
Abstract
Background:
X-linked inhibitor of apoptosis protein (XIAP) is the strongest member of the family of inhibitor of apoptosis protein. Studies found that the expression of XIAP in colon cancer tissue was significantly higher than that in adjacent tissues. Studies have shown that the expression of microRNA-215 (miR-215) was significantly lower than that of the adjacent tissues. This study investigated whether dysregulated miR-215 and XIAP play important roles in colon cancer cell apoptosis and the incidence of colon cancer.
Materials and Methods:
Forty-two patients with colorectal cancer (CRC) diagnosed and treated in the authors' hospital were selected. Human CRC cell line HCT116 and normal colonic mucosal epithelial cells (CMECs) were used. Luciferase reporter gene vector was constructed and dual-luciferase reporter gene assay was performed. HCT116 cells were cultured in vitro and divided into five groups: mimic normal control (NC) group, miR-215 mimic group, si-NC group, si-XIAP group, and miR-215 mimic + si-XIAP group. Western blot and polymerase chain reaction were conducted to examine XIAP and caspase-3. Apoptosis was detected by flow cytometry and cell proliferation was detected by cell counting kit-8 assay.
Results:
Compared with the adjacent tissues, the expression of miR-215 in colon cancer tissue was significantly lower, whereas the expression of XIAP in colon cancer tissue was significantly higher. The apoptosis rate and miR-215 expression level of HCT116 cells were lower than that of normal CMECs, whereas XIAP expression was significantly higher than that in normal colon mucosa epithelial cells. MiR-215 targeted the 3′-untranslated regions of XIAP and inhibited its expression. Overexpressing miR-215 and (or) silencing XIAP expression could significantly enhance the activity of caspase-9 and caspase-3, and promote the apoptosis of HCT116 cells.
Conclusion:
MiR-215 inhibited the expression of XIAP and promoted the apoptosis of HCT116 cells.
Introduction
Colorectal cancer (CRC) is a common malignant tumor of gastrointestinal tract occurring in the rectum or sigmoid colon, and the morbidity rate of males to females is 2:1–3:1, with the highest morbidity rate in the 40- to 50-year age group. 1 X-linked inhibitor of apoptosis protein (XIAP) is the strongest member among the inhibitor of apoptosis proteins (IAPs), which plays an antiapoptotic role by inhibiting caspase activity, and its abnormally increased expression is associated with the occurrence and progression of tumors and drug resistance. 2,3
Xiang et al.'s studies showed that XIAP expression in the CRC tissues and tumor cell lines was significantly higher than that in paracancerous tissues and normal colonic epithelial cells, and its expression level was associated with the survival and prognosis of patients. 4 Studies by Takeuchi et al. revealed that the expression level of XIAP in CRC tissues was closely associated with the progression and metastasis of the tumor, suggesting that XIAP overexpression may be involved in CRC occurrence. 5
MicroRNA, a small noncoding RNA of about 18–22 nucleotides in eukaryotes, is one of the important contents of epigenetic regulation, which can regulate the expressions of >30% human genes. Through complete or incomplete complementary pairing with target gene messenger RNA (mRNA), it can promote mRNA degradation of target gene or inhibit its translation, which involves in the regulation of biological processes such as cell proliferation, cycle, differentiation, and apoptosis. 6 During the tumorigenesis, microRNAs act as a dual role of tumor suppressor and promoter by inhibiting the expression of carcinogenic gene and antioncogene, and its expression and functional abnormality are related to the occurrence of various tumors. 7,8
The microRNA-215 (miR-215) expression in tumor tissues of CRC patients was significantly lower than that in the paracancerous tissues, 9,10 suggesting that miR-215 expression may affect the occurrence of colon cancer. It was reported that XIAP was regulated by microRNA-137 and thus promoting apoptosis in ovarian cancer cells, as well as other kinds of microRNA also had this effect. 11 The authors first chose miR-215, with an ability of inhibiting XIAP expression strongly, for further confirmation in CRC. In this experiment, the relationships of the expression levels of miR-215 and XIAP in CRC patients with clinicopathological features were detected, and bioinformatics analysis revealed that there was a good targeted complementary relationship between miR-215 and 3′-untranslated regions (UTR) of XIAP. Moreover, they investigated whether miR-215 and XIAP expression disorder played a role in regulating apoptosis of CRC cells.
Materials and Methods
Main reagents and materials
Human colonic cancer cell line HCT116 was purchased from Shanghai Yu Bo Biological Technology, and normal colonic mucosal epithelial cell (CMEC) was purchased from Shanghai SUER company. Roswell Park Memorial Institute (RPMI) 1640, Dulbecco's modified Eagle medium (DMEM), fetal bovine serum (FBS), and streptomycin were purchased from Gibco. Trizol and Lipofectamine 2000 were purchased from Invitrogen; reverse transcription kits and Synergy Brands (SYBR) dye were purchased from Japan Toyobo; microRNA nucleotide fragments and polymerase chain reaction (PCR) primers were designed and synthesized by Guangzhou Ribobio; Rabbit antibodies against anti-β-actin and XIAP were purchased from Abcam (Cambridge, MA). The mouse antibodies anticleaved caspase-9, anticleaved caspase-3, and the secondary antibody were purchased from Cell Signaling (Danvers, MA). Apoptosis of tumor cells was assessed by Annexin V/propidium iodide (PI) Apoptosis detection reagents (Yeasen Biological, Shanghai, China) and caspase-9, caspase-3 Activity Assay Kits (Beyotime, Shanghai, China). Dual-Luciferase® Reporter Assay System and pGL3-promoter plasmids were purchased from Promega.
Clinical data
In this experiment, samples from 42 patients with CRC, of which the pathologies are adenocarcinoma, treated in the authors' hospital from October 2015 to May 2016 were selected. The specimen from each patient was collected from the operation and confirmed by histopathology, including tumor tissues and their corresponding nontumor adjacent tissues at 5 cm near the cancer. The in vitro fresh samples were quickly stored in supernatant nitrogen in 10 min, and then stored at −80°C for standby. All samples were collected with informed consent of patients and approved by the Ethics Committee of the hospital.
Cell culture
Human CRC cell line HCT116 and normal CMECs were cultured in RPMI 1640 medium containing 10% FBS and 1% streptomycin in an incubator (37°C, 5% carbon dioxide), and the liquid was changed once every 2 d. Experiments were carried out when cells grew reaching at 70%–80% confluence.
Construction of luciferase reporter gene vector
To explore the molecular mechanism of miR-215 in CRC apoptosis, potential target genes of miR-215 could be predicted by accessing to
Dual-luciferase reporter gene assay
Four hundred nanograms pGL3-PITX2-3′-UTR, 25 nmol miR-215 mimic (or negative control), and 25 ng pRL-TK were mixed in Opti-MEM and transfected into HEK293 cells by Lipofectamine 2000. Four to 6 h later, the Opti-MEM was discarded and replaced with DMEM medium containing 10% FBS and 1% streptomycin for culture 48 h continuously. Steps for dual-luciferase activity test: cells were rinsed by phosphate-buffered saline (PBS) twice, added to 100 μL phospholamban lysis solution, shaken 30 min at room temperature, centrifuged 5 min at 1000 rpm, and then 100 μL LAR II was added to 20 μL cell lysate supernatant to determine the fluorescence value I with a microplate reader, and 100 μL Stop & Glo was added to stop the reaction to immediately determine the fluorescence value II. The ratio of fluorescence value I/fluorescence value II was used as the relative luciferase activity.
Grouping and cell transfection
HCT116 cells were cultured in vitro and divided into five groups: mimic normal control (NC) group, miR-215 mimic group, si-NC group, si-XIAP group, miR-215 mimic + si-XIAP group. The nucleotide fragments and Lipofectamine 2000 were diluted with Opti-MEM and incubated at room temperature for 5 min. Then, the nucleotide fragments were mixed with Lipofectamine 2000 and gently mixed upside down. After incubation at room temperature for 30 min, the cells were added to serum-free cell culture medium; 6 h later, the medium was changed to RPMI 1640 medium containing 10% FBS and 1% streptomycin for continuous culture 48 h, and then cells were collected for testing. All nucleotide sequences mentioned earlier were mimic NC, 5′-UUCUCCGAACGUGUCACGUTT-3′; miR-497 mimic, 5′-AUGACCUAUGAAUUGACAGAC-3′; si-XIAP sense strand, 5′-GTGGTAGTCCTGTTTCAGC-3′; si-XIAP antisense strand, 5′-GCTGAAACAGGACTACCAC-3′; si-NC sense strand, 5′-UUCUCCGAACGUGUCACGUTT-3′; si-NC antisense strand, 5′-ACGUGACACGUUCGGAGAATT-3′.
Detection of the expression levels of XIAP and miR-215 by quantitative real-time polymerase chain reaction
XIAP and miR-215 expression levels were further determined by quantitative real-time polymerase chain reaction (qRT-PCR) analyses of 42 patients' CRC tissues and corresponding paracancerous tissues and HCT116 cells. The total RNAs of cells were extracted by the Trizol method. The specific steps: cells were lysed with Trizol, extracted with chloroform in layers, RNA was precipitated by isopropanol, centrifuged and washed with 70% ethanol and then dissolved in diethypyrocarbonate. Ten microliters of reverse transcription reaction system (total volume) included 1 μg of total RNA, 2 μL of RT Buffer (5 × ), 0.5 μL of oligo dT + Random primer Mix, 0.5 μL of RT Enzyme Mix, 0.5 μL of RNase inhibitor, added with double distilled water (ddH2O) to 10 μL, with the reaction condition of 37°C for 15 min, 98°C for 5 min and the resulting complementary RNA (cDNA) was stored in −20°C refrigerator for standby. Using cDNA as a template, the PCR amplification reaction was performed under the action of Taq DNA polymerase. The primer sequences used were: XIAP-forward: 5′-ACTTCCCAAGTAGTAGTCCTGTT-3′, XIAP-reverse: 5′-TATTGCCGCATGACAACTGAA-3′; miR-215-forward: 5′-CTCGAGATGTCATCCTCAG-3′, miR-215-reverse: 5′-CCTGCTACAACCACCTTCTTG-3′. In a PCR system with a total volume of 10 μL, 4.5 μL of 2 × SYBR Green Mixture, 1.0 μL of 2.5 μm/L forward primer, 1.0 μL of 2.5 μm/L reverse primer, 1 μL of cDNA, and 3.0 μL of ddH2O were added up to 10.0 μL. The PCR conditions: 95°C for 15 s, 60°C for 30 s, 74°C for 30 s, and a total of 40 cycles in Bio-Rad CFX96 quantitative PCR system, then the fluorescence data were collected.
Western blot
The level of XIAP expression was measured by Western blotting analysis and samples were normalized to β-actin. Cells and tissue samples were lysed by radio immunoprecipitation assay, placed still on the ice for 30 min, centrifuged 10 min at 10,000 rpm, to get the supernatant and detect the protein mass concentration. Fifty micrograms protein was loaded onto an 8% polyacrylamide gel electrophoresis for electrophoresis separation (voltage: 45 V, time: 300 min), then transferred to polyvinylidene difluoride membranes by electroporation for 100 min (current: 300 mA). The membranes were blocked with 5% skimmed milk for 60 min, followed by incubation with primary antibody overnight at 4°C, then washed by phosphate buffer solution (PBST) three times, 5 min each time, and then horseradish peroxidase (HRP)-labeled secondary antibody was added to incubate 60 min at room temperature, then washed by PBST three times, 5 min each time. After disposed with electrochemiluminescence solution in the dark room, the membranes were subsequently visualized with an enhanced chemiluminescence system kit, and gray intensity analysis of the protein bands was performed using the Quantity One software.
Detection of the expression levels of XIAP in 42 patients' specimens by immunohistochemistry
The procedure was carried out under the instruction of the immunohistochemistry (IHC) SP-9001 Kit (Beijing Zhongshan Golden Bridge Biological Technology, Beijing, China). After dewaxing of the paraffin sections, the endogenous peroxidase was inactivated with 3% hydrogen peroxide. Then, the citrate buffer was used for thermal remediation and proteins were blocked with 10% goat serum. Primary rabbit XIAP antibody (dilution, 1:400) were added and incubated at 4°C overnight. Then the authors washed the slices with PBS and added secondary goat antirabbit (HRP) immunoglobulin G antibody (dilution, 1:2000) and incubated for 15 min. After that, sections were washed with PBS and diaminobenzidine solution was used for development in the dark. Then hematoxylin was used for restaining the samples. Finally, they took pictures under the light microscope and selected three representative regions for each sample. According to the percentage of positive cells and the staining intensity, immunohistochemical staining of 42 × 2 samples were evaluated by rating and grading the expression. The number of positive cells (magnification, × 400) was classified as <5%, 5%–25%, 26%–50%, and >50% (recorded as 0, 1, 2, and 3 points), and the staining intensity was classified as stainless, brownish brown, tan, and sepia (recorded as 0, 1, 2, and 3 points, respectively). Finally, the scores of the two indexes were added, and total scores >3 were regarded as positive expression and ≤2 as negative expression.
Detection of caspase-3 and caspase-9 activity
Ten millimolar pNA was diluted to 0, 10, 20, 50, 100, and 200 μM as standards. The A405 was measured by a microplate reader and a standard curve of pNA concentration relative to A405 was produced. The cell culture medium was absorbed for standby. Adherent cells were digested by trypsin and collected into the spare cell culture medium, centrifuged 5 min at 600 g, 4°C. The supernatant was absorbed carefully, washed once with PBS, to discard the supernatant, then 100 μL lysate was added to 2 × 106 cells, to lyse 15 min at 4°C, then centrifuged 10 min at 4°C, 18,000 g, then the supernatant was transferred to the precooled Eppendorf tube for standby. The steps for the detection of caspase-3 enzymatic activity: appropriate amount of Ac-DEVD-pNA was taken and placed on ice, the detection buffer solution was added to the reaction system, and then test samples were added, followed by 10 μL of Ac-DEVD-pNA was added to mix well, and then incubated 120 min under the condition of 37°C. When the color change was obvious, A405 was determined. The steps for the detection of caspase-9 enzymatic activity: appropriate amount of Ac-DEVD-pNA (2 mM) was taken and placed on ice, the detection buffer solution was added to the reaction system, and then test samples were added, followed by 10 μL of Ac-DEVD-pNA was added to mix well, and then incubated 120 min under the condition of 37°C. When the color change was obvious, A405 was determined.
Apoptosis detection by flow cytometry
Cells were digested by ethylenediaminetetraacetic acid-free trypsin. After washing twice with PBS, 100 μL 1 × Binding Buffer was added, and after gently mixing, 5 μL Annexin V-fluorescein isothiocyanate and 5 μL PI were added and gently mixed for reaction 10 min in the darkness, and then 400 μL of 1 × Binding Buffer was used to resuspend cells, and tested on the machine immediately.
Cell proliferation assay
HCT116 cells were counted and plated in 96-well plates at 2000 cells per well, 24 h later treated by mimic NC; miR-215 mimic; si-NC; si-XIAP; miR-215 mimic + si-XIAP for 3 d, each group with 6 repeated wells. Ten microliters of cell counting kit (CCK)-8 reagent (CCK-8; Dojindo Laboratories, Japan) was added into each well and incubated for 30 min. Absorbance at 450 nm was detected and each experiment was performed in triplicate for three times.
Statistical analysis
Data statistical analysis was performed by SPSS 18.0. Measurement data that met normal distribution were expressed as mean ± standard deviation. The measurement data between groups that met normal distribution were compared using t-test. The expression levels of miR-215, XIAP in colon cancer and paracancerous tissues were compared by Mann–Whitney U-test. The positivity of expression of XIAP in CRC and tumor-adjacent normal tissues was analyzed by McNemar's test. The correlation between miR-215 and XIAP mRNA expression levels was analyzed by Spearman rank correlation analysis. p-Value <0.05 was considered statistically significant difference.
Results
Sample and patients information
Among 42 patients, 20 male cases and 22 female cases, at the age of 46–75 years, 53.9 years on average, there were no statistically significant differences in age and gender between them. All CRC patients were diagnosed and staged based on the seventh edition of tumor-node-metastasis (TNM) staging criteria of the American Joint Committee on Cancer. All the patients were below III stage. There was no distance metastasis in all patients. None of the patients received chemotherapy, radiotherapy, or other special treatments for cancer before operation.
The expression of miR-215 was decreased and the expression of XIAP was increased in CRC tissues
From qRT-PCR results of 42 samples, including CRC tissues and nontumor tissues, the authors found that miR-215 levels of CRC tissues were significantly downregulated (Fig. 1A), whereas XIAP mRNA expression levels were significantly upregulated (Fig. 1B), compared with the paracancerous tissues. Western blot analysis of tumor tissues and nontumor adjacent tissues from 15 patients showed that the protein level of XIAP in the CRC tissues was significantly higher than that in the paracancerous tissues (Fig. 1C). The IHC scores were counted, the positive expression rates of XIAP in CRC and tumor-adjacent normal tissues were 76.2% (32/42) and 23.8% (10/42), respectively, and the difference was statistically significant (p < 0.05) (Table 1). Spearman correlation analysis showed that there was a negative correlation between miR-215 and XIAP mRNA expression (r = −0.723, p = 0.033), suggesting that miR-215 may has an ability of downregulating XIAP expression. In addition, the relationships of miR-215 and XIAP with clinicopathological features of CRC patients were analyzed, suggesting that miR-215 and XIAP expression levels are associated with lymph node metastasis, degrees of differentiation and TNM staging of CRC patients (p < 0.05; Table 2), whereas associations of them with age and gender had no statistically significant differences (p > 0.05).

Expression of miR-215 and XIAP in colon cancer tissues and nontumor adjacent tissues of 42 patients. Determination of the expression of miR-215
Positivity of Expression of X-Linked Inhibitor of Apoptosis Protein in Colorectal Cancer and Tumor-Adjacent Normal Tissues (n = 42)
XIAP, X-linked inhibitor of apoptosis protein.
Associations of X-Linked Inhibitor of Apoptosis Protein and MicroRNA-215 Expression Levels with Clinicopathological Data of Colorectal Cancer Patients (Mean ± Standard Deviation, n = 42)
miR-215, microRNA-215; TNM, tumor-node-metastasis; XIAP, X-linked inhibitor of apoptosis protein.
MiR-215 inhibited the expression of XIAP by targeting its 3′-UTR in CRC cells
Compared with CMEC, miR-215 expression of CRC cell line HCT116 was significantly decreased, whereas XIAP expression was significantly increased (Fig. 2A, B). Compared with CMEC background apoptosis rate at 7.4%, HCT116 cells had a lower background apoptosis rate about 3.7% (Fig. 2C). According to

MiR-215 inhibited the expression of XIAP in colon cancer cell line HCT116.
MiR-215 promoted the apoptosis of CRC cells by inhibiting XIAP expression
In HCT116 cells, miR-215 mimics-mediated upregulation of miR-215 and/or small interfering RNA-induced downregulation of XIAP could significantly reduce mRNA and protein levels of XIAP (Fig. 3A, B), significantly enhance the activity of caspase-9 and caspase-3 (Fig. 3B, C) and increase apoptosis rate (Fig. 3D). Results of CCK-8 assay also showed that inhibition rate of cells were the highest in miR-215 mimics + si-XIAP group compared with other groups (Fig. 3E). Moreover, the inhibition effect on HCT116 cells growth was more distinct when inhibition of XIAP and upregulation of miR-215 worked together, compared with inhibition of XIAP or upregulation of miR-215 alone.
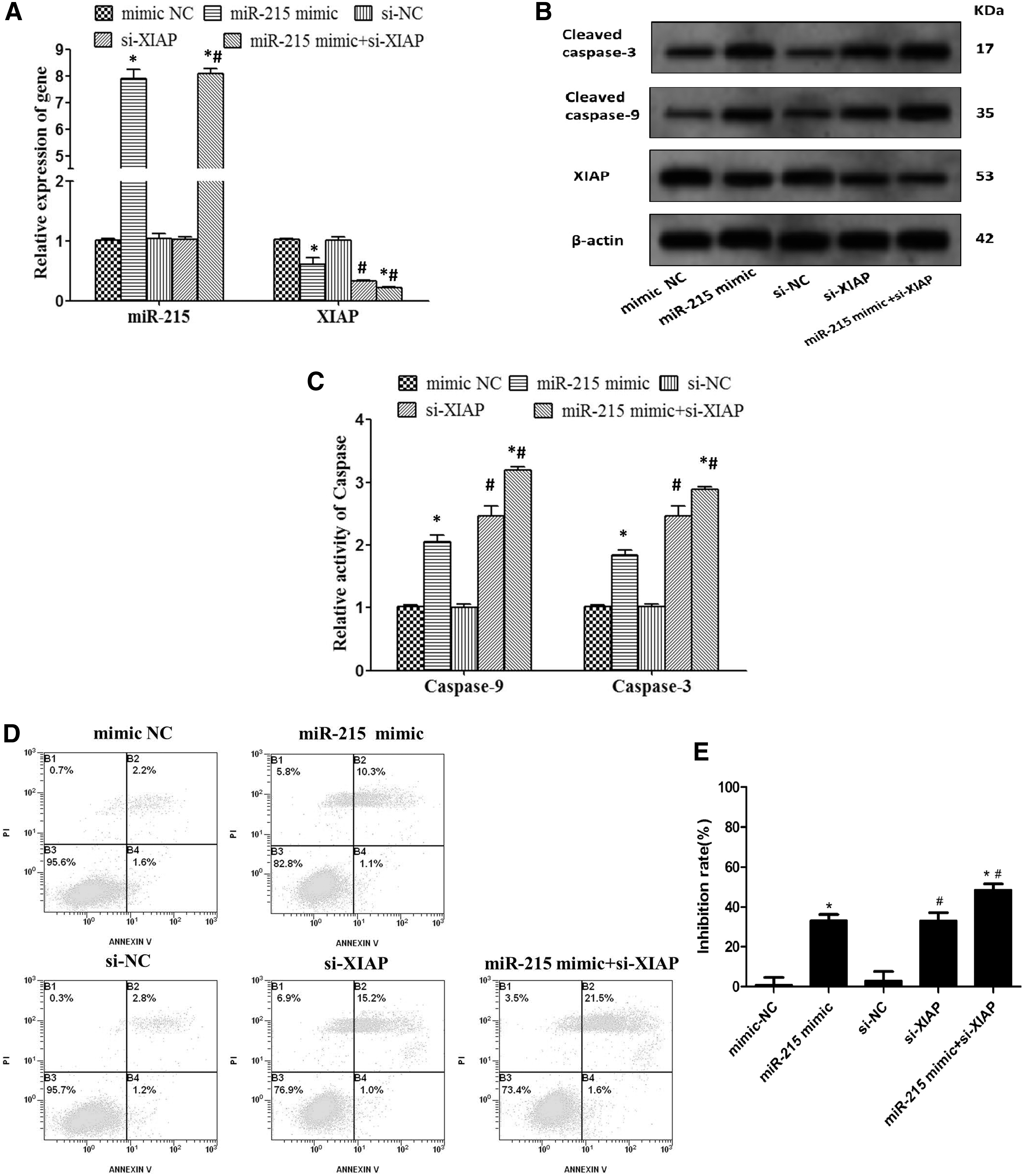
HCT116 cells were transfected with mimic NC, miR-215 mimic, si-NC, si-XIAP, miR-215 mimic + si-XIAP for 48 h.
Discussion
CRC is the third most common malignancy in the world and its mortality rate ranks the fourth place among the common malignant tumors. It was reported that there were 136,830 new cases of CRC patients with the number of mortality about 50,310 cases, accounting for 10% of total number of mortality in the United States in 2014. 12 In recent years, with the improvement of living standards, changes in diets, aging of populations, advancement of diagnosis technologies, and the census of CRC, the CRC incidence is in a rising trend in China, which brings serious threats to human life and health. 13 According to statistical data, a rapid increase in CRC incidence, as high as 50/100,000, is now observed. In addition, CRC has a very high degree of malignancy, and distant metastasis approximately occurs in half of CRC patients at the time of diagnosis. 14 Despite great improvement in the early diagnosis and clinical treatment techniques, the overall effect of CRC has not improved significantly, and its 3-year survival rate was 65.9%, 5-year survival rate was 42.3%, and the 5-year survival rate of patients with advanced CRC or distant metastasis was as low as 15%. 15 Unfortunately, the molecular mechanisms underlying the occurrence and progression of CRC remain generally unknown. Therefore, studies on these mechanisms make for its early diagnosis, good therapeutic effect, and prognosis.
The occurrence and formation of tumor is due to the activation of proto-oncogene and inactivation of antioncogene, and result in disorders of the normal cell proliferation, differentiation, and apoptosis process. 16 The dynamic balance between proliferation and apoptosis is closely associated with the occurrence and progression of the tumor. It will lead to uncontrolled growth, and evenly tumorigenesis by blocking apoptosis of cells with gene mutations. 17 IAPs is a kind of protein family that is structurally homologous and has the function of inhibiting apoptosis. XIAP, one of the IAPs family, is the sole apoptotic antagonizing factor that can inhibit the initiating stage and effective stage of apoptosis simultaneously. 18 XIAP has the characteristic structure of the IAPs family-BIR (baculovirus inhibitor of apoptosis repeat) domain, and with the three IAP repetitive sequences in the N-terminal and zinc finger domain in the C-terminal in BIR domain. 19 XIAP can directly bind and inhibit the apoptosis initiation molecule caspase-9 and apoptosis execution molecule caspase-3/-7, to hinder the conduction of downstream apoptotic signaling pathways and protect cells from apoptosis caused by various factors. 20 Overexpression of XIAP is closely associated with the occurrence, progression, and prognosis of a variety of tumors. Studies by Xiang et al. revealed that the expression of XIAP in the CRC tissues and tumor cell lines was significantly higher than that in the paracancerous tissues and normal colonic epithelial cells, and its expression was correlated with the survival and prognosis of patients. 4 Studies by Takeuchi et al. 5 showed that the expression of XIAP in the CRC tissues was closely associated with the progression and metastasis of the tumor, suggesting that the elevated expression of XIAP was possibly involved in the occurrence of the CRC. Consistent with the results of aforementioned research, this study showed that XIAP expression in CRC tissues was significantly higher compared with the paracancerous tissues and increased as the TNM staging went from I + II to III+IV. The background apoptotic rate of HCT116 cells was significantly lower, and the XIAP expression of HCT116 cells was significantly higher than that of CMEC, suggesting that overexpression of XIAP may play a role in the proliferation of CRC cells.
In studies about miR-215 and CRC, the expression levels of miR-215 in CRC tumor tissues was significantly lower than that in paracancerous tissues, 9,10 suggesting that the decrease of miR-215 may play a role in the occurrence of CRC. Researchers found that miR-215 was significantly reduced in CRC stem cells, and overexpression of miR-215 could inhibit clone formation and malignant growth ability of CRC stem cells, whereas decreasing miR-215 showed the opposite effect, indicating that miR-215 played an anticancer effect in CRC. 21 Also evidenced by Slattery, the higher of the TNM staging of CRC, the lower the expression of miR-215, suggesting that the decreased expression of miR-215 played a role in promoting the occurrence of CRC. 22 Li et al. revealed that patients with decreased miR-215 expression had a higher likelihood of recurrence than those with high miR-215 expression, and the patients with high miR-215 were more sensitive to chemotherapy than those with low miR-215, 23 suggesting that miR-215 upregulation had the role of enhancing the chemosensitivity and inhibiting the recurrence of CRC. Interestingly, miR-215 could play a role of antitumor, such as regulating the expression of P53 gene, upregulating the CDKN1A/p21 gene expression and blocking the cell cycle progression. 24 In addition, miR-215 could upregulate the expression of P21 in a P53-dependent manner, and induce cell cycle arrest in G2 phase, thus inhibiting the proliferation of CRC cells. 25 This study also showed that the expression levels of miR-215 in CRC tissues and CRC cell line HCT116 were significantly lower than those in the paracancerous tissues and CMEC, respectively.
Association analyses of clinicopathological features with miR-215 and XIAP showed that they both were associated with degrees of differentiation, TNM staging and lymph node metastasis of CRC patients (p < 0.05; Table 1). There was a negative correlation between the expression levels of miR-215 and XIAP in CRC tissues (r = −0.723, p = 0.033). Furthermore, to explore the molecular mechanism of miR-215 and XIAP in CRC apoptosis, it could be forecasted by accessing to
In summary, the low expression of miR-215 and the high expression of XIAP in CRC tissues may be potential tumor markers as well as relevant therapeutic targets. Also, miR-215 promotes apoptosis of CRC cells by downregulating XIAP.
Footnotes
Author Contributions
The authors declare that all the listed authors have participated actively in the study and all meet the requirements of the authorship. C.L. and M.H. designed the study and wrote the protocol. B.C. performed research/study, K.L. contributed important reagents. Y.L. and X.Z. managed the literature searches and analyses. M.H. undertook the statistical analysis. C.L. and G.S. prepared the first draft of the article.
Disclosure Statement
The authors have read and understood Current Oncology's policy on disclosing conflicts of interest and declare that they have none.
Funding Information
This study has no fund support and is self-funded.