
Abstract
Background:
LncRNA-ZNF281 suppresses glioma, whereas its role in hepatocellular carcinoma (HCC) is unclear. This study aimed to investigate the interaction between ZNF281 and miR-539 in HCC.
Materials and Methods:
This study included 66 HCC patients (40 men and 26 women; 36–68 years, 53.1 ± 6.2 years) who were selected from the 133 HCC cases admitted to The First Affiliated Hospital of Fujian Medical University from February 2011 to June 2014. Levels of ZNF281 and miR-539 expression in two types of tissues (HCC and nontumor) were measured by performing quantitative polymerase chain reaction (qPCR). Dual-luciferase assay was performed to analyze the interactions between miR-539 and ZNF281 in both SNU-475 and PLHC-1 cells. The effects of ZNF281 and miR-539 overexpression on the invasion and migration of HCC cells were analyzed by performing transwell assays.
Results:
The authors showed that ZNF281 was upregulated and miR-539 was downregulated in HCC tissues and were negatively correlated. High levels of ZNF281 and low levels of miR-539 predicted the poor survival of HCC patients. Overexpression analysis showed that ZNF281 and miR-539 overexpression led to the downregulation of each other. Transwell assays showed that ZNF281 overexpression led to enhanced and miR-539 overexpression led to suppressed HCC cell invasion and migration. In addition, miR-539 overexpression attenuated the effects of ZNF281 overexpression.
Conclusions:
Therefore, ZNF281 may interact with miR-539 to promote HCC cell invasion and migration.
Introduction
According to the latest GLOBOCAN statistics, liver cancer in 2018 affected 841,080 new cases, accounting for 4.7% of all new cancer cases. In the same year, liver cancer caused 781,631 deaths, accounting for 8.2% of all cancer deaths. 1 Owing to the extremely aggressive nature, liver cancer now is considered as the third or fourth leading cause of cancer deaths worldwide. 2,3 Hepatocellular carcinoma (HCC) is the major subtype of primary liver cancer and accounts for 85%–90% of all cases. Infections of hepatitis B and C viruses are the major risk factors of HCC. 4,5 However, hepatitis virus infections are not sufficient for the development and progression of HCC, indicating the involvement of genetic factors. 6
A growing body of literature has shown that noncoding RNAs (ncRNAs) are not the “noise” of the transcriptome but critical players in both physiological and pathological processes.
7,8
Analysis of the functionality of ncRNAs may provide novel therapeutic targets for human diseases, including HCC.
9
However, functions of most ncRNAs, such as long noncoding RNAs (lncRNAs) and microRNAs (miRNAs), remain unclear. ZNF281 is a recently identified lncRNA in glioma.
10
The expression data in TCGA data set were analyzed and found that ZNF281 was upregulated in HCC tissues than in nontumor tissues (11.02vs.5, 39;
Materials and Methods
HCC patients and follow-up
This study included 66 HCC patients (40 men and 26 women; 36–68 years, 53.1 ± 6.2 years) who were selected from the 133 HCC cases admitted to The First Affiliated Hospital of Fujian Medical University from February 2011 to June 2014. Ethics Committee of aforementioned hospital approved this study. Inclusion criteria: (1) newly diagnosed HCC cases; (2) with normal functions of other major organs. Exclusion criteria: (1) recurrent HCC; (2) therapies were initiated; (3) multiple disorders were diagnosed. According to AJCC staging system, the 66 HCC patients included 20, 24, and 22 cases at stages II–IV, respectively. According to their medical record, there were 23 cases of HBV-positive, 22 cases of hepatitis-positive, 10 cases of both positive, and 11 cases of both negative. From the day of admission, the 66 HCC patients were monitored for 5 years or until their death to perform survival analysis. The following patients were excluded in survival analysis: (1) patients died of other diseases; (2) patients died of accident; (3) patients lost during follow-up. Informed consent was signed by all patients and controls.
Biopsies and HCC cell lines
Under the guidance of magnetic resonance imaging, liver biopsy was performed to collect both HCC and adjacent (within 2 cm around tumor) nontumor tissue samples. All the tissue samples were subjected to histopathological examinations to confirm that correct samples were harvested.
Two human HCC cell lines SNU-475 and PLHC-1 (ATCC, USA) were included in this study. The cell culture medium was the mixture of 10% fetal bovine serum (FBS) and 90% RPMI-1640 Medium. Cell culture conditions were 37°C, 5% carbon dioxide, and 95% humidity.
Cell transfections
ZNF281 expression plasmid was constructed using pcDNA3.1 vector (Sangon, Shanghai, China). Negative control (NC) small interfering RNA and miR-539 mimic were purchased from RIBOBIO (Guangzhou, China). To perform cell transfections, SNU-475 and PLHC-1 cells were harvested at 70%–80% confluence and lipofectamine 2000 (Sangon) was used to transfect 12 nM vector (empty pcDNA3.1 vector as NC group) or 40 nM miRNA (NC miRNA and NC group) into 3 × 106 cells. Untransfected cells were used as Control (C) cells. Following experiments were performed at 24 h post-transfection.
RNA extractions
SNU-475 and PLHC-1 cells were harvested at 24 h post-transfection and cells were counted. Liver biopsies were ground in liquid nitrogen. RNAzol (Sigma-Aldrich, USA) was used to extract total RNAs from 3 × 105 cells and 0.02 g tissue samples with all operations performed according to the instructions from Sigma-Aldrich. miRNAs were harvested by precipitating RNAs using 85% ethanol.
Quantitative polymerase chain reaction assay
DNase I was used to treat all RNA samples to remove genomic DNAs. MMLV Reverse Transcriptase 1st-Strand cDNA Synthesis Kit (Lucigen) was used to reverse transcribe RNA samples into computed tomography (cDNA). With cDNA as template, quantitative polymerase chain reaction (qPCR) assays were performed using TB Green Advantage qPCR Premix (Clontech, USA). With 18S rRNA as endogenous control, the expression levels of ZNF281 were normalized.
The expression levels of mature miR-539 were also measured. The addition of poly (A), miRNA reverse transcription, and miRNA qPCR assays were all performed using All-in-One™ miRNA qRT-PCR Detection Kit (Genecopoeia).
All Ct values were processed using 2−ΔΔCT method. All PCRs were performed in triplicate manner.
Dual-luciferase assay
The interaction between miR-539 and ZNF281 was analyzed by dual-luciferase assay. In this assay, psiCHECK-1 vector (Promega) instead of pcDNA3.1 vector was used. Cells were transfected with either ZNF281 vector + NC miRNA or ZNF281 vector + miR-539. Dual-Luciferase Reporter Assay (Promega) was used to normalize renilla luminescence by firefly luminescence.
Cell invasion and migration analysis
SNU-475 and PLHC-1 cells were harvested at 24 h post-transfection and cells were counted. After that 1 mL serum-free RPMI-1640 Medium was used to resuspend cell pellets containing 3 × 104 cells to make cell suspensions (single cell). To perform transwell assays, the upper chamber was filled with cell suspensions, whereas the lower chamber was filled with a mixture of 20% FBS and 80% RPMI-1640 Medium. It is worth noting that, before invasion assay, upper chamber membranes were coated with Matrigel (250 mg/mL; Millipore, USA) for 12 h at 37°C. Under aforementioned conditions, cells were cultivated for 12 h, following by using crystal violet (0.1%, Sigma-Aldrich) to stain the lower face of the membranes for 20 min at room temperature. Finally, a light microscope was used to observe and count invading and migrating cells.
Statistical analysis
Mean values of three replicates were calculated and used in the following statistical analysis. Correlations were analyzed by Pearson's correlation coefficient. Differences between two types of tissues or two groups were analyzed by paired or unpaired t-test, respectively. Differences among multiple groups were analyzed by analysis of variance (one-way) and Tukey test. With the median expression levels of ZNF281/miR-539 as cutoff values, the 66 patients were grouped into high- and low-level groups (n = 33). Survival curves were plotted using K–M plotter and compared by log-rank test. p-Value <0.05 was statistically significant.
Results
ZNF281 and miR-539 were inversely correlated in HCC
Levels of ZNF281 and miR-539 expression in two types of tissues (HCC and nontumor) were measured by performing qPCR. Expression data were compared between 2 groups by performed paired t-test. It was observed that expression levels of ZNF281 were significantly higher in HCC tissues than in nontumor tissues (Fig. 1A, p < 0.05). In contrast, expression levels of miR-539 were significantly lower in HCC tissues than in nontumor tissues (Fig. 1B, p < 0.05). The correlations between ZNF281 and miR-539 across HCC tissues (Fig. 1C) and nontumor tissues (Fig. 1D) were analyzed by performing Pearson' correlation coefficient. It was observed that ZNF281 and miR-539 were inversely and significantly correlated in HCC tissues but not in nontumor tissues.
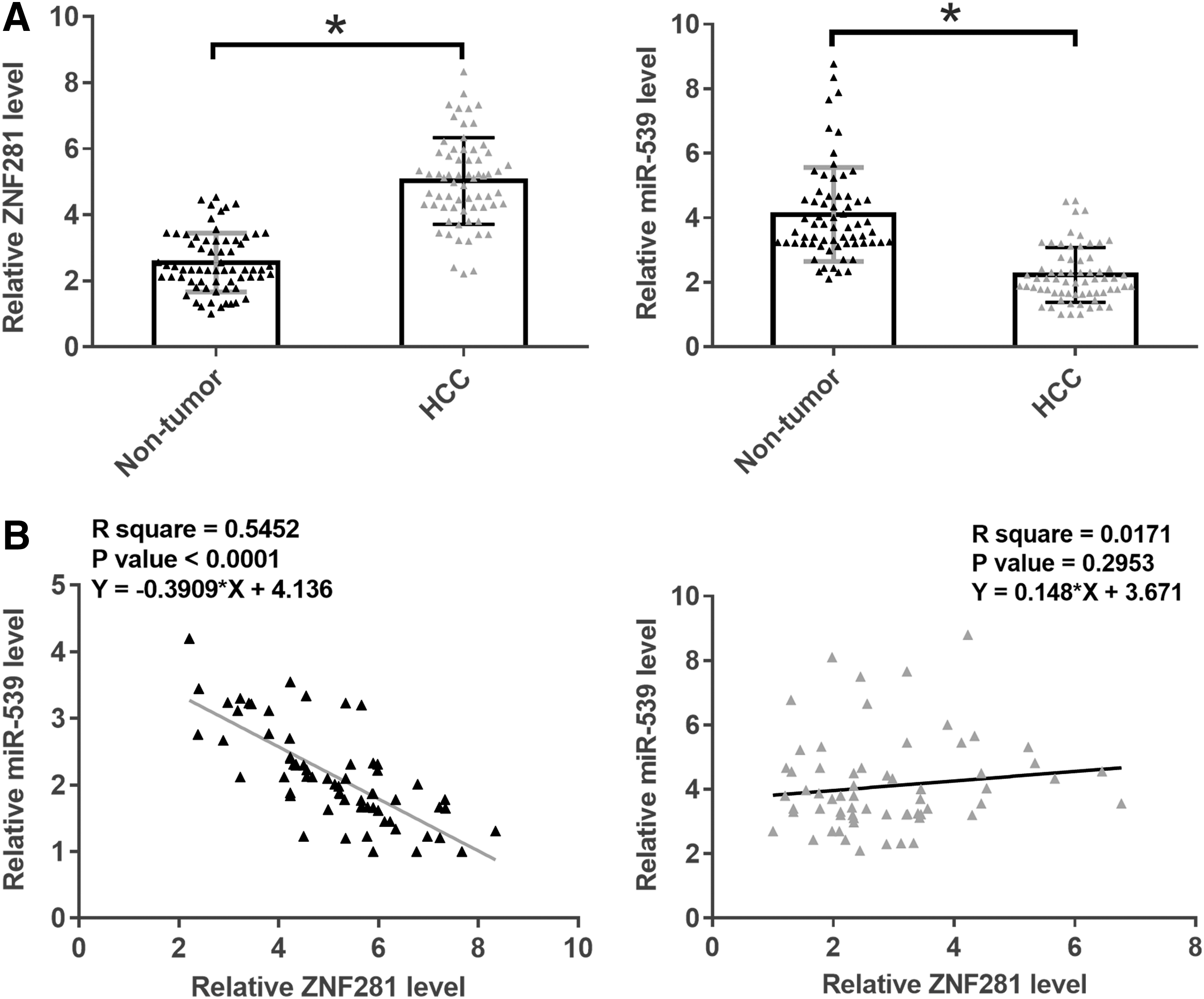
ZNF281 and miR-539 were inversely correlated in HCC. Levels of ZNF281 and miR-539 expression in two types of tissues (HCC and nontumor) were measured by performing qPCR. Expression data were compared between 2 groups by performed paired t-test
Altered expression levels of ZNF281 and miR-539 predicted poor survival
Survival curves were plotted and compared using aforementioned methods. Comparing with patients in low ZNF281 level groups, patients in high ZNF281 level groups showed significantly lower overall survival (Fig. 2A). However, the overall survival rate of patients in low miR-539 group was significantly lower than that in high-level group (Fig. 2B).
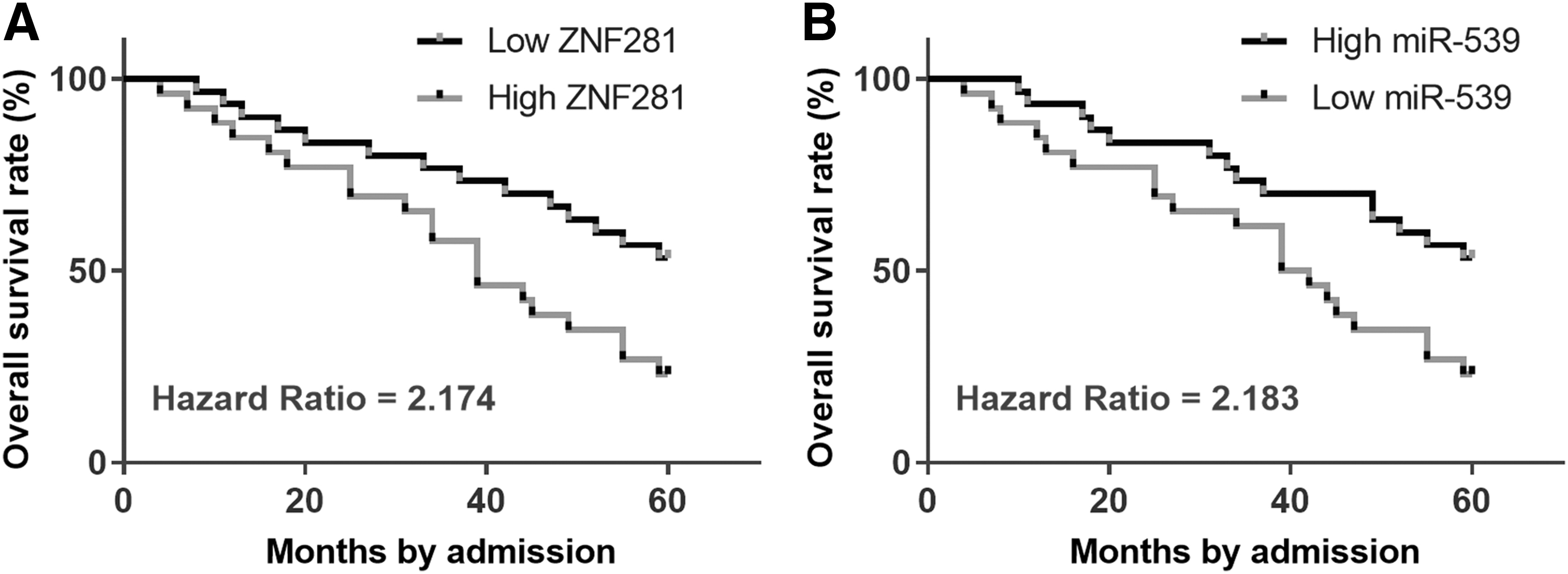
Altered expression levels of ZNF281 and miR-539 predicted poor survival. With the median expression levels of ZNF281
ZNF281 and miR-539 downregulated each other in HCC cells
Dual-luciferase assay was performed to analyze the interactions between miR-539 and ZNF281 in both SNU-475 and PLHC-1 cells. Comparing with cells transfected with ZNF281 vector + NC miRNA (NC group), significantly higher luciferase activity was observed in cells transfected with ZNF281 vector + miR-539 (miR-539 group) (Fig. 3A, p < 0.05). To further analyze the relationship between these two factors, SNU-475 and PLHC-1 cells were transfected with ZNF281 expression vector and miR-539 mimic. Comparing with C and NC groups, expression levels of ZNF281 and miR-539 were significantly increased at 24 h post-transfection (Fig. 3B, p < 0.05). Comparing with two controls, ZNF281 overexpression led to the downregulation of miR-539 (Fig. 3C, p < 0.05). In addition, miR-539 overexpression also led to the downregulation of ZNF281 (Fig. 3D, p < 0.05).
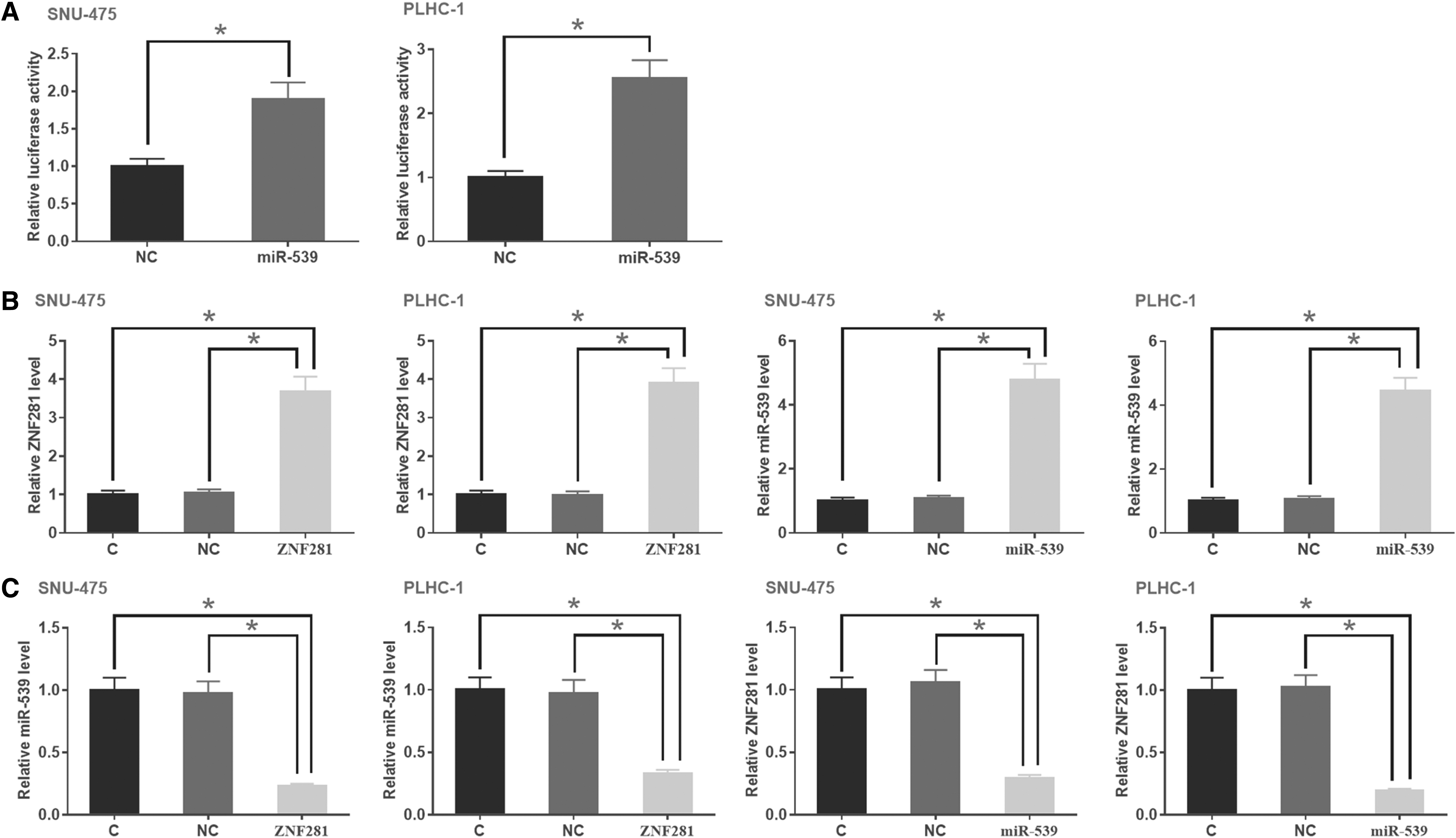
ZNF281 and miR-539 downregulated each other in HCC cells. Dual-luciferase assay was performed to analyze the interactions between miR-539 and ZNF281 in both SNU-475 and PLHC-1 cells. Comparing with cells transfected with ZNF281 vector + NC miRNA (NC group), significantly higher luciferase activity was observed in cells transfected with ZNF281 vector + miR-539 (miR-539 group)
The interaction between ZNF281 and miR-539 regulated the invasion and migration of HCC cells
The effects of ZNF281 and miR-539 overexpression on the invasion (Fig. 4A) and migration (Fig. 4B) of HCC cells were analyzed by performing transwell assays. Comparing with C and NC groups, ZNF281 overexpression led to enhanced and miR-539 overexpression led to suppressed HCC cell invasion and migration. In addition, miR-539 overexpression attenuated the effects of ZNF281 overexpression.
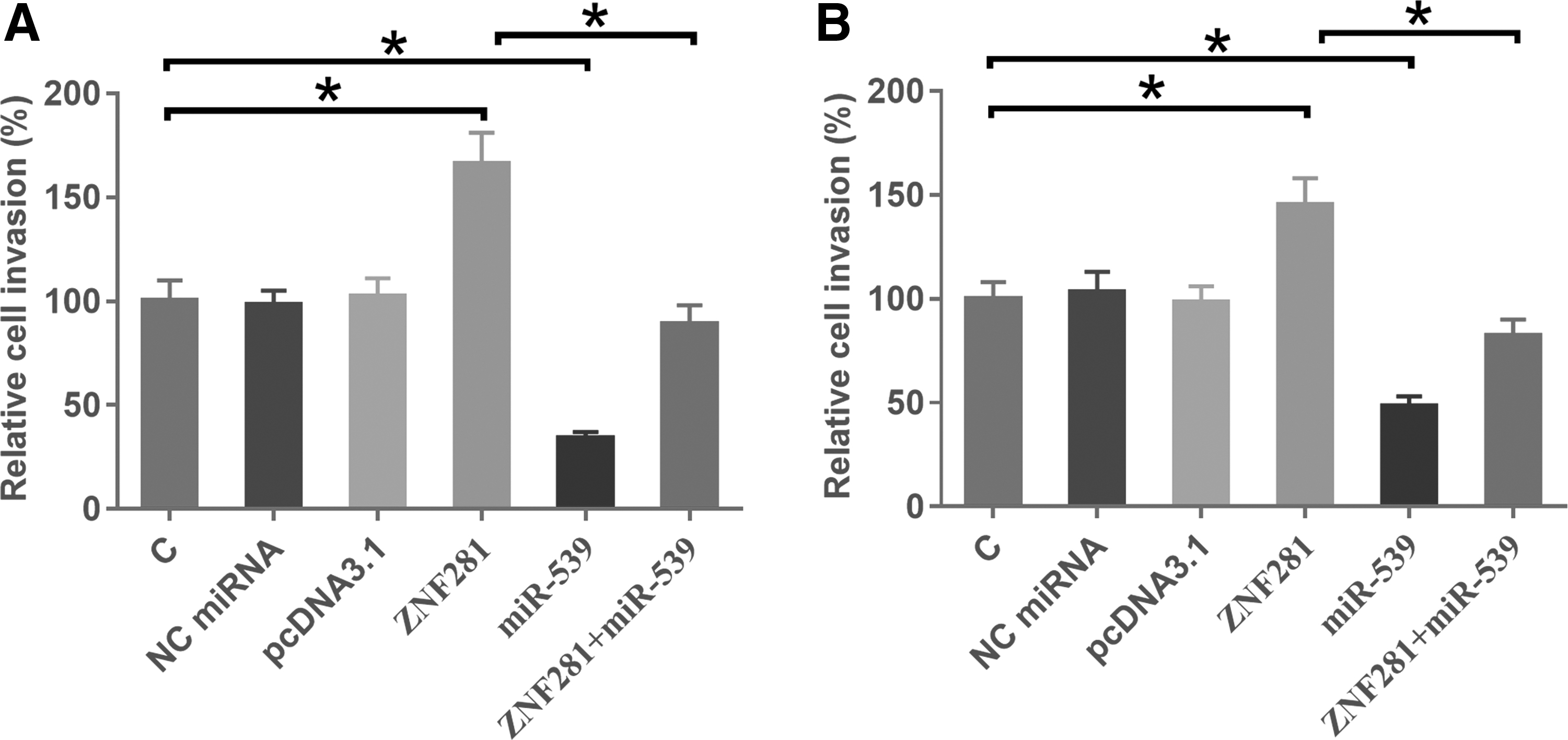
The interaction between ZNF281 and miR-539 regulated the invasion and migration of HCC cells. The effects of ZNF281 and miR-539 overexpression on the invasion
Discussion
The expression pattern and functions of ZNF281 in HCC were explored in this study. It was found that ZNF281 was upregulated in HCC and promotes cancer cell invasion and migration. The authors also proved that the functions of ZNF281 in HCC are likely mediated by miR-539.
A previous study reported the downregulation of ZNF281 in glioma cells, and in vitro cell experiments revealed is inhibitory effects of ZNF281 on the proliferation, migration, and invasion of glioma cells.
10
Therefore, ZNF281 is a tumor suppressor in glioma. However, by analyzing available TCGA data set the authors found that ZNF281 was upregulated in glioma (GBM;
By literature review it was found that an lncRNA may play similar roles in different types of cancers. 12,13 For instance, lncRNA HORAIR as a master regulator of chromatin dynamics promotes the development of most, if not all types of cancer. 14 However, lncRNA TUG1 suppresses glioma but promote most types of cancer. 15 In this study the authors showed the upregulation of ZNF281 and its enhancing effects on HCC cell invasion and migration. Therefore, ZNF281 plays oncogenic roles in HCC. The different functions of lncRNA TUG1 and ZNF281 in glioma is likely mediated by the unique pathogenesis of this disease. 16
In this study the authors showed that ZNF281 and miR-539 can interact with each other to form a negative regulation loop. The authors observed altered expression of ZNF281 and miR-539 was closely correlated with poor survival, whereas deaths in cancer patients are mainly caused by metastasis. 17 The authors' following in vitro cell experiments showed that this negative regulation loop participated in the regulation of invasion and migration of HCC cells. However, the mechanisms that mediate the interaction between these two factors remain unclear. The authors observed that ZNF281 and miR-539 were not significantly correlated in HCC. Therefore, certain disease factors may mediate the interaction between them. The authors' future studies will explore these mediators.
Conclusion
In conclusion, ZNF281 is upregulated in HCC and forms a negative feedback regulation loop with miR-539 to regulate HCC cell invasion and migration.
Ethics Approval and Consent to Participate
Ethical approval was obtained from the Ethics Committee of The First Affiliated Hospital of Fujian Medical University. Written informed consent was obtained from all individual participants included in this study.
Footnotes
Authors' Contributions
Z.B.Z. performed the experiments study, data analysis, funding acquisition, and article writing. L.F.Y., X.Q.Y., M.Z.Y., and G.P.L. did the experiment work, literature research, and clinical research. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank the financial support from National Key Clinical Speciality Construction Project (2016-SLCZD); Traumatic Medical Center of Fujian Province (HBPS).