
Abstract
Background:
Nasopharyngeal carcinoma (NPC) is leading form of cancer occurring in a few well-defined regions, including southern China. NPC possess a unique and intricate etiology that remains to be elucidated. Herein, we determine expression patterns of CCNB2 and NKX3-1 and identify their roles in NPC.
Materials and Methods:
Gene-expression profiles of NPC in the Gene Expression Omnibus (GEO) were analyzed. Cell viability, invasion, apoptosis, cell cycle entry and mitochondrial membrane potential (MMP) were evaluated in the presence of NKX3-1 or in the absence of CCNB2.
Results:
In all, 187 upregulated genes and 683 downregulated genes were obtained by analyzing GSE13597. NKX3-1, the downregulated gene, and CCNB2, the upregulated one, were further confirmed by in vitro studies. Overexpression of NKX3-1 was shown to inhibit NPC cell viability and invasion. Knockdown of CCNB2 was demonstrated to reduce cell cycle entry and MMP but induce apoptosis in NPC cells.
Conclusions:
Taken together, the key finding obtained from the study supports CCNB2 and NKX3-1 as two promising therapeutic candidates for NPC. Molecular mechanisms that control CCNB2 or NKX3-1 disturbance require further investigation and clarification.
Introduction
Nasopharyngeal carcinoma (NPC) is a disease which accounts for 1.14% of total cancer cases and 0.96% of total cancer-related mortality in 2013 in China, with 42,100 new morbidities and 21,320 mortalities that year. 1 It has been documented that epithelial–mesenchymal transition is involved in metastasis of malignancies. 2 Despite great improvement of decentralized control of NPC by intensity-modulated radiation therapy and its combination with chemotherapy, the clinical therapy effect is not satisfied enough. 3 Unfortunately, the majority of NPC patients can be cured if early detection and treatment are performed, whereas distant metastasis is still the most menace and challenge to heal NPC patients at the advanced stages. 4 Therefore, it is in urgency to discover novel biomarkers of NPC to improve the diagnosis and treatment of NPC.
Belonging to the cyclin protein family, cyclin B2 (CCNB2) has been documented to be overexpressed in human cancers, and measurement of serum circulating CCNB2 mRNA can be used to observe and investigate metastasis and therapeutic treatments. 5 A prior study showed overexpression of CCNB2 in nonsmall cell lung cancer (NSCLC) tissues relative to corresponding adjacent normal lung tissues. 6 Meanwhile, another study also revealed that CCNB2 was upregulated in different types of human pituitary adenomas. 7 NKX3-1 gene is regarded as a prostate specific tumor suppressor. It can regulate multiple signaling pathways related to cell proliferation, cell cycle, apoptosis, and invasion. 8 In addition, despite that NKX3-1 can bind to some ER cis-regulatory elements in vivo, it plays a critical role in transcriptional repression of estrogen signaling. 9 Furthermore, a study conducted by Tan et al. provided evidence that there were promoting effects of transcriptional regulatory network among NKX3-1, androgen receptor, and the RAB GTPase signaling pathway on prostate cancer survival. 10 However, there are limited researches about the effect of CCNB2 and NKX3-1 on NPC. Therefore, the current study was performed to evaluate the specific mechanism of both CCNB2 and NKX3-1 in NPC.
Materials and Methods
Gene-expression profiles of NPC in the Gene Expression Omnibus
Microarray data deposited with Gene Expression Omnibus under accession number GSE13597 between normal nasopharyngeal tissues (n = 3) and NPC tissues (n = 25) were analyzed. Genes with a false discovery rate (FDR) p < 0.05 and an absolute |log2-fold change (FC)| > 1 were screened as differentially expressed genes (DEGs) between normal nasopharyngeal tissues and NPC tissues. The screened DEGs were then subject to pathway enrichment analysis using gene ontology (GO) using the Database for Annotation, Visualization and Integrated Discovery (DAVID, version 6.7,
CCNB2 knockdown or NKX3-1 overexpression
NPC cell lines CNE-2 and immortalized normal nasopharyngeal cell lines NP69, both purchased from Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, were maintained in RPMI-1640 supplemented with 10% fetal bovine serum (FBS; Gibco, Carlsbad, CA) and 1% penicillin–streptomycin in a humidified atmosphere of 5% CO2 at 37°C. Medium was renewed every 24–48 h. When cell confluence reached 80%–90%, cells were passaged. A siRNA targeting CCNB2 (5′-GGAGUUAAUUCUAAAGUUAAG-3′) was designed and delivered into CNE-2 cells to specifically deplete CCNB2 using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) as per the manufacturer's protocol. The pcDNA vector containing full-length wild-type NKX3-1 was transformed into the CNE-2 cells to express NKX3-1 also using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instruction. CNE-2 cells treated with phosphate-buffered saline (PBS) were served as negative controls.
Cell Counting Kit-8 assay
Forty-eight hours after cell transfection, cells were seeded into 96-well plates and then incubated with 10 μL Cell Counting Kit-8 (CCK-8) solution (5 mg/mL; Sigma-Aldrich, St Louis, MO) every other day at 37°C. Two hours later, the media was renewed using 150 μL dimethyl sulfoxide (Sigma-Aldrich) in each well to dissolve the formazan crystals. Absorbance was read at 450 nm using a Microplate reader (Bio-Rad, Hercules, CA).
Transwell invasion assays
Cell invasion assays were manipulated using transwell chamber assays (BD Biosciences, San Jose, CA) as per the manufacturer's protocol. In brief, 48 h after cell transfection, cells were added to the top chambers at 1 × 105 cells per 100 μL. Dulbecco's modified Eagle's medium (500 μL) with 10% FBS was used as a chemoattractant in the bottom chambers, which were coated with 40 μL Matrigel (BD Biosciences). Forty-eight hours after cell harvest at 37°C with 5% CO2, the cells that transferred to the bottom chamber were stained with 0.1% crystal violet at room temperature for 10 min and then counted in six fields per well using the Olympus inverted microscope (Olympus, Tokyo, Japan).
Flow cytometry analysis
An apoptosis assay was performed using the FITC Annexin V Apoptosis Detection Kit (KeyGen, Nanjing, Jiangsu, China) following the protocol. In brief, following 48 h cell transfection, a suspension (100 μL) of 4 × 105 CNE-2 cells was stained with 10 μL of Annexin V and 5 μL of Propidium Iodide (PI) at room temperature for 15 min and analyzed by flow cytometry (FACScan®; BD Biosciences) equipped with CellQuest software (BD Biosciences). Cells for cell cycle analysis were stained with PI alone using the CycleTEST™ Plus DNA Reagent Kit (BD Biosciences) as per the manufacturer's recommendations and also analyzed by FACScan.
Measurements of mitochondrial membrane potential
Forty-eight hours after cell transfection, cells were seeded into six-well plates at 1 × 105 cells/mL and resuspended in 500 μL PBS. Living cells were stained by Rhodamine123 (10 mg/L) and analyzed by FACScan.
RNA isolation and quantitation
Total RNA was extracted from cultured cells using TRIzol reagent (Invitrogen) and was reverse-transcribed to complementary DNA (cDNA) using First-Strand cDNA Synthesis Kit (Tiangen Biotech, Beijing, China) according to the protocol. Real-time polymerase chain reaction (PCR) was determined using SYBR®Premix Ex TaqTM II (RR820A; Takara Bio, Inc., Dalian, China) on the Applied Biosystems 7500 Real-Time PCR System with each reaction run in triplicate (Applied Biosystems, Carlsbad, CA). For all samples, the expression levels of CCNB2 and NKX3-1, relative to housekeeping gene GAPDH, were determined using the comparative threshold cycle (Ct) method with the formula 2−ΔΔCt.
Western blot analysis
Proteins were extracted from cultured cells using lysis buffer (BIOSS, Beijing, China). Cell lysate proteins (30 μg for each well) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Beyotime Institute of Biotechnology). The membranes were incubated with anti-CCNB2 antibody (1: 1000, Cat No. ab185622; Abcam, Cambridge, MA), anti-NKX3-1 antibody (1: 1000, Cat No. ab196020; Abcam), and anti-β-actin antibody (1:5000, Cat No. ab8227; Abcam) overnight at 4°C. Next, the membranes were incubated with horseradish peroxidase-conjugated goat antirabbit IgG antibody (1:3000, Cat No. ab205718; Abcam) at room temperature for 1 h, and the bands were visualized and analyzed by the ImageJ software available from the NIH Web site (Bethesda, MD). β-actin was used for normalization.
Statistical analysis
Representative experiments repeated at least thrice are expressed as mean ± standard deviation. All pairwise comparisons were performed using the least significant difference test. Differences between normally distributed values of multiple experimental groups were evaluated using one-way analysis of variance. A two-tailed probability value less than 0.05 was set as the level of significance. All statistics were performed using IBM SPSS Statistics 21.0 (Chicago, IL).
Results
DEGs between normal nasopharyngeal tissues and NPC tissues from GSE13597
First, the authors analyzed GSE13597 microarray and obtained 11,139 upregulated genes and 11,076 downregulated genes between normal nasopharyngeal tissues and NPC tissues. According to a FDR p < 0.05 and an absolute |log2 FC| > 1, a common set of 870 DEGs (Fig. 1) was yielded, including 187 genes with increased abundance (>1-log2FC) and 683 genes with decreased abundance (<-1-log2FC) in NPC tissues relative to normal nasopharyngeal tissues.
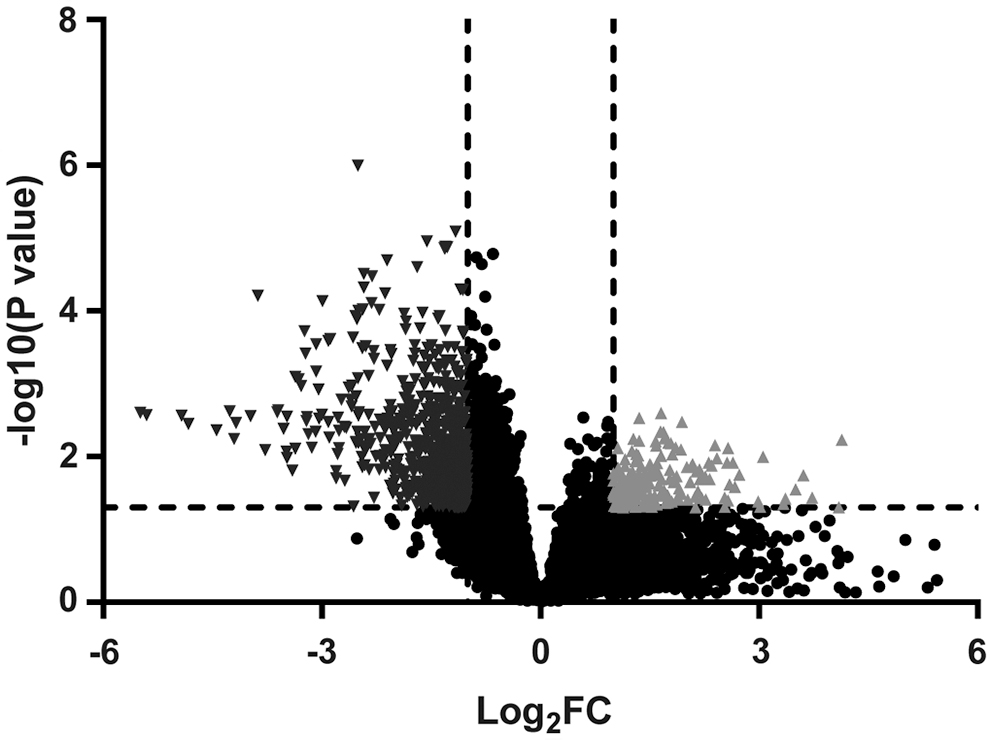
A volcano plot that depicts 187 upregulated genes and 683 downregulated genes between normal nasopharyngeal tissues (n = 3) and NPC tissues (n = 25) according to a FDR p < 0.05 and an absolute |log2 FC| > 1. Red points represent upregulated genes. Blue points represent downregulated genes. Black points represent altered genes showing no significant difference between normal nasopharyngeal tissues and NPC tissues. FC, fold change; FDR, false discovery rate; NPC, nasopharyngeal carcinoma.
Enrichment analysis for 870 DEGs
Next, 870 DEGs were subject to pathway enrichment analysis using GO using the Database for DAVID and KEGG pathway. There were 24 GO terms (Table 1) in which at least 15 DEGs were enriched (p < 0.05), and there were 13 pathways (Table 2) in which at least 10 DEGs were enriched (p < 0.05). The authors selected CCNB2, the upregulated gene (FC: 2.39, p = 0.007), and NKX3-1, the downregulated one (FC: −3.20, p = 0.003), both of which were associated with cancer-related pathways and cell cycle control pathways.
Gene Ontology Analysis for 870 Differentially Expressed Genes
Kyoto Encyclopedia of Genes and Genomes Analysis for 870 Differentially Expressed Genes
AMPK, AMP-activated protein kinase; cAMP, cyclic adenosine 3′,5′-monophosphate; FoxO, forkhead box O.
Expression patterns of CCNB2 and NKX3-1 in NPC cells
To determine expression patterns of CCNB2 and NKX3-1 in NPC, the authors quantified the expressions of CCNB2 and NKX3-1 in NPC cell lines CNE-2 and immortalized normal nasopharyngeal cell lines NP69. The results showed that the NKX3-1 was downregulated and the CCNB2 was upregulated in CNE-2 cells in contrast to NP6 cells (Fig. 2A). Then, the authors manipulated the expressions of CCNB2 and NKX3-1 in CNE-2 cells to study their effects on NPC. The qRT-PCR and Western blot analyses showed that siRNA targeting CCNB2 reduced the mRNA expression and protein level of CCNB2 (p < 0.01), and the delivery of pcDNA vectors containing full-length wild-type NKX3-1 increased the mRNA expression and protein level of NKX3-1 in CNE-2 cells (p < 0.01, Fig. 2B, C).

Expression patterns of CCNB2 and NKX3-1 in NPC cells.
Overexpression of NKX3-1 inhibits NPC cell viability and invasion
Since NKX3-1 was enriched in cancer-related pathways by enrichment analysis, the authors performed CCK-8 and Transwell invasion assays to investigate the effect of NKX3-1 on CNE-2 cell viability and invasion. As depicted in Figure 3A and B, 48 h after delivery of pcDNA vectors containing NKX3-1 into CNE-2 cells, cell viability and invasion were reduced, relative to untreated CNE-2 cells and PBS-treated CNE-2 cells (p < 0.05). These results uncovered that overexpression of NKX3-1 was shown to inhibit NPC cell viability and invasion, suggesting that NKX3-1 might participate in NPC development.
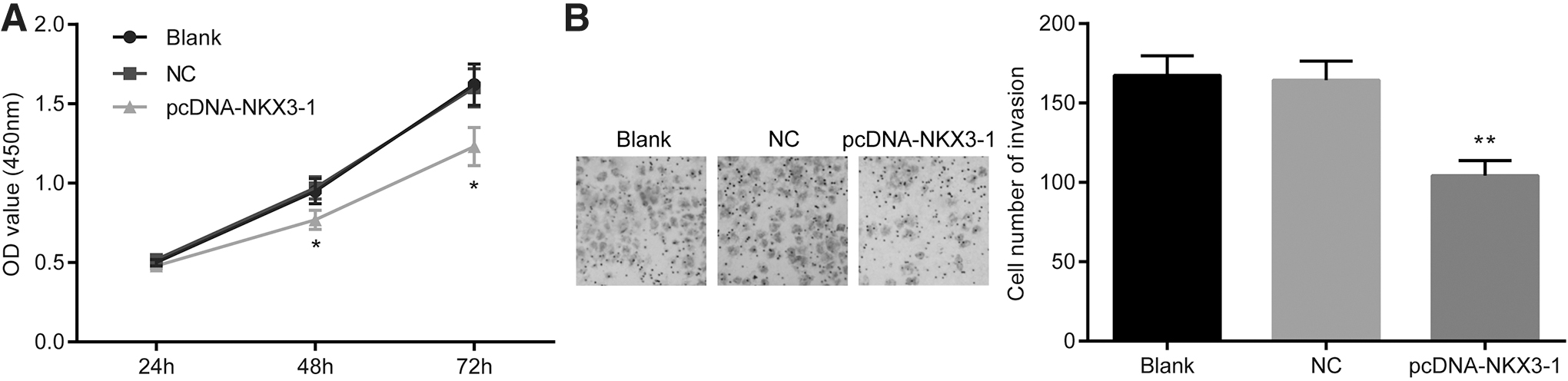
Overexpression of NKX3-1 inhibits CNE-2 cell viability and invasion. The pcDNA vector containing full-length wild-type NKX3-1 was transformed into the CNE-2 cells to express NKX3-1 (pcDNA-NKX3-1 group). PBS-treated CNE-2 cells were served as negative controls (NC group).
Knockdown of CCNB2 reduces cell cycle entry and mitochondrial membrane potential but induces apoptosis in NPC cells
As mentioned above, CCNB2 was enriched in cell cycle control pathways. Cell arrest in G2/M phase is indicative of cell apoptosis, and thus, the authors examined cell cycle entry in the absence of CCNB2. The results showed that more CNE-2 cells were arrested in G2/M phase after CCNB2 depletion (p < 0.05, Fig. 4A). The authors also found that CCNB2 depletion increased CNE-2 cell apoptosis (p < 0.05, Fig. 4B). Since changes of mitochondrial membrane potential (MMP) also reflected cell apoptosis, the authors measured MMP in CCNB2-depleted CNE-2 cells and found that CCNB2 knockdown reduced MMP (p < 0.05, Fig. 4C). The data suggested that knockdown of CCNB2 inhibits NPC cell apoptosis.

Knockdown of CCNB2 reduces cell cycle entry and MMP but induces apoptosis in CNE-2 cells. A siRNA targeting was designed and delivered into CNE-2 cells to specifically deplete CCNB2 (si-CCNB2 group). PBS-treated CNE-2 cells were served as negative controls (NC group).
Discussion
As a rare malignancy, NPC is characterized by specific geographic spread and mostly occurs in southern China. 11 Moreover, loco-regionally advanced disease occurs in 75%–90% of newly diagnosed NPC patients, which is generally accompanied by cervical nodal metastases. 12 Despite great advances of prognosis and treatment for NPC, distant failure remains a key challenge. 13 Therefore, it is necessary to find novel biomarkers for treatment of NPC. The present study aimed to explore the effect of CCNB2 and NKX3-1 on NPC. The major finding of this work showed that NKX3-1 could inhibit the proliferation and apoptosis of NPC cells, and CCNB2 could regulate the G2/M phase of cell cycle and inhibit the apoptosis of NPC cells.
The first interesting finding was that CCNB2 expression was increased and NKX3-1 expression was decreased in NPC cells. As demonstrated in a prior study, similar results were obtained that CCNB2 expression was higher in NSCLC tissues than in adjacent normal lung tissues. 6 Moreover, there was strong evidence that CCNB2 was highly expressed in different types of human pituitary adenomas, which were consistent with their identification. 7 This finding is consistent with that of Bowen et al. who observed loss of NKX3-1 in human prostate cancers. 14 Observations of a recent study displayed that NKX3-1 is poorly expressed in androgen-independent PC cells. 15 Importantly, gene expression profiles of NPC in their study screen the upregulated CCNB2 and downregulated NKX3-1. Therefore, overexpression of CCNB2 and inhibitory expression of NKX3-1 might be associated with the development of NPC.
The obvious finding to emerge from this analysis was that cell proliferation was decreased and cell apoptosis was increased following NKX3-1 upregulation. It was observed by Ren et al. that when NKX3-1 expression was increased in different prostate cancer cell lines, suppression of cell proliferation and cell survival was observed, as well as increased apoptotic cell. 8 Besides, a prior study revealed that cell proliferation was inhibited, cell death was promoted in PC3 cells by overexpression of either NKX3-1 or p27(KIP1), and the coexpression of Nkx3.1 and p27(KIP1) resulted in increase of cell apoptosis by downregulating Bcl-2 oncogene and upregulating Bax expression. 15 In addition, the results obtained from their study highlighted that CCNB2 silencing suppressed cell cycle arrest and MMP and enhanced the apoptosis of NPC cells. It is common knowledge that mitochondrial dysfunction is implicated in cell apoptosis and is central to the apoptotic pathway. 16 Moreover, a recent study provided evidence that restoration of MMP was involved in the inhibition of apoptosis induced by Gelsolin. 17 It was documented that downregulation of CCNB2 induced by KPNA2 knockdown contributed to repression of cell proliferation and promotion of cell cycle arrest in the G2/M phase. 18 Another study presented that apoptosis rate and cell cycle arrest were enhanced by metformin, which was associated with downregulation of CCNB2. 19 Based on the above, CCNB2 downregulation or NKX3-1 overexpression represses NPC cell proliferation and induces cell apoptosis.
Conclusion
Taken together, NKX3-1 could inhibit the proliferation and apoptosis of NPC cells, and CCNB2 could regulate the G2/M phase of cell cycle and inhibit the apoptosis of NPC cells. This finding suggested that NKX3-1 and CCNB2 are promising therapeutic modules for patients who suffered from NPC. Nevertheless, because of the case that the authors researched was small and they only use one cell line to verify the hypothesis, the results may have some limitations. The authors will avoid this problem in the follow-up study and analyze more data to make the results more convincing. Since the in-depth investigations on the function of NKX3-1 and CCNB2 on NPC are still under development, further explorations of the underlying mechanisms of NKX3-1 and CCNB2 are still necessary. Besides, despite interesting promises for the prognostic and therapeutic approaches in NPC, clinical experiments of fully developed NKX3-1 and CCNB2-based therapeutic treatment should be also enhanced in the future.
Footnotes
Acknowledgment
The authors give their sincere gratitude to the reviewers for their comments and the support by their institution.
Authorship Confirmation Statement
Substantial contributions to the conception or design of the work: D.Q.; Acquisition, analysis, or interpretation of data for the work: C.C., G.J., and J.H.; Drafting the work and revising it critically for important intellectual content: W.Z.; and Final approval of the version to be published: D.Q. Authors listed have approved the article that is enclosed.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.