig. 4A). They further confirmed this hypothesis by luciferase reporter assays (p < 0.01, Fig. 4B). Subsequently, the relative protein level of CRKL was identified in SW620 and SW480 cells, which were transfected with miR-1205 mimics or NC mimics. These results demonstrated that after transfecting with miR-1205 mimics, protein levels of CRKL were significantly downregulated (p < 0.01, Fig. 4D).

CRKL was a direct target of miR-1205.
Moreover, the expression of mRNA CRKL was measured in human CRC tissues and normal tissues (n = 50). The data revealed that the mRNA level of CRKL was significantly upregulated in human CRC tissues compared with control (p < 0.01, Fig. 4E). In addition, the correlation between CRKL and ZEB2-AS1, and CRKL and miR-1205, was analyzed, suggesting that the expression of CRKL positively correlated with ZEB2-AS1 level (r = 0.5721, p < 0.001), while negatively correlated with miR-1205 level (r = 0.4911, p < 0.001, Fig. 4F). Overall, these results revealed that CRKL was considered to be a target of miR-1205.
ZEB2-AS1 accelerates EMT in CRC cells via modulating miR-1205/CRKL pathway
Given that ZEB2-AS1 directly interacted with miR-1205, and meanwhile, CRKL was a direct target of miR-1205, the authors hypothesized that ZEB2-AS1 might exert its role in regulating EMT in CRC cells via modulating the miR-1205/CRKL pathway. To this end, they performed a series of gain-of -function experiments. First, they transfected ZEB2-AS1 expression plasmid pcDNA3.1- ZEB2-AS1 into SW620 cells and acquired ZEB2-AS1 stably overexpressed in SW620 cells (p < 0.01, Fig. 5A). Furthermore, cell migration and invasion were determined by functional experiments in SW620 cells, which were cotransfected with pcDNA3.1- ZEB2-AS1 plasmid or vector, and miR-1205 mimic or NC mimic.

ZEB2-AS1 accelerates EMT in SW620 cells via modulating the miR-1205/CRKL pathway.
As shown in Figure 5B and C, the overexpression of ZEB2-AS1 promoted cell migration and invasion (p < 0.05, p < 0.01). However, this effect was reversed by miR-1205 (p < 0.05, p < 0.01). Moreover, immunofluorescence assay and Western blot were conducted to determine the EMT markers such as E-cadherin, N-cadherin, ZEB1, Snail, and MMP2 in transfected SW620 cells. The results demonstrated that overexpression of ZEB2-AS1 could accelerate the EMT process by downregulating E-cadherin, while upregulating N-cadherin, ZEB1, Snail, and MMP2 protein. However, this effect was attenuated by overexpression of miR-1205 (p < 0.01, Fig. 5D, E), indicating that ZEB2-AS1 accelerates EMT in SW620 cells via modulating the miR-1205/CRKL pathway.
Discussion
Despite great progress defining molecular and cellular mechanisms of the EMT process in CRC, there is still no effective clinical treatment for CRC patients, especially of those in advanced stage with distant metastasis. 28 Most are the failures of control of EMT program. 29,30 As a complex process, EMT can be regulated by many transcriptional regulators and noncoding RNAs in tumor budding, circulating, and drug resistance. 31,32 For instance, Bai et al. showed that lncRNA SNHG1 could promote the EMT course intrinsically by cooperating with miR-497 and miR-195-5p in CRC. 33
Many other studies also identified that lncRNAs serve as competing endogenous RNAs (ceRNAs), or in other words “sponge,” in the regulatory network referring to miRNA, lncRNA, and their target genes. One example is that in Liang et al's study, lncRNA H19 acts as a ceRNA of miR-138 and miR-200a, suppressing their functions to promote the EMT process in CRC. 34 In addition, HIF1A-AS2 positively affected the progression and EMT formation in CRC by regulating the miR-129-5p/DNMT3A axis. 35
In the present study, the authors focused on a new lncRNA called ZEB2-AS1, which is a highly conserved natural antisense transcript that corresponds to the 5′UTR of ZEB2. 36 ZEB2-AS1 has been found to be highly expressed in many different types of cancer tissues and cells; meanwhile, it also proved to be an oncogenic lncRNA in human cancer by accelerating cell proliferation, invasion, and MET process. 24 –26 Not surprisingly, this study found that ZEB2-AS1 was also highly expressed in CRC tissues. Moreover, by conducting a series of loss-of-function experiments, they identified that knockdown of ZEB2-AS1 attenuates the EMT process in CRC cell line, suggesting that ZEB2-AS1 indeed plays a regulatory role in the CRC pathologic process.
The authors further would like to identify the potential molecular mechanisms of ZEB2-AS1 in CRC cell lines. By using bioinformatic tools, they interestingly found that miR-1205 could be a potential target miRNA of ZEB2-AS, which was further confirmed by luciferase reporter assays. Moreover, CRKL was a direct target of miR-1205. In addition, mRNA CRKL expression was positively correlated with ZEB2-AS1 level, while it negatively correlated with miR-1205 expression, suggesting that there could be a potential regulatory effect among those factors.
As an important downstream factor in this study, CRKL also confirmed being involved in CRC pathological progressions in many published studies. For instance, Franke et al. reported that the expression of CRKL is markedly elevated in human CRC tissue. Moreover, CRKL could act as a signal adaptor to interact with SASH1 and further regulate EMT and metastasis in CRC. 37 By inhibiting the expression of CRKL in colon cancer cells, the proliferation, migration, and invasion abilities of cells were suppressed, indicating that CRKL acts as a tumor promoter in CRC. 38
In the last part of this study, to identify the regulatory effect of ZEB2-AS1 on miR-1205 and CRKL, the authors performed a series of gain-of-function experiments. The results demonstrated that ZEB2-AS1 could promote the EMT process, and this effect can be attenuated by overexpression of miR-1205, suggesting that ZEB2-AS1 accelerates EMT in CRC cells via modulating the miR-1205/CRKL pathway. However, there are some limitations in the present study; for instance, they did not introduce CRKL knockdown or overexpression system into the functional experiments. In addition, from this study, they only confirmed that the high expression of ZEB2-AS1 correlated with a lower survival rate of CRC patients. However, the correlation between ZEB2-AS1 expression and other clinical features remains unknown.
To sum up, this study provides the evidence of LncRNA ZEB2-AS1 as a regulator of EMT process in CRC. ZEB2-AS1 could accelerate EMT via modulating the miR-1205/CRKL pathway in CRC. These findings contribute to a better understanding of molecular mechanism of EMT process in CRC progression and might provide a new therapeutic target for CRC treatment.
Footnotes
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Ethics Approval and Consent to Participate
The animal use protocol listed below has been reviewed and approved by the Animal Ethics and Welfare Committee.
Authorship Confirmation Statement
Y.H.J. and X.L. conceived and designed the experiments; G.M.L. and W.Y. analyzed and interpreted the results of the experiments; and J.J.X., C.F.S., and X.W.W. performed the experiments.
Informed Consent
Written informed consent was obtained from a legally authorized representative(s) for anonymized patient information to be published in this article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.