
Abstract
Background:
Cervical cancer (CC) ranks fourth in cancers that resulted in death among women, accumulating the attention of researchers. It has been ascertained that long noncoding RNAs (lncRNAs) are crucial players in the pathological processes of a host of cancers. And, SNHG7 has been reported to enhance the occurrence of various cancers; however, its function in CC sustains obscure.
Aim of the Study:
This study explored the function of SNHG7 in CC and further investigates the specific molecular mechanism of SNHG7 in regulating CC.
Methods:
The levels of SNHG7 in CC cells were reflected by quantitative real-time polymerase chain reaction. The functions of SNHG7 on CC tumorigenesis were explored by colony formation, CCK-8 (Cell Counting Kit-8), EdU (ethynyl deoxyuridine), and Western blot assays. The influences of SNHG7 depletion on the binding of EZH2 to DKK1 promoter and H3K27me3 occupancy in DKK1 promoter were studied by chromatin immunoprecipitation assay.
Results:
SNHG7 was conspicuously higher expressed in CC cells. Knockdown of SNHG7 was detected to ameliorate the malignant behaviors of CC cells. Importantly, the contribution of SNHG7 to CC development was relied on activated Wnt pathway through DDK1-mediated manner. Furthermore, it was confirmed that SNHG7 silencing weakened the binding of EZH2 to DKK1 promoter as well as the occupancy of H3K27me3 in DKK1 promoter.
Conclusions:
SNHG7 epigenetically silences DKK1 to exacerbate the malignancy of CC via Wnt/β-catenin signaling pathway.
Introduction
Cervical cancer (CC) ranks as the fourth most dreadful cancer among women, with ∼300,000 deaths all over the world per year. 1 Extensive work has discovered that around 99.8% CC sufferers are featured with human papillomavirus (HPV) infection, 2 of which HPV-16 and HPV-18 account for nearly two thirds of CC cases. 3 Oncogenes of high-risk HPV such as E6 and E7 have been reported to serve as pivotal players in the malignancy of CC. 4 Nevertheless, previous research has illustrated that HPV infection itself is not sufficient to result in CC; oncogenic genes and anticancer genes are also implicated in the tumorigenesis or cancers. 5 Therefore, the exploration of the biological molecules involved in CC occurrence will be beneficial to the improvement of prognosis.
Long noncoding RNAs (lncRNAs) have been recently identified as a type of noncoding RNA, characterized by its length of more than 200 nucleotides. 6 Although lncRNAs were considered to possess no biological function, 7 extensive work has demonstrated that lncRNAs can modulate gene expression via the transcriptional and post-transcriptional regulation. 8 In the past decades, it has been validated that lncRNAs act as tumor activators or suppressors in a host of cancers, including CC, suggesting that lncRNAs can be utilized as indicators in the diagnosis of cancers at early stage. 9 –11 Existing studies pointed out that lncRNA small nucleolar RNA host gene 7 (SNHG7) serves as a malignancy promoter in several cancers 12,13 ; however, the biological role of SNHG7 in CC waits to be uncovered.
Wnt/β-catenin signaling pathway has been elucidated to expedite the malignant neoplasia in cancers, including CC. 14 –16 Previously, SNHG7 has been revealed to activate Wnt/β-catenin pathway in bladder cancer. 17 Therefore, Western blot and luciferase assays were employed to explore the regulation of SNHG7 on Wnt/β-catenin signaling pathway in CC. Moreover, DKK1 has been verified to deactivate Wnt/β-catenin signaling pathway. 18 And at the transcription level, lncRNAs could bind with enhancer of zeste homolog 2 (EZH2) to epigenetically suppress the expression of their downstream messenger RNAs (mRNAs). 19,20 Hence, the authors hypothesized that SNHG7 might epigenetically suppress DKK1 to activate Wnt/β-catenin signaling pathway.
This study was carried out to quest for the influences of SNHG7 on the cellular process of CC. Besides, the regulation of SNHG7 on Wnt/β-catenin signaling pathway would be uncovered. More importantly, the mechanism through which SNHG7 regulates DKK1 would be clarified by RNA immunoprecipitation (RIP), chromatin immunoprecipitation (ChIP), and Western blot assays.
Materials and Methods
Cell culture and treatment
Human immortal cervical epithelial H8 cells and four internationally accepted CC cell lines (C-33A, CaSki, SiHa, and HeLa) were bought from American Type Culture Collection (ATCC, Manassas, VA) maintaining in RPMI 1640 medium. Both 10% fetal bovine serum (FBS; Hyclone, Camarillo, CA) and 1% penicillin–streptomycin (Hyclone) were used to supplement medium at 37°C in 5% CO2. LiCl (Invitrogen, Camarillo, CA), the Wnt signaling pathway activator, was added into the culture medium after transfection.
Quantitative real-time polymerase chain reaction
According strictly to the established protocol of TRIzol Reagent (Invitrogen), cellular RNA was extracted from CaSki and C-33A cells for reverse transcription. Using PrimeScript™ II 1st Strand cDNA Synthesis Kit from Takara (Tokyo, Japan), the first-stand cDNA was acquired. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed by use of SYBR® Premix Ex Taq™ II Kit (Takara) on 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA), with the GAPDH as an endogenous control for the quantification. Gene expressions were calculated following the comparative Ct method. All primer sequences utilized are listed in Table 1.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Lentiviral infection and cell transfection
CaSki and C-33A cells were plated into six-well plates with the cell density of 2 × 105 cells per well. For short hairpin RNA (shRNA) interfering transfection, the shRNA eukaryotic expression vector was designed by GenePharma (Shanghai, China) to target SNHG7, EZH2, and DKK1 genes, as well as control shRNA sequences. The pcDNA3.1/SNHG7 and pcDNA3.1 expression vectors were designed in similar way. The recombinant lentiviral particles containing above plasmids were obtained in line with the previous protocol. 21 Thereafter, cells were processed with the lentiviral particles for 72 h, with transfection efficiency assessed by qPCR. Finally, the stably transfected cells were chosen under puromycin selection.
Cell Counting Kit-8 assay
One hundred microliters of cell suspension was preseeded into 96-well plates. 0, 12, 24, 48, or 96 h after cell transfection, 10 μL of Cell Counting Kit-8 (CCK-8) reagent (Solarbio, Beijing, China) was added for 2 h of incubation in the dark, followed by the absorbance measuring at 450 nm.
Colony formation assay
CaSki and C-33A cells were put in six-well plates (500 cells per well) and cultivated for 14 d. Cells were then dyed by 0.5% crystal violet in methanol for 15 min, following the determination of clone formation rate.
Ethynyl deoxyuridine staining assay
Ethynyl deoxyuridine (EdU) kit was initially purchased from RiboBio (Guangzhou, China) for EdU staining in light of the user manual. CaSki and C-33A cells were transfected and fixed in 4% paraformaldehyde. After nuclei staining with DAPI (RiboBio) in the dark, the percentage of EdU-positive cells was assessed by Zeiss Photomicroscope (Carl Zeiss, Oberkochen, Germany).
Western blot
Total cellular lysates were isolated from transfected CaSki and C-33A cells in RIPA lysis buffer (Beyotime, Shanghai, China) containing protease inhibitors (Beyotime). Following separation by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, protein samples were transferred to polyvinylidene difluoride membrane (PVDF; Millipore, Billerica, MA), which was then treated with 5% nonfat milk for 2 h at room temperature. Primary antibodies against Bax (ab81083), Bcl-2 (ab201566), cleaved caspase-3 (ab49822), total caspase-3 (ab13847), DKK1 (ab93017), β-catenin (ab22656), c-myc (ab39688), cyclin D1 (ab40754), EZH2 (ab191080), and GAPDH (ab37168) as loading control, and horseradish peroxidase (HRP)-conjugated secondary antibody were both obtained from Abcam (Cambridge, MA). The target proteins were visualized with an enhanced chemiluminescence system (ECL; Beyotime).
Luciferase reporter assay
TOP/FOP-Flash luciferase reporter assay was conducted for the Wnt/β-catenin pathway activity. TOP-Flash/FOP-Flash and pRL-TK reporter from Promega (Madison, WI) were separately co-transfected with SNHG7 silence vector (sh-SNHG7#1 or sh-NC), (pcDNA3.1/SNHG7 or pcDNA3.1), and (sh-DKK1#1 or sh-NC), into CaSki and C-33A cells. For DKK1 promoter analysis, cells were planted in 24-well plates and co-transfected with luciferase reporter plasmid containing DKK1 promoter, pRL-TK-Renilla plasmid, and SNHG7 silence vector. Dual-Luciferase Reporter Assay System (Promega) was applied at 48 h after transfection.
Nuclear–cytoplasmic fractionation assay
Cytoplasmic and Nuclear RNA Purification Kit from Norgen (Belmont, CA) was purchased to isolate cytoplasmic and nuclear RNA from CaSki and C-33A cells as requested by supplier. Expression level of SNHG7 (specific gene), GAPDH (cytoplasm control), and U6 (nucleus control) was assayed by qRT-PCR.
RIP assay
EZ Magna RIP Kit was first bought from Millipore for RIP assay. CaSki and C-33A cells were cultured in complete RIP lysis buffer, and the lysates were treated with RIP buffer, which contained magnetic beads with anti-EZH2 or anti-IgG antibodies (Millipore) at 4°C for 4 h. The relative RNA enrichment was determined by qRT-PCR in precipitated complex.
ChIP assay
CaSki and C-33A cells were subjected to formaldehyde for 10 min for obtaining DNA–protein cross-links. Then, cell lysates were sonicated to chromatin fragments of 200–500 base pairs and immunoprecipitated with anti-EZH2, anti-H3K27me3, or anti-IgG antibodies (Millipore). At length, the retrieved chromatin from precipitated complex was analyzed by qRT-PCR.
Fluorescence in situ hybridization
After fixing, the rinsed CC cells were collected and incubated with the specific fluorescence in situ hybridization (FISH) probe for SNHG7 (Ribobio) in the hybridization buffer. Olympus microscope was used to observe cells after nuclear counterstaining with DAPI.
RNA pull-down assay
Biotinylated SNHG7 and anti-sense SNHG7 RNAs were mixed with the cell lysates of CaSki and C-33A cells and then incubated with Streptavidin Magnetic Bead. The immunoprecipitated mixture was analyzed by qRT-PCR assay.
Statistical analyses
All above assays were replicated independently for at least three times. Data are shown as mean ± standard deviation (SD) and analyzed with paired Student's t-test or one-way analysis of variance (ANOVA) using GraphPad PRISM 6 (GraphPad, San Diego, CA). A p-value of <0.05 was used as the threshold of statistical significance.
Results
SNHG7 exhibits oncogenic properties in CC
First of all, the expression of SNHG7 was observed to be notably elevated in CC cells (SiHa, HeLa, C-33A, and CaSki) than in human immortal cervical epithelial H8 cells (Fig. 1A). Then, it was quantified by qRT-PCR that SNHG7 exhibited remarkable lower expression in cells transfected with sh-SNHG7#1/2 (Fig. 1B), indicating that sh-SNHG7#1/2 was qualified to be applied in the following functional assays. In colony formation assay, the number of colonies was detected to be reduced by absence of SNHG7 (Fig. 1C). Moreover, CCK-8 assay suggested that cell proliferative ability was restrained by SNHG7 deficiency (Fig. 1D). Subsequently, EdU assay confirmed that cell proliferation was weakened in sh-SNHG7#1/2-transfected CaSki and C-33A cells (Fig. 1E).
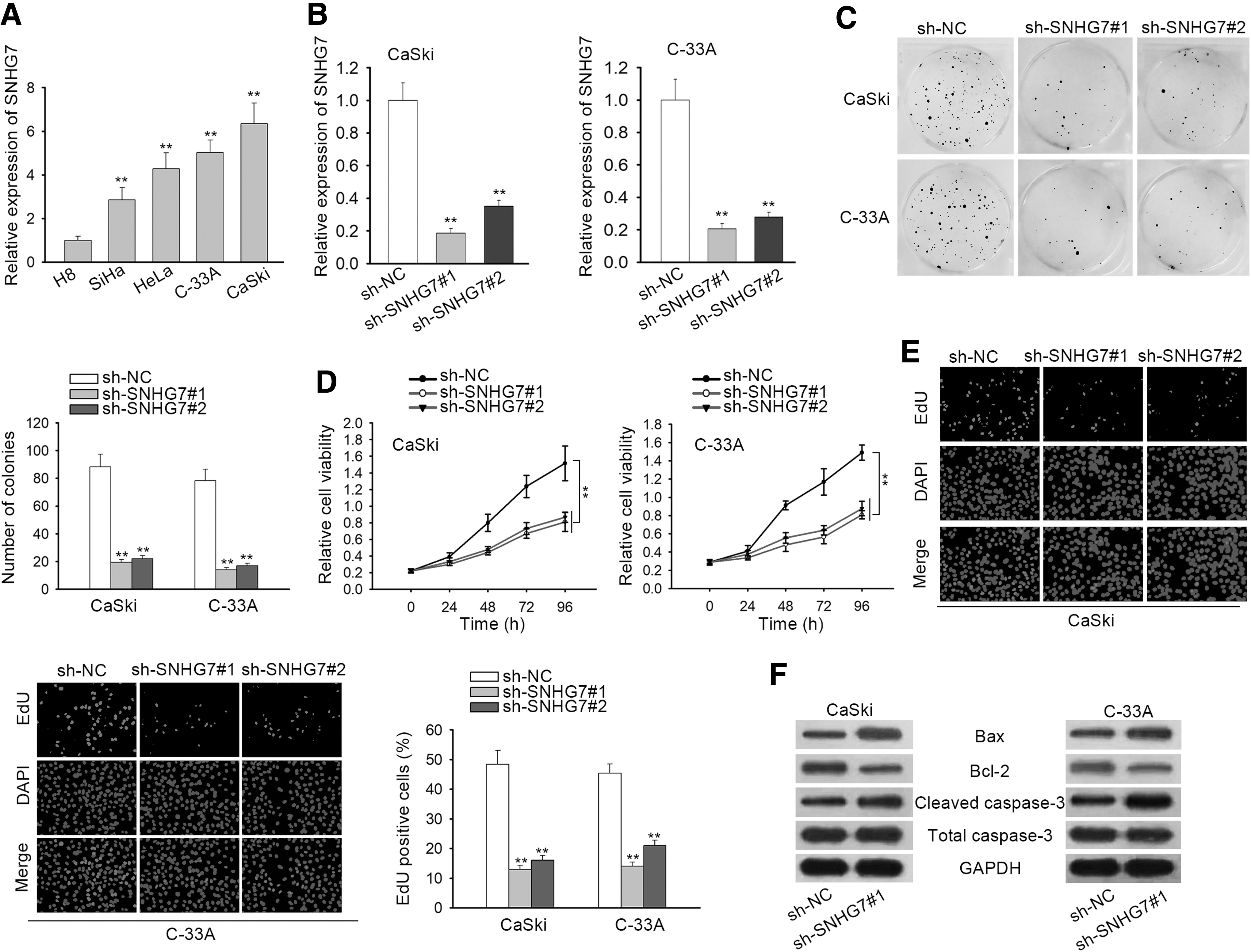
SNHG7 exhibits oncogenic properties in CC.
Finally, Western blot assay demonstrated that the level of Bcl-2 was decreased by SNHG7 depletion while the level of Bax and cleaved caspase-3 was increased by SNHG7 knockdown in CaSki and C-33A cells (Fig. 1F and Supplementary Fig. S2A), indicating that cell apoptosis was inhibited by SNHG7 downregulation. Additionally, SNHG7 expression was reduced by the transfection of sh-SNHG7#1/2 into SiHa and HeLa cells (Supplementary Fig. S1A). Cell proliferation was impaired and cell apoptosis was facilitated by SNHG7 depletion (Supplementary Fig. S1B–E).
SNHG7 upregulates DKK1 expression and activates Wnt/β-catenin signaling pathway
To start, Western blot assay revealed that the expression of Wnt/β-catenin signaling pathway-related proteins (β-catenin, c-myc, and cyclin D1) was suppressed, whereas that of DKK1 (the inhibitor of Wnt/β-catenin signaling pathway) was enhanced in sh-SNHG7#1-transfected CaSki and C-33A cells (Fig. 2A and Supplementary Fig. S2B). TOP/FOP-Flash reporter could be utilized to monitor the concentration of Wnt proteins in conditioned media. 22,23 Then, the luciferase activity of TOP/FOP-Flash reporter in CaSki and C-33A cells was manifested to be markedly attenuated by SNHG7 silencing (Fig. 2B), implying that Wnt/β-catenin signaling pathway was restrained by SNHG7 deficiency. In subsequence, SNHG7 level was enhanced in pcDNA3.1/SNHG7-transfetced cells (Fig. 2C).
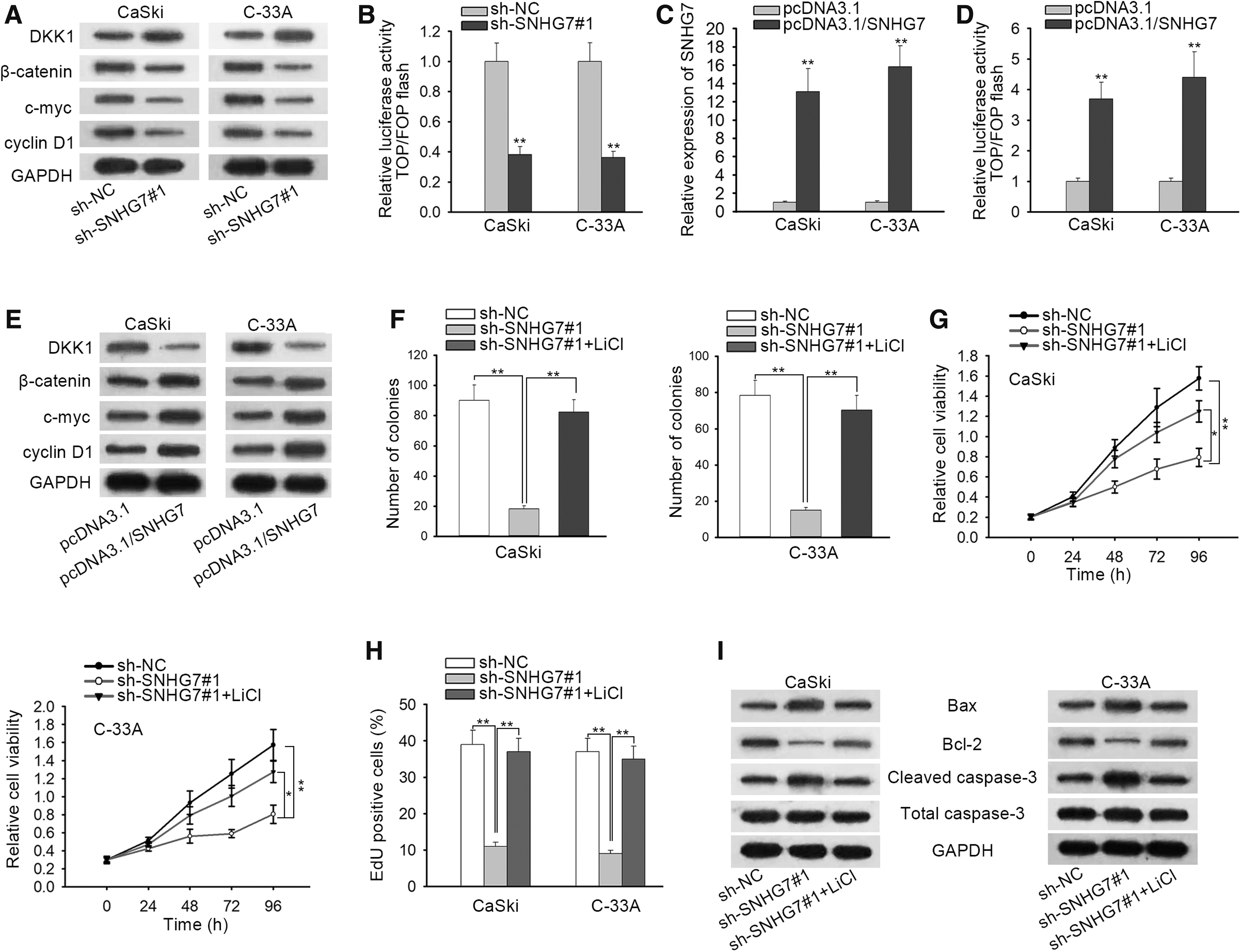
SNHG7 upregulates DKK1 expression and activates Wnt/β-catenin signaling pathway.
Enforced expression of SNHG7 was observed to strengthen the luciferase activity of TOP/FOP-Flash reporter, downregulate DKK1, and upregulate Wnt/β-catenin signaling pathway-related proteins (Fig. 2D, E), implying that SNHG7 could activate Wnt/β-catenin signaling pathway. To confirm that SNHG7 promotes CC development via activating Wnt/β-catenin signaling pathway, LiCl (the activator of Wnt/β-catenin signaling pathway) was employed. Colony formation, CCK-8 assay, and EdU assay verified that the suppressed cell proliferation in CaSki and C-33A cells triggered by SNHG7 inhibition was mostly compensated by the treatment of LiCl (Fig. 2F–H). Furthermore, it was validated by Western blot assay that the treatment of LiCl could countervail the SNHG7 deficiency-mediated enhancive effect on cell apoptosis in CaSki and C-33A cells (Fig. 2I).
SNHG7 reduces the expression of DKK1 via recruiting EZH2 to DKK1 promoter
As described in Figure 3A, DKK1 expression was remarkably reduced by the transfection of sh-DKK1#1/2 into CaSki and C-33A cells, of which sh-NC possessed the higher efficiency. Besides, the absence of DKK1 could enhance the expression of Wnt/β-catenin signaling pathway-related proteins and the luciferase of TOP/FOP-Flash reporter (Fig. 3B, C), confirming that DKK1 could inactivate Wnt/β-catenin signaling pathway. And as the results of subcellular fractionation assay suggested, SNHG7 was occupied in both the nuclear and cytoplasmic part of CC cells (Fig. 3D), suggesting that SNHG7 may participate in the transcriptional or post-transcriptional regulation. FISH data indicated that SNHG7 was primarily situated in the nuclear region of selected CC cells (Fig. 3E).

SNHG7 reduced the expression of DKK1 by recruiting EZH2 to DKK1 promoter.
Previous literature figured out that lncRNAs could recruit EZH2 to epigenetically inhibit the expression of their downstream mRNAs. 19,20 We studied the impacts of SNHG7 on EZH2. Both the mRNA and protein levels of EZH2 were largely unaffected by overexpressing SNHG7 (Fig. 3F). Thus, RIP assay was carried out to research the interaction between SNHG7 and EZH2, and the results implied that SNHG7 could bind with EZH2 in CaSki and C-33A cells (Fig. 3G). RNA pull-down assay further verified the direct physical binding between SNHG7 and EZH2 (Fig. 3H). Next, EZH2 was knocked down, and qRT-PCR results measured that EZH2 level was significantly downregulated after the transfection of sh-EZH2 into CaSki and C-33A cells (Fig. 3I).
Subsequently, the mRNA and protein levels of DKK1 were examined to be conspicuously elevated by EZH2 depletion, as proofed by qRT-PCR and Western blot assay, respectively (Fig. 3J). Furthermore, it was revealed by luciferase reporter assay that SNHG7 deficiency promoted the luciferase activity of DKK1 promoter reporter, implying that the transcription of DKK1 could be suppressed by SNHG7 (Fig. 3K). Finally, ChIP assay was used to study the influence of SNHG7 on the affinity between EZH2 and DKK1 promoter. Its data showed that SNHG7 knockdown remarkably depleted the binding of EZH2 and the occupancy of H3K27me3 in the promoter region of DKK1 (Fig. 3L).
SNHG7 enhances CC development via downregulating DKK1
As shown in Figure 4A and B, the suppressed activity of Wnt/β-catenin signaling pathway in sh-SNHG7#1-transfected cells could be restored by DKK1 inhibition. Then, it can be inferred from the results of colony formation, CCK-8 assay, and EdU assay that the restrained cell proliferative ability induced by the absence of SNHG7 could be restored after LiCl being treated into CaSki and C-33A cells (Fig. 4C–E). In addition, Western blot assay revealed that the treatment of LiCl could largely abrogate the enhanced cell apoptosis caused by SNHG7 depletion (Fig. 4F).
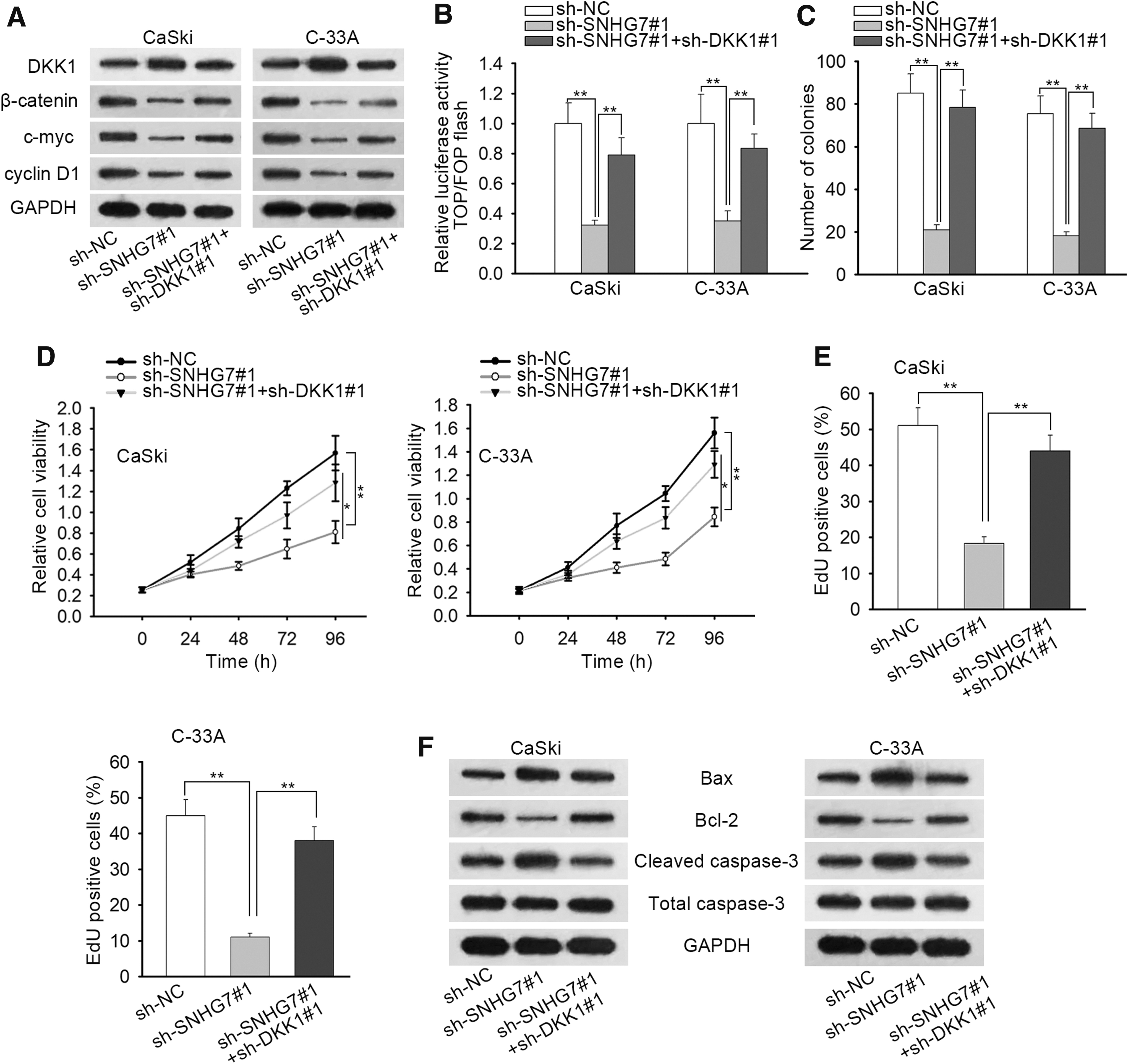
SNHG7 inhibited CC development via downregulating DKK1.
Discussion
It has been established that lncRNAs can regulate gene expression at a variety of levels, such as chromatin modification, transcriptional processing, and post-transcriptional processing. 24,25 Accumulating data has implied that aberrant expression of lncRNAs enable themselves to serve as crucial genes in the carcinogenesis, and lncRNAs could be utilized as biomarkers to diagnose, treat, and prognose cancers. 26,27 SNHG7 has been revealed to promote the development of quite a few cancers. 12,13 In this study, it was discovered that SNHG7 was remarkably overexpressed in CC and exacerbated cell proliferation in CC. To put it more clearly, SNHG7 functions as a malignancy enhancer in CC progression.
As a classical signaling pathway, the activation of Wnt/β-catenin signaling pathway is confirmed to be associated with the biological behavior of cancers, such as liver cancer, 28 pancreatic cancer, 29 and CC. 30
Here, the authors investigated whether Wnt/β-catenin signaling pathway engages in the SNHG7-regulated CC occurrence. In this work, it was revealed that the expression of Wnt/β-catenin signaling pathway-related proteins (β-catenin, c-myc, and cyclin D1) was suppressed, whereas that of dickkopf-1 (DKK1, the inhibitor of Wnt/β-catenin signaling pathway) was enhanced by SNHG7 depletion in CC cells. Besides, the absence of SNHG7 weakened the luciferase activity of TOP/FOP-Flash reporter. More importantly, the introduction of LiCl compensated for the restrained CC cells cellular process induced by SNHG7 deficiency. In brief, SNHG7 reduces DKK1 expression and leads to the activation of Wnt/β-catenin signaling pathway.
Afterward, it was affirmed that DKK1 suppressed the activity of Wnt/β-catenin signaling pathway in CC cells. Therefore, the authors went on to investigate the regulation of SNHG7 on DKK1. To figure out the regulatory mechanism, subcellular fractionation assay was conducted. The results indicated that SNHG7 was located in both the nuclear and cytoplasmic part of CC cells, meaning that SNHG7 might participate in the transcriptional or post-transcriptional regulation. At post-transcriptional level, lncRNAs could act as competitive endogenous RNA or cooperating with RNA binding protein to realize gene modulation. However, at transcriptional level, lncRNAs are capable of modulating the expression of target gene via binding with different kinds of histone modification enzymes, thus influencing cellular functions in human cancers. 31,32
Notably, increasing evidence revealed that about 20% of lncRNAs could realize gene expression silence through binding to EZH2, an important subunit of the polycomb repressive complex 2 (PRC2), which mediates gene expression by methylating lysine 27 on histone H3. For instance, lncRNA LINC00511 was found to bind to EZH2 and further affect the histone modification pattern of the target genes in non-small-cell lung cancer. 33 Previously, DKK1 has been found to be regulated by lncRNA NBAT1 via an EZH2-depedent manner. 34 Hence, the authors hypothesized that SNHG7, which distribute mainly in the nuclear part of selected CC cells, may modulate DKK1 at transcription level.
In this literature, it was unveiled that SNHG7 could bind with EZH2, and EZH2 inversely regulated the level of DKK1. SNHG7 was unearthed to promote the binding of EZH2 to DKK1 promoter and H3K27me3 occupancy in DKK1 promoter, indicating that DKK1 level was suppressed by SNHG7-recruited EZH2. Interestingly, lncRNA LINC00467 was reported to epigenetically silence DKK1, consequently activate Wnt/β-catenin signaling pathway in lung adenocarcinoma. 35
Here, the authors also found that SNHG7 regulate Wnt/β-catenin signaling pathway in a DKK1-dependant way. Finally, for the exploration of whether DKK1 take part in the SNHG7-regulated CC tumorigenesis, several rescue assays were taken out. Results demonstrated that DKK1 takes part in the regulation of SNHG7 on CC occurrence. Eventually, a conclusion can be reached that SNHG7 epigenetically suppresses DKK1 expression to enhance the malignant neoplasia of CC via activating Wnt/β-catenin signaling pathway, offering a new insight into the therapeutic strategies for CC patients.
Footnotes
Acknowledgment
The authors appreciate the support of the First Affiliated Hospital of Soochow University.
Disclosure Statement
The authors declare that no competing interest exists in this study.
Funding Information
This study was supported by 2017 Suzhou industrial technology innovation projection (52010130010021, SYS201750) and Jiangsu maternal and child health research project (F201532).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.